

07.Молекулярная генетика 2012.ppt
- Количество слайдов: 121
 Эволюция понятия – «ген» n Ген – единица наследственности, определяющая развитие какоголибо признака организма (1909) n Ген – неделимая единица мутации, рекомбинации и функции (середина 20 -х годов ХХ века) n 50 -е годы ХХ века – сформировано важнейшее положение биохимической генетики «Один ген – один фермент» n 1957 год – гипотеза «Один ген – один фермент» уточнена – «Один ген одна полипептидная цепь» 1
Эволюция понятия – «ген» n Ген – единица наследственности, определяющая развитие какоголибо признака организма (1909) n Ген – неделимая единица мутации, рекомбинации и функции (середина 20 -х годов ХХ века) n 50 -е годы ХХ века – сформировано важнейшее положение биохимической генетики «Один ген – один фермент» n 1957 год – гипотеза «Один ген – один фермент» уточнена – «Один ген одна полипептидная цепь» 1
 Основы молекулярной генетики 2
Основы молекулярной генетики 2
 Эволюция понятия – «ген» Современное представление о структуре гена, его функционировании, регуляции его активности складывалось во 2 -й половине XX столетия: n Открыта двухспиральная структура ДНК n Выделена РНК и выяснена ее роль в передаче наследственной Информации от ДНК к белку n Расшифрован генетический код n Показана дробимость гена при рекомбинации и мутации 3
Эволюция понятия – «ген» Современное представление о структуре гена, его функционировании, регуляции его активности складывалось во 2 -й половине XX столетия: n Открыта двухспиральная структура ДНК n Выделена РНК и выяснена ее роль в передаче наследственной Информации от ДНК к белку n Расшифрован генетический код n Показана дробимость гена при рекомбинации и мутации 3
 Эволюция понятия – «ген» n n n Единицу рекомбинации назвали реконом, а единицу мутации – мутоном, однако в дальнейшем было показано, что и мутон и рекон соответствует одной паре нуклеотидов Единицу генетической функции назвали цистроном, что совпадает с первоначальным понятием «ген» В настоящее время термин «цистрон» употребляется достаточно редко как синоним понятия гена в тех случаях, когда хотят подчеркнуть его функциональное значение 4
Эволюция понятия – «ген» n n n Единицу рекомбинации назвали реконом, а единицу мутации – мутоном, однако в дальнейшем было показано, что и мутон и рекон соответствует одной паре нуклеотидов Единицу генетической функции назвали цистроном, что совпадает с первоначальным понятием «ген» В настоящее время термин «цистрон» употребляется достаточно редко как синоним понятия гена в тех случаях, когда хотят подчеркнуть его функциональное значение 4
 Эволюция понятия – «ген» n В современной генетической литературе нет единого общепринятого определения «ген» n В основе одних определений положена структурная организация гена, других – функция в организме, в третьих – ген рассматривается как единица в процессе транскрипции n Структурный ген – это участок ДНК (или РНК у некоторых вирусов), определяющий линейную последовательность полипептидной цепи или одной молекулы т. РНК или р. РНК 5
Эволюция понятия – «ген» n В современной генетической литературе нет единого общепринятого определения «ген» n В основе одних определений положена структурная организация гена, других – функция в организме, в третьих – ген рассматривается как единица в процессе транскрипции n Структурный ген – это участок ДНК (или РНК у некоторых вирусов), определяющий линейную последовательность полипептидной цепи или одной молекулы т. РНК или р. РНК 5
 ¡ История гена с 1860 года до программы ENCODE
¡ История гена с 1860 года до программы ENCODE
 Определение 1860 – 1900 х: «Ген – это дискретная единица наследственности» ¡ Первоначальные понятия гена основывались на выделении отличительного признака организма и его наследуемости. Слово ген впервые использовал Вильгельм Иоганнсен (Wilhelm Johannsen) в 1909 году, основываясь на концепции Грегора Менделя (Mendel 1866).
Определение 1860 – 1900 х: «Ген – это дискретная единица наследственности» ¡ Первоначальные понятия гена основывались на выделении отличительного признака организма и его наследуемости. Слово ген впервые использовал Вильгельм Иоганнсен (Wilhelm Johannsen) в 1909 году, основываясь на концепции Грегора Менделя (Mendel 1866).
![¡ Иоганнсен назвал геном «определенные условия, основы и детерминанты, присутствующие [в гаметах], являющиеся уникальными](https://present5.com/presentation/325077925_438361713/image-8.jpg "¡ Иоганнсен назвал геном «определенные условия, основы и детерминанты, присутствующие [в гаметах], являющиеся уникальными") ¡ Иоганнсен назвал геном «определенные условия, основы и детерминанты, присутствующие [в гаметах], являющиеся уникальными и самостоятельными, и поэтому независимыми путями определяющие многие характеристики организма»
¡ Иоганнсен назвал геном «определенные условия, основы и детерминанты, присутствующие [в гаметах], являющиеся уникальными и самостоятельными, и поэтому независимыми путями определяющие многие характеристики организма»
 Определение 1910 х: «Ген – это отдельный локус» ¡ Следующее важное открытие сделал американский генетик Томас Хант Морган (Thomas Hunt Morgan) со своими студентами, изучая расщепление мутаций у плодовых мушек Drosophila melanogaster. Они предложили модель, в которой гены расположены линейно и их способность к обмену (кроссоверу) пропорциональна расстоянию между ними.
Определение 1910 х: «Ген – это отдельный локус» ¡ Следующее важное открытие сделал американский генетик Томас Хант Морган (Thomas Hunt Morgan) со своими студентами, изучая расщепление мутаций у плодовых мушек Drosophila melanogaster. Они предложили модель, в которой гены расположены линейно и их способность к обмену (кроссоверу) пропорциональна расстоянию между ними.
 Определение 1940 х: «Ген – это чертеж белка» ¡ Изучая метаболизм у гриба Neurospora, Бидл и Тейтем (Beadle and Tatum 1941) обнаружили, что мутации в генах могли вызывать дефекты в звеньях метаболических путей. Это привело к определению «один ген – один фермент» , которое впоследствии преобразовалось в «один ген – один полипептид» . С этой точки зрения, под «геном» неявно подразумевалась информация, стоящая за отдельными молекулами в биохимическом пути. Этот подход стал более явным и механистическим в последующие десятилетия.
Определение 1940 х: «Ген – это чертеж белка» ¡ Изучая метаболизм у гриба Neurospora, Бидл и Тейтем (Beadle and Tatum 1941) обнаружили, что мутации в генах могли вызывать дефекты в звеньях метаболических путей. Это привело к определению «один ген – один фермент» , которое впоследствии преобразовалось в «один ген – один полипептид» . С этой точки зрения, под «геном» неявно подразумевалась информация, стоящая за отдельными молекулами в биохимическом пути. Этот подход стал более явным и механистическим в последующие десятилетия.
 Определение 1950 х: «Ген – это физическая молекула» ¡ Тот факт, что рентгеновские лучи могут вызывать мутации, показал, что наследственность имеет физическую молекулярную основу (Muller 1927).
Определение 1950 х: «Ген – это физическая молекула» ¡ Тот факт, что рентгеновские лучи могут вызывать мутации, показал, что наследственность имеет физическую молекулярную основу (Muller 1927).
 того, что живые невирулентные штаммы бактерии") ¡ Следующим доказательством была демонстрация Гриффитом (Griffith 1928) того, что живые невирулентные штаммы бактерии Pneumococcus могут извлекать нечто из вирулентных, но мертвых штаммов и превращаться в вирулентные
¡ Следующим доказательством была демонстрация Гриффитом (Griffith 1928) того, что живые невирулентные штаммы бактерии Pneumococcus могут извлекать нечто из вирулентных, но мертвых штаммов и превращаться в вирулентные
 ¡ Позднее выяснилось, что это вещество может разрушаться под действием фермента ДНКазы (Avery et al. 1944). В 1955 году Херши и Чейз установили, что веществом, которое передается бактериофагом своему потомку, является ДНК, а не белок
¡ Позднее выяснилось, что это вещество может разрушаться под действием фермента ДНКазы (Avery et al. 1944). В 1955 году Херши и Чейз установили, что веществом, которое передается бактериофагом своему потомку, является ДНК, а не белок
 ¡ Более того, идея о том, что продуктом гена является вещество, способное к диффузии, легла в основу цис-транс-теста (cis-trans-test), использовавшегося в то время в бактериологии для определения принадлежности двух рецессивных аллелей (мутаций), сходных в фенотипическом выражении, к одному и тому же или разным генам, по способности соответствующих участков ДНК комплементировать друг с другом. С этой точки зрения С. Бензер предложил для определения понятия гена почти не употребляемый в настоящее время термин «цистрон» – участок ДНК, отвечающий за единичную функцию (Benzer 1955).
¡ Более того, идея о том, что продуктом гена является вещество, способное к диффузии, легла в основу цис-транс-теста (cis-trans-test), использовавшегося в то время в бактериологии для определения принадлежности двух рецессивных аллелей (мутаций), сходных в фенотипическом выражении, к одному и тому же или разным генам, по способности соответствующих участков ДНК комплементировать друг с другом. С этой точки зрения С. Бензер предложил для определения понятия гена почти не употребляемый в настоящее время термин «цистрон» – участок ДНК, отвечающий за единичную функцию (Benzer 1955).
 Определение 1960 х: «Ген – это транскрибируемый код» ¡ Трехмерная структура ДНК, открытая Уотсоном и Криком (Watson and Crick 1953), позволила объяснить механизм, обеспечивающий ее функции в качестве молекулы наследственности. Спаривание оснований показало, как генетическая информация может копироваться, а существование двух нитей ДНК – как случайные ошибки репликации могут приводить к мутациям в одной из дочерних копий ДНК.
Определение 1960 х: «Ген – это транскрибируемый код» ¡ Трехмерная структура ДНК, открытая Уотсоном и Криком (Watson and Crick 1953), позволила объяснить механизм, обеспечивающий ее функции в качестве молекулы наследственности. Спаривание оснований показало, как генетическая информация может копироваться, а существование двух нитей ДНК – как случайные ошибки репликации могут приводить к мутациям в одной из дочерних копий ДНК.
 последовательностей ДНК, кодирующих") ¡ В 1965 году обнаружили, что молекулы РНК – копии (транскрипты) последовательностей ДНК, кодирующих белки – преобразуются (транслируются) в аминокислотные последовательности (Nirenberg et al. 1965, Söll et al. 1965). Фрэнсис Крик (Francis Crick 1958) обобщил понятие «экспрессия гена» как поток информации от нуклеиновой кислоты к белку (а не обратно) и сформулировал «центральную догму» молекулярной генетики: «Перенос генетической информации идет в направлении ДНК – РНК – белок» .
¡ В 1965 году обнаружили, что молекулы РНК – копии (транскрипты) последовательностей ДНК, кодирующих белки – преобразуются (транслируются) в аминокислотные последовательности (Nirenberg et al. 1965, Söll et al. 1965). Фрэнсис Крик (Francis Crick 1958) обобщил понятие «экспрессия гена» как поток информации от нуклеиновой кислоты к белку (а не обратно) и сформулировал «центральную догму» молекулярной генетики: «Перенос генетической информации идет в направлении ДНК – РНК – белок» .
 ¡ ¡ Однако уже тогда существовали исключения из этого определения: было известно, что некоторые гены кодируют не белки, а функциональные молекулы РНК, такие как рибосомальная РНК (р. РНК). Кроме того, в РНК-содержащих вирусах ген имеет рибонуклеиновую природу. Молекулярный взгляд на ген 1960 -х годов можно сформулировать так: ген представляет собой заключенный в нуклеиновой кислоте код, по которому образуется функциональный продукт.
¡ ¡ Однако уже тогда существовали исключения из этого определения: было известно, что некоторые гены кодируют не белки, а функциональные молекулы РНК, такие как рибосомальная РНК (р. РНК). Кроме того, в РНК-содержащих вирусах ген имеет рибонуклеиновую природу. Молекулярный взгляд на ген 1960 -х годов можно сформулировать так: ген представляет собой заключенный в нуклеиновой кислоте код, по которому образуется функциональный продукт.
 Определение 1970 х-1980 х: «Ген – это открытая рамка считывания (open reading frame – ORF) ¡ Развитие в 1970 х годах методик клонирования генов (получения множества их копий) и секвенирования вкупе со знаниями о генетическом коде привело к прорыву в области молекулярной биологии. Мы получили подробную информацию об организации экспрессии генов – того, как из генов получаются белки.
Определение 1970 х-1980 х: «Ген – это открытая рамка считывания (open reading frame – ORF) ¡ Развитие в 1970 х годах методик клонирования генов (получения множества их копий) и секвенирования вкупе со знаниями о генетическом коде привело к прорыву в области молекулярной биологии. Мы получили подробную информацию об организации экспрессии генов – того, как из генов получаются белки.
 ¡ Во многих случаях по последовательности нуклеотидов можно предположить структуру и функцию гена и его продукта. Эта ситуация породила новую концепцию «номинального гена» , который определяется предсказанной последовательностью, а не генетическим локусом, ответственным за фенотип (Griffiths and Stotz 2006)
¡ Во многих случаях по последовательности нуклеотидов можно предположить структуру и функцию гена и его продукта. Эта ситуация породила новую концепцию «номинального гена» , который определяется предсказанной последовательностью, а не генетическим локусом, ответственным за фенотип (Griffiths and Stotz 2006)
 ¡ Во многих случаях гены эффективно определяли как прокомментированные открытые рамки считывания – нуклеотидные последовательности ДНК (реже – РНК), потенциально способные кодировать белок и расположенные между первым, инициирующим кодоном и последним, стоп-кодоном (Doolittle 1986).
¡ Во многих случаях гены эффективно определяли как прокомментированные открытые рамки считывания – нуклеотидные последовательности ДНК (реже – РНК), потенциально способные кодировать белок и расположенные между первым, инициирующим кодоном и последним, стоп-кодоном (Doolittle 1986).
 Определение 1990 х-2000 х: «Аннотированная структурная единица генома ¡ По определению Human Genome Nomenclature Organization, ген – это «сегмент ДНК, влияющий на фенотип или функцию. В отсутствие проявленной функции ген может быть охарактеризован последовательностью, транскрипцией или гомологией» (Wain et al. 2002). А согласно определению Sequence Ontology Consortium, ген – это «локализуемый участок последовательности генома, соответствующий единице наследственности, которая ассоциирована с регуляторными, транскрипционными и другими функциональными участками последовательности» (Pearson 2006).
Определение 1990 х-2000 х: «Аннотированная структурная единица генома ¡ По определению Human Genome Nomenclature Organization, ген – это «сегмент ДНК, влияющий на фенотип или функцию. В отсутствие проявленной функции ген может быть охарактеризован последовательностью, транскрипцией или гомологией» (Wain et al. 2002). А согласно определению Sequence Ontology Consortium, ген – это «локализуемый участок последовательности генома, соответствующий единице наследственности, которая ассоциирована с регуляторными, транскрипционными и другими функциональными участками последовательности» (Pearson 2006).
 Современная компьютерная аналогия: «Ген – это подпрограмма в операционной системе генома» ¡ Наиболее популярная компьютерная метафора – это представление генов в качестве подпрограмм в огромных операционных системах (ОС). Как нуклеотиды генома складываются в код, который реализуется в процессах транскрипции и трансляции, так и геном можно рассматривать как ОС живого существа, а гены – как ее отдельные подпрограммы, которые неоднократно «запускаются» в процессе транскрипции.
Современная компьютерная аналогия: «Ген – это подпрограмма в операционной системе генома» ¡ Наиболее популярная компьютерная метафора – это представление генов в качестве подпрограмм в огромных операционных системах (ОС). Как нуклеотиды генома складываются в код, который реализуется в процессах транскрипции и трансляции, так и геном можно рассматривать как ОС живого существа, а гены – как ее отдельные подпрограммы, которые неоднократно «запускаются» в процессе транскрипции.
 ENCODE ¡ это сокращение от ENCyclopedia Of DNA Elements. Цель «Энциклопедии» – вывести науку на новый уровень понимания механизмов работы генома и его взаимосвязи с биохимическими процессами, идущими в организме.
ENCODE ¡ это сокращение от ENCyclopedia Of DNA Elements. Цель «Энциклопедии» – вывести науку на новый уровень понимания механизмов работы генома и его взаимосвязи с биохимическими процессами, идущими в организме.
 ¡ Новое определение гена формулируется так: «Ген – это совокупность геномных последовательностей, кодирующих сцепленный набор потенциально перекрывающихся функциональных продуктов»
¡ Новое определение гена формулируется так: «Ген – это совокупность геномных последовательностей, кодирующих сцепленный набор потенциально перекрывающихся функциональных продуктов»
 Особенности структуры вирусного генома ¡ Одно- и двух цепочечные ДНК, двух цепочечные РНК, одно - цепочечные РНК с «плюс-цепью» и с «минус-цепью» ¡ Наличие перекрывающихся генов (ген в гене) – имеются различные рамки считывания с одной и той же нуклеотидной последовательности 25
Особенности структуры вирусного генома ¡ Одно- и двух цепочечные ДНК, двух цепочечные РНК, одно - цепочечные РНК с «плюс-цепью» и с «минус-цепью» ¡ Наличие перекрывающихся генов (ген в гене) – имеются различные рамки считывания с одной и той же нуклеотидной последовательности 25
 Особенности структуры прокариотического генома n n n Содержит двухцепочечную ДНК, которая свободно «плавает» в цитоплазме Представлен двумя структурами n Хромосомами n Плазмидами Основное количество ДНК упаковано в 1 -2 хромосомы, которые могут быть линейными, но чаще - кольцевые Прокариотическая хромосома Плазмиды 26
Особенности структуры прокариотического генома n n n Содержит двухцепочечную ДНК, которая свободно «плавает» в цитоплазме Представлен двумя структурами n Хромосомами n Плазмидами Основное количество ДНК упаковано в 1 -2 хромосомы, которые могут быть линейными, но чаще - кольцевые Прокариотическая хромосома Плазмиды 26
 Особенности структуры генов прокариот n Гены объединены в группы или кластеры – все сцепленные гены кластера кодируют ферменты одного биосинтетического пути и транскрибируются на общую молекулу м. РНК – полицистронную м. РНК n Группа структурных генов прокариот, находящихся под контролем одного регуляторного участка, называют опероном n Между оперонами располагаются межгенные участки – спейсеры 27
Особенности структуры генов прокариот n Гены объединены в группы или кластеры – все сцепленные гены кластера кодируют ферменты одного биосинтетического пути и транскрибируются на общую молекулу м. РНК – полицистронную м. РНК n Группа структурных генов прокариот, находящихся под контролем одного регуляторного участка, называют опероном n Между оперонами располагаются межгенные участки – спейсеры 27
 Организация генетического материала у эукариот 28
Организация генетического материала у эукариот 28
 — один из двух типов нуклеиновых кислот, обеспечивающих хранение,") ¡ Дезоксирибонуклеи новая кислота (ДНК) — один из двух типов нуклеиновых кислот, обеспечивающих хранение, передачу из поколения в поколение и реализацию генетической программы ра звития и функционирования живых организмов. Основная роль ДНК в клетках — долговременное хранение информации о структуре РНК и белков.
¡ Дезоксирибонуклеи новая кислота (ДНК) — один из двух типов нуклеиновых кислот, обеспечивающих хранение, передачу из поколения в поколение и реализацию генетической программы ра звития и функционирования живых организмов. Основная роль ДНК в клетках — долговременное хранение информации о структуре РНК и белков.
 История изучения ¡ Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин» , поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальныеклетки, в которых нет ядра, тоже содержат нуклеиновые кислоты.
История изучения ¡ Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин» , поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальныеклетки, в которых нет ядра, тоже содержат нуклеиновые кислоты.
 ¡ ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
¡ ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
 ¡ Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т. е. представляет собой двойную спираль. Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0, 34 нм, на один оборот спирали приходится 10 пар нуклеотидов.
¡ Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т. е. представляет собой двойную спираль. Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0, 34 нм, на один оборот спирали приходится 10 пар нуклеотидов.
 — состоит из : 1) азотистого основания,") ¡ ¡ Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из : 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
¡ ¡ Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из : 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
 ¡ Гуанин Гуаниловый ( Г ) Тимин") Название нуклеотида ¡ Аденин Адениловый ( A) ¡ Гуанин Гуаниловый ( Г ) Тимин Тимидиловый ( Т) Цитозин Цитидиловый ( Ц) ¡
Название нуклеотида ¡ Аденин Адениловый ( A) ¡ Гуанин Гуаниловый ( Г ) Тимин Тимидиловый ( Т) Цитозин Цитидиловый ( Ц) ¡
 ¡ Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3'углеродом остатка дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого возникает фосфоэфирная связь (относится к категории прочных ковалентных связей). Один конец полинуклеотидной цепи заканчивается 5'-углеродом (его называют 5'-концом), другой — 3'-углеродом (3'-концом).
¡ Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3'углеродом остатка дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого возникает фосфоэфирная связь (относится к категории прочных ковалентных связей). Один конец полинуклеотидной цепи заканчивается 5'-углеродом (его называют 5'-концом), другой — 3'-углеродом (3'-концом).
 ¡ Закономерность, согласно которой нуклеотиды разных цепей ДНК строго упорядоченно располагаются (аденин — тимин, гуанин — цитозин) и избирательно соединяются друг с другом, называется принципом комплементарности.
¡ Закономерность, согласно которой нуклеотиды разных цепей ДНК строго упорядоченно располагаются (аденин — тимин, гуанин — цитозин) и избирательно соединяются друг с другом, называется принципом комплементарности.
 Репликация ДНК ¡ процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным.
Репликация ДНК ¡ процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным.
 ¡ С помощью геликаз в определенных участках ДНК расплетается, одноцепочечные участки ДНК связываются дестабилизирующими белками, образуется репликационная вилка.
¡ С помощью геликаз в определенных участках ДНК расплетается, одноцепочечные участки ДНК связываются дестабилизирующими белками, образуется репликационная вилка.
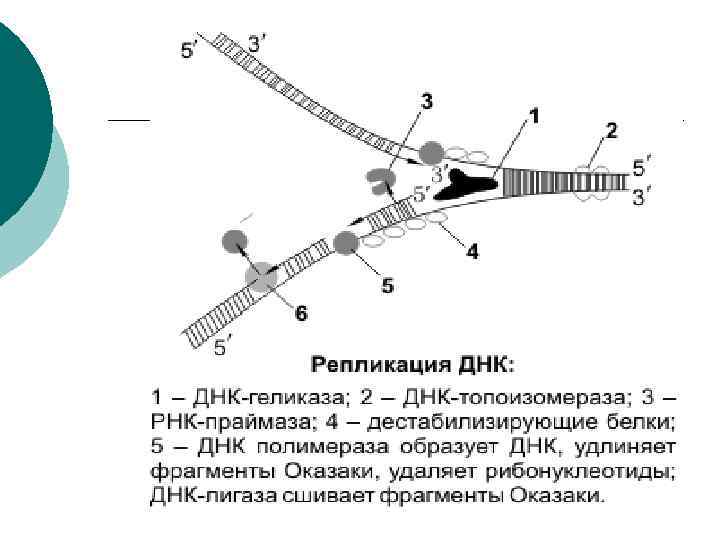
 ¡ ДНК-полимераза может присоединять нуклеотид только к 3'-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3'конца к 5'-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по -разному и в противоположных направлениях
¡ ДНК-полимераза может присоединять нуклеотид только к 3'-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3'конца к 5'-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по -разному и в противоположных направлениях
 ¡ На цепи 3'– 5' синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей. На цепи 5'– 3' — прерывисто, фрагментами (фрагменты Оказаки), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстаю щей).
¡ На цепи 3'– 5' синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей. На цепи 5'– 3' — прерывисто, фрагментами (фрагменты Оказаки), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстаю щей).
 ¡ Особенностью ДНК-полимеразы является то, что она может начинать свою работу только с «затравки» (праймера). Роль «затравок» выполняют короткие последовательности РНК, образуемые при участи фермента РНК-праймазы и спаренные с матричной ДНК. РНКзатравки после окончания сборки полинуклеотидных цепочек удаляются.
¡ Особенностью ДНК-полимеразы является то, что она может начинать свою работу только с «затравки» (праймера). Роль «затравок» выполняют короткие последовательности РНК, образуемые при участи фермента РНК-праймазы и спаренные с матричной ДНК. РНКзатравки после окончания сборки полинуклеотидных цепочек удаляются.
 ¡ Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
¡ Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
 Молекула ДНК может существовать в разных формах Молекула ДНК может в зависимости от условий существовать в разных формах. ¡ Наиболее распространой является В-форма. ¡ В этой форме находится основная часть ДНК в клетках. При такой организации плоскости азотистых оснований практически перпендикулярны оси двойной спирали, и каждая пара повёрнута относительно предыдущих на 36. На один виток спирали приходится примерно 10 нуклеотидных пар (9, 7 и 10, 6 в различных кристаллах), а длина составляет 3, 4 нм. ¡
Молекула ДНК может существовать в разных формах Молекула ДНК может в зависимости от условий существовать в разных формах. ¡ Наиболее распространой является В-форма. ¡ В этой форме находится основная часть ДНК в клетках. При такой организации плоскости азотистых оснований практически перпендикулярны оси двойной спирали, и каждая пара повёрнута относительно предыдущих на 36. На один виток спирали приходится примерно 10 нуклеотидных пар (9, 7 и 10, 6 в различных кристаллах), а длина составляет 3, 4 нм. ¡
 способствует переходу В") ¡ Установлено, что понижение относительной влажности при добавлениинеэлектролитов (напри мер, этанола) способствует переходу В формы в А-форму
¡ Установлено, что понижение относительной влажности при добавлениинеэлектролитов (напри мер, этанола) способствует переходу В формы в А-форму
 ¡ Существенным отличием А-формы от В-формы является то, что в Аформе пары оснований сдвинуты к периферии спирали почти на половину её радиуса, в результате чего пространство вдоль оси оказывается пустым.
¡ Существенным отличием А-формы от В-формы является то, что в Аформе пары оснований сдвинуты к периферии спирали почти на половину её радиуса, в результате чего пространство вдоль оси оказывается пустым.

 двуцепочечной ДНК. В отличие") ¡ Помимо В-формы и А-формы встречается также Zформа (нижнее изображение) двуцепочечной ДНК. В отличие от первых двух, она представляет собой левозакрученную спираль с длиной витка 4, 4 нм, на который приходится 12 нуклеотидных пар
¡ Помимо В-формы и А-формы встречается также Zформа (нижнее изображение) двуцепочечной ДНК. В отличие от первых двух, она представляет собой левозакрученную спираль с длиной витка 4, 4 нм, на который приходится 12 нуклеотидных пар

 Репарация ¡ ¡ ¡ Репарацией называется процесс устранения повреждений нуклеотидной последовательности ДНК. ферменты репарации В процессе восстановления структуры ДНК можно выделить следующие этапы: 1) ДНКрепарирующие нуклеазы распознают и удаляют поврежденный участок, в результате чего в цепи ДНК образуется брешь; 2) ДНКполимераза заполняет эту брешь, копируя информацию со второй цепи; 3) ДНК-лигаза «сшивает» нуклеотиды, завершая репарацию.
Репарация ¡ ¡ ¡ Репарацией называется процесс устранения повреждений нуклеотидной последовательности ДНК. ферменты репарации В процессе восстановления структуры ДНК можно выделить следующие этапы: 1) ДНКрепарирующие нуклеазы распознают и удаляют поврежденный участок, в результате чего в цепи ДНК образуется брешь; 2) ДНКполимераза заполняет эту брешь, копируя информацию со второй цепи; 3) ДНК-лигаза «сшивает» нуклеотиды, завершая репарацию.
 фоторепарация, 2) эксцизная, или дорепликативная, репарация,") Наиболее изучены три механизма репарации: ¡ ¡ 1) фоторепарация, 2) эксцизная, или дорепликативная, репарация, 3) пострепликативная репарация. Изменения структуры ДНК происходят в клетке постоянно под действием реакционноспособных метаболитов, ультрафиолетового излучения, тяжелых металлов и их солей и др. Поэтому дефекты систем репарации повышают скорость мутационных процессов, являются причиной наследственных заболеваний.
Наиболее изучены три механизма репарации: ¡ ¡ 1) фоторепарация, 2) эксцизная, или дорепликативная, репарация, 3) пострепликативная репарация. Изменения структуры ДНК происходят в клетке постоянно под действием реакционноспособных метаболитов, ультрафиолетового излучения, тяжелых металлов и их солей и др. Поэтому дефекты систем репарации повышают скорость мутационных процессов, являются причиной наследственных заболеваний.
 ¡ ДОКАЗАТЕЛЬСТВА РОЛИ ДНК КАК МАТЕРИАЛЬНОГО НОСИТЕЛЯ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ
¡ ДОКАЗАТЕЛЬСТВА РОЛИ ДНК КАК МАТЕРИАЛЬНОГО НОСИТЕЛЯ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ
 Опыты Ф. Гриффитса ¡ ¡ ¡ использовались два штамма пневмококков с противоположными признаками: с наличием и отсутствием капсул. Клетки капсульного штамма S были вирулентными, а бескапсульного - R - безвредными. Животные, зараженные вирулентным штаммом S, погибали. При введении бескапсульных бактерий (R) и клеток S-штамма, убитых нагреванием, мыши выживали. Казалось бы, полученные результаты были закономерны, а их причины - очевидны.
Опыты Ф. Гриффитса ¡ ¡ ¡ использовались два штамма пневмококков с противоположными признаками: с наличием и отсутствием капсул. Клетки капсульного штамма S были вирулентными, а бескапсульного - R - безвредными. Животные, зараженные вирулентным штаммом S, погибали. При введении бескапсульных бактерий (R) и клеток S-штамма, убитых нагреванием, мыши выживали. Казалось бы, полученные результаты были закономерны, а их причины - очевидны.
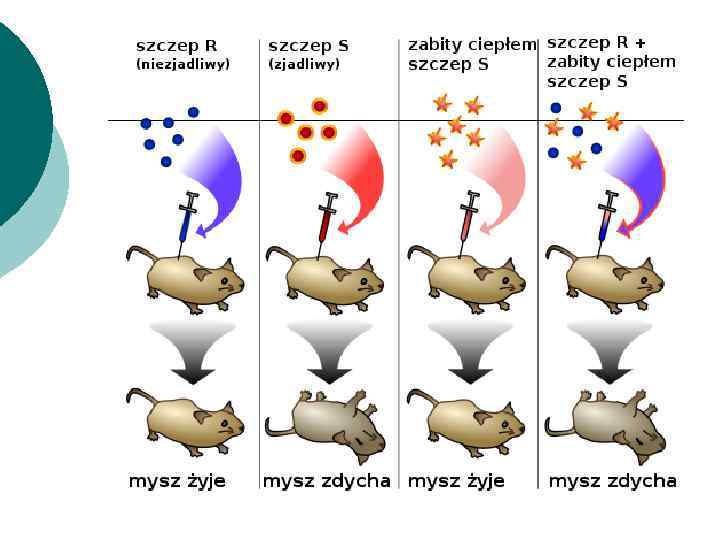
 ¡ Но совершенно обескураживающие результаты были получены у последней группы белых мышей. Этим животным вводили суспензию, содержащую живые клетки бескапсульного штамма и убитые вирулентные бактерии. Через некоторое время у мышей обнаруживались клинические признаки пневмококковой инфекции и животные погибали.
¡ Но совершенно обескураживающие результаты были получены у последней группы белых мышей. Этим животным вводили суспензию, содержащую живые клетки бескапсульного штамма и убитые вирулентные бактерии. Через некоторое время у мышей обнаруживались клинические признаки пневмококковой инфекции и животные погибали.
 ¡ Однако природу трансформирующего агента в то время установить не удалось. Было известно, что это вещество небелкового происхождения, т. к. все белки при нагревании подвергались денатурации.
¡ Однако природу трансформирующего агента в то время установить не удалось. Было известно, что это вещество небелкового происхождения, т. к. все белки при нагревании подвергались денатурации.
 ¡ ¡ ¡ Проведенный бактериологический анализ показал, что в тканях погибших мышей содержатся клетки пневмококка, окруженные капсулой. Следовательно, невирулентный бескапсульный штамм пневмококков под воздействием убитых бактерий S-штамма получал новый признак - капсулу - и приобретал вирулентные свойства. Такое явление Гриффитс назвал трансформацией.
¡ ¡ ¡ Проведенный бактериологический анализ показал, что в тканях погибших мышей содержатся клетки пневмококка, окруженные капсулой. Следовательно, невирулентный бескапсульный штамм пневмококков под воздействием убитых бактерий S-штамма получал новый признак - капсулу - и приобретал вирулентные свойства. Такое явление Гриффитс назвал трансформацией.
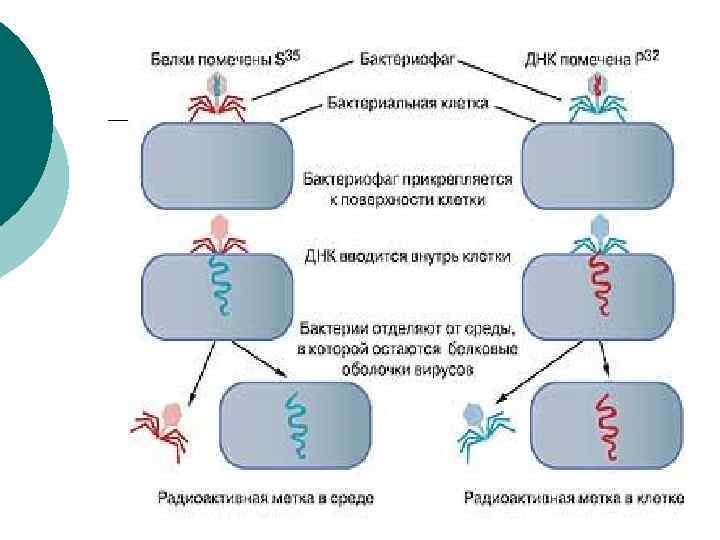
 роль ДНК в передаче наследственной информации ¡ ¡ ¡ была установлена в 1952 г. американскими вирусологами А. Д. Херши и М. Чейзом при изучении разложения фага Т 2 (вируса бактерий). Опыт состоял в том, что белки, входящие в протеиновую оболочку вириона, были помечены радиоизотопной меткой - S 35 (сера), а ДНК - радиоактивным фосфором - Р 32. В дальнейшем вирус культивировался в клетках бактерий. После этого дочерние вирионы - потомство фага - подвергались радиометрическому анализу на распределение радиоактивных меток.
роль ДНК в передаче наследственной информации ¡ ¡ ¡ была установлена в 1952 г. американскими вирусологами А. Д. Херши и М. Чейзом при изучении разложения фага Т 2 (вируса бактерий). Опыт состоял в том, что белки, входящие в протеиновую оболочку вириона, были помечены радиоизотопной меткой - S 35 (сера), а ДНК - радиоактивным фосфором - Р 32. В дальнейшем вирус культивировался в клетках бактерий. После этого дочерние вирионы - потомство фага - подвергались радиометрическому анализу на распределение радиоактивных меток.
 ¡ Исследования показали, что новое поколение фаговых частиц содержало только фосфор - Р 32. Исследователи сделали справедливый вывод о том, что именно ДНК, а не белок передается от родителей к потомству.
¡ Исследования показали, что новое поколение фаговых частиц содержало только фосфор - Р 32. Исследователи сделали справедливый вывод о том, что именно ДНК, а не белок передается от родителей к потомству.
 ¡ ¡ одна из трёх основных макромолекул (две другие") Рибонуклеи новая кисло та (РНК) ¡ ¡ одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов. Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы.
Рибонуклеи новая кисло та (РНК) ¡ ¡ одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов. Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы.
 ¡ Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
¡ Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
 ¡ РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (м. РНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице м. РНК при участии рибосом
¡ РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (м. РНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице м. РНК при участии рибосом
 ¡ малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах. ¡ Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы), у отдельных РНК обнаружена собственная ферментативная активность: способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами. ¡
¡ малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах. ¡ Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы), у отдельных РНК обнаружена собственная ферментативная активность: способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами. ¡
 ¡ Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
¡ Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
 Гипотеза РНК-мира ¡ Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира»
Гипотеза РНК-мира ¡ Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира»
 ¡ Предполагают, что РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности р. РНК и т. РНК — это реликты РНК-мира.
¡ Предполагают, что РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности р. РНК и т. РНК — это реликты РНК-мира.
 Структура гена эукариот В геноме эукариот выделяют три типа последовательностей ДНК: n Уникальные последовательности – представлены одной или несколькими копиями (60 -80% генома) n Умеренные повторы – от десятка до нескольких тысяч копий на геном (10 -20% генома) n Высокоповторяющаяся ДНК – от нескольких тысяч до миллиона копий на геном (10 -20% генома) 69
Структура гена эукариот В геноме эукариот выделяют три типа последовательностей ДНК: n Уникальные последовательности – представлены одной или несколькими копиями (60 -80% генома) n Умеренные повторы – от десятка до нескольких тысяч копий на геном (10 -20% генома) n Высокоповторяющаяся ДНК – от нескольких тысяч до миллиона копий на геном (10 -20% генома) 69
 Структура гена эукариот n Большинство функционирующих генов являются уникальными последовательностями, некоторые представлены умеренными повторами n Высокоповторная ДНК представлена очень короткими последовательностями (около 300 п. н. ) – вероятно играет структурную или регуляторную роль и локализована в основном в прицентромерном гетерохроматине 70
Структура гена эукариот n Большинство функционирующих генов являются уникальными последовательностями, некоторые представлены умеренными повторами n Высокоповторная ДНК представлена очень короткими последовательностями (около 300 п. н. ) – вероятно играет структурную или регуляторную роль и локализована в основном в прицентромерном гетерохроматине 70
 Структура гена эукариот n Большинство эукариотических генов имеет «мозаичную» экзонинтронную структуру n Экзоны – участки гена, кодирующие структуру полипептида n Интроны – участки гена, не кодирующие структуру полипептида П Э И Э И Э Т П – промотор; Э – экзоны; И – интроны; Т – терминатор 71
Структура гена эукариот n Большинство эукариотических генов имеет «мозаичную» экзонинтронную структуру n Экзоны – участки гена, кодирующие структуру полипептида n Интроны – участки гена, не кодирующие структуру полипептида П Э И Э И Э Т П – промотор; Э – экзоны; И – интроны; Т – терминатор 71
 Структура гена эукариот n Количество интрон-экзонных переходов в пределах гена может варьироваться от 0 до 50 n Колебание размеров более характерно для интронов – от 20 до более чем 10000 п. н. n Роль интронов до конца не ясна, вероятно они участвуют в процессах генетической рекомбинации, а также в процессах регуляции экспрессии 72
Структура гена эукариот n Количество интрон-экзонных переходов в пределах гена может варьироваться от 0 до 50 n Колебание размеров более характерно для интронов – от 20 до более чем 10000 п. н. n Роль интронов до конца не ясна, вероятно они участвуют в процессах генетической рекомбинации, а также в процессах регуляции экспрессии 72
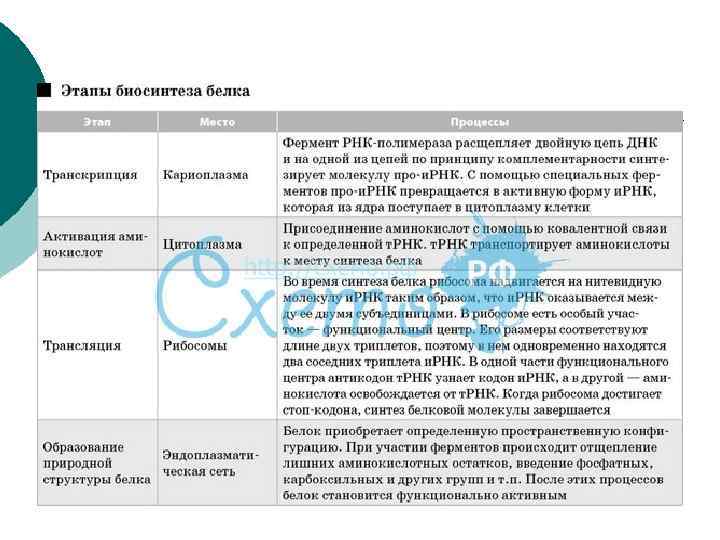
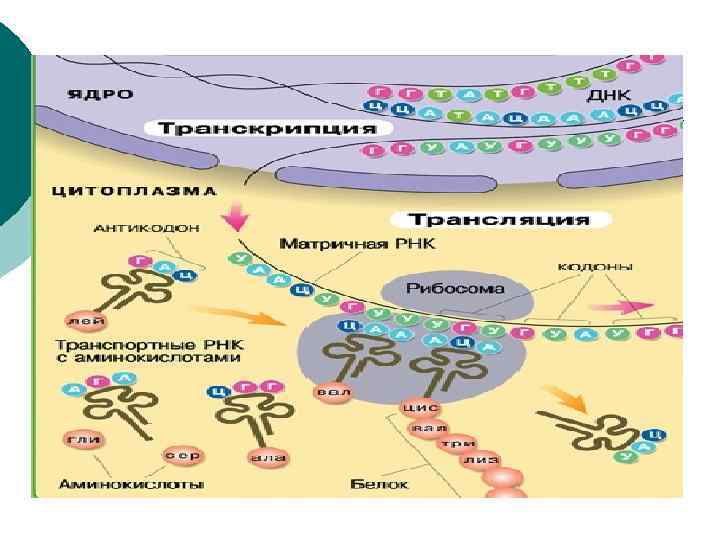
 Экспрессия генов n Экспрессия генов – реализация генетической информация, выражающаяся в процессах транскрипции и трансляции n Транскрипция – процесс переноса генетической информации с ДНК на РНК n Трансляция – биосинтез полипептидной цепи белка на матрице м. РНК 73
Экспрессия генов n Экспрессия генов – реализация генетической информация, выражающаяся в процессах транскрипции и трансляции n Транскрипция – процесс переноса генетической информации с ДНК на РНК n Трансляция – биосинтез полипептидной цепи белка на матрице м. РНК 73
 Транскрипция n Матрицей для синтеза РНК служит только одна из двух цепей ДНК, так называемая смысловая цепь (3’ → 5’) n Транскрипция происходит не на всей молекуле ДНК, а только на участке одного гена n Помимо структурных генов (несущих информацию о структуре белков), имеются гены, кодирующие р. РНК и т. РНК 74
Транскрипция n Матрицей для синтеза РНК служит только одна из двух цепей ДНК, так называемая смысловая цепь (3’ → 5’) n Транскрипция происходит не на всей молекуле ДНК, а только на участке одного гена n Помимо структурных генов (несущих информацию о структуре белков), имеются гены, кодирующие р. РНК и т. РНК 74
 Типы РНК р-РНК т-РНК 75
Типы РНК р-РНК т-РНК 75
 Стадии процесса транскрипции n Инициация n Элонгация n Терминация 76
Стадии процесса транскрипции n Инициация n Элонгация n Терминация 76
 Стадии процесса транскрипции n Инициация – начало синтеза n К промотору присоединяется комплекс ферментов, включая ДНКзависимую РНК-полимеразу n Присоединение к промотору инициирует раскручивание двойной цепочки ДНК и разрыв нуклеотидных связей 77
Стадии процесса транскрипции n Инициация – начало синтеза n К промотору присоединяется комплекс ферментов, включая ДНКзависимую РНК-полимеразу n Присоединение к промотору инициирует раскручивание двойной цепочки ДНК и разрыв нуклеотидных связей 77
 Стадии процесса транскрипции n Элонгация – последовательное присоединение свободных нуклеотидов к «смысловой цепи» ДНК по принципу комплементарности (А-У, Г-Ц) и соединение их в полирибонуклеотидную цепь при помощи РНК-полимеразы n Терминация – завершение синтеза РНК в участке терминаторе, который узнается РНК-полимеразой при участии регуляторных белковых факторов терминации 78
Стадии процесса транскрипции n Элонгация – последовательное присоединение свободных нуклеотидов к «смысловой цепи» ДНК по принципу комплементарности (А-У, Г-Ц) и соединение их в полирибонуклеотидную цепь при помощи РНК-полимеразы n Терминация – завершение синтеза РНК в участке терминаторе, который узнается РНК-полимеразой при участии регуляторных белковых факторов терминации 78
 Результат транскрипции n При экспрессии генов, кодирующих структуру белка, образуется гетерогенная ядерная РНК (гя. РНК) или пре-м. РНК, которая копирует всю нуклеотидную последовательность ДНК от промотора до терминатора, включая нетранслируемые области n Эта гя. РНК подвергается созреванию – процессингу 79
Результат транскрипции n При экспрессии генов, кодирующих структуру белка, образуется гетерогенная ядерная РНК (гя. РНК) или пре-м. РНК, которая копирует всю нуклеотидную последовательность ДНК от промотора до терминатора, включая нетранслируемые области n Эта гя. РНК подвергается созреванию – процессингу 79
 участки РНК, транскрибированные") Процессинг n Процессинг – процесс образования функционально активных м. РНК 1) участки РНК, транскрибированные с интронов, вырезаются, а участки РНК, транскрибированные с экзонов – сшиваются – сплайсинг 2) к 5’-концу гя. РНК присоединяется 7 -метил-ГТФ – кэпирование ( «кэп» - шапочка) 3) к 3’-концу присоединяется полиадениловый участок размером в 100 -250 нуклеотидов – полиаденилирование 80
Процессинг n Процессинг – процесс образования функционально активных м. РНК 1) участки РНК, транскрибированные с интронов, вырезаются, а участки РНК, транскрибированные с экзонов – сшиваются – сплайсинг 2) к 5’-концу гя. РНК присоединяется 7 -метил-ГТФ – кэпирование ( «кэп» - шапочка) 3) к 3’-концу присоединяется полиадениловый участок размером в 100 -250 нуклеотидов – полиаденилирование 80
 Структура гя. РНК и м. РНК n Структура гя. РНК: 1 – 5’-нетранслируемая область; 2 - 3’нетранслируемая область» ; 3 – копии экзонов; 4 – копии интронов 1 n 3 4 3 4 3 2 Структура м. РНК: 1 – «кэп» ; 2 – полиадениловый участок; 3 – копии экзонов 1 3 3 2 81
Структура гя. РНК и м. РНК n Структура гя. РНК: 1 – 5’-нетранслируемая область; 2 - 3’нетранслируемая область» ; 3 – копии экзонов; 4 – копии интронов 1 n 3 4 3 4 3 2 Структура м. РНК: 1 – «кэп» ; 2 – полиадениловый участок; 3 – копии экзонов 1 3 3 2 81
 раз больше м.") Результат процессинга n Обычно гя. РНК в несколько (иногда в десятки) раз больше м. РНК если гя. РНК составляет примерно 10% генома, то м. РНК – только 12% n n Функции «кэпа» заключаются в прикреплении м. РНК к рибосоме и инициации процесса трансляции – синтеза белка Функции полиаденилового хвоста – защита м. РНК от ферментативного разрушения во время транспортировки к рибосомам 82
Результат процессинга n Обычно гя. РНК в несколько (иногда в десятки) раз больше м. РНК если гя. РНК составляет примерно 10% генома, то м. РНК – только 12% n n Функции «кэпа» заключаются в прикреплении м. РНК к рибосоме и инициации процесса трансляции – синтеза белка Функции полиаденилового хвоста – защита м. РНК от ферментативного разрушения во время транспортировки к рибосомам 82
 Сплайсинг n Термин "сплайсинг" происходит от английского слова splice соединять встык n Макромолекулярный комплекс, называемый сплайсосомой, вырезает интроны и сшивает вместе экзоны n В состав сплайсосомы входят пять рибонуклеопротеинов, которые обозначают U 1, U 2, U 4, U 5 и U 6, образованные молекулами малой ядерной РНК (мя. РНК) и белками 83
Сплайсинг n Термин "сплайсинг" происходит от английского слова splice соединять встык n Макромолекулярный комплекс, называемый сплайсосомой, вырезает интроны и сшивает вместе экзоны n В состав сплайсосомы входят пять рибонуклеопротеинов, которые обозначают U 1, U 2, U 4, U 5 и U 6, образованные молекулами малой ядерной РНК (мя. РНК) и белками 83
 Альтернативный сплайсинг n Наряду с последовательным вырезанием интронов существует т. н. альтернативный сплайсинг n При альтернативном сплайсинге удаляются не только интроны, но и некоторые экзоны n В результате получается отличающийся вариант м. РНК, на основе которой в рибосоме синтезируется другой белок 84
Альтернативный сплайсинг n Наряду с последовательным вырезанием интронов существует т. н. альтернативный сплайсинг n При альтернативном сплайсинге удаляются не только интроны, но и некоторые экзоны n В результате получается отличающийся вариант м. РНК, на основе которой в рибосоме синтезируется другой белок 84
 Альтернативный сплайсинг n n Открытие альтернативного сплайсинга вновь заставляет модифицировать правило «Один ген - одна полипептидная цепь» , так как с одного гена может получаться несколько различающихся полипептидных цепей Пример альтернативного сплайсинга у человека: ген структурного белка тропомиозина дает начало пяти разным вариантам этого белка в разных тканях организма: скелетной мышце, гладкой мышце, фибробластах, печени и мозге 85
Альтернативный сплайсинг n n Открытие альтернативного сплайсинга вновь заставляет модифицировать правило «Один ген - одна полипептидная цепь» , так как с одного гена может получаться несколько различающихся полипептидных цепей Пример альтернативного сплайсинга у человека: ген структурного белка тропомиозина дает начало пяти разным вариантам этого белка в разных тканях организма: скелетной мышце, гладкой мышце, фибробластах, печени и мозге 85
 Сплайсинг 86
Сплайсинг 86
 Регуляция экспрессии генов определяет: n Согласованное действие всех элементов генома n Определение времени активации/репрессии генов n Экспрессия генов может регулироваться как на уровне транскрипции, так и посредством посттранскрипционных процессов, включая трансляцию 87
Регуляция экспрессии генов определяет: n Согласованное действие всех элементов генома n Определение времени активации/репрессии генов n Экспрессия генов может регулироваться как на уровне транскрипции, так и посредством посттранскрипционных процессов, включая трансляцию 87
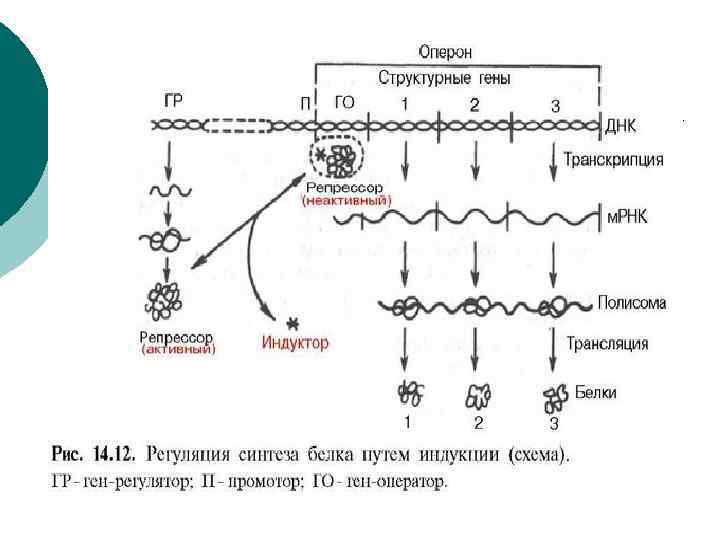
 Регуляция экспрессии генов у прокариот Практически полностью осуществляется на уровне транскрипции n Единицей транскрипции генов у прокариот является оперон n Оперон включает в себя: - промотор – участок присоединения РНК-полимеразы - оператор – участок присоединения белка-регулятора - терминатор – участок окончания синтеза генов оперона - ген-регулятор – кодирует белок регулятор Р П О С 1 С 2 С 3 Т 88
Регуляция экспрессии генов у прокариот Практически полностью осуществляется на уровне транскрипции n Единицей транскрипции генов у прокариот является оперон n Оперон включает в себя: - промотор – участок присоединения РНК-полимеразы - оператор – участок присоединения белка-регулятора - терминатор – участок окончания синтеза генов оперона - ген-регулятор – кодирует белок регулятор Р П О С 1 С 2 С 3 Т 88
 ¡ Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулироватьэкспрессию (транскрипцию ) этих генов.
¡ Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулироватьэкспрессию (транскрипцию ) этих генов.
 ¡ ¡ Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб и Моно, за что получили Нобелевскую премию в 1965 году. Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов).
¡ ¡ Концепцию оперона для прокариот предложили в 1961 году французские ученые Жакоб и Моно, за что получили Нобелевскую премию в 1965 году. Опероны по количеству цистронов делят на моно-, олиго- и полицистронные, содержащие, соответственно, только один, несколько или много цистронов (генов).
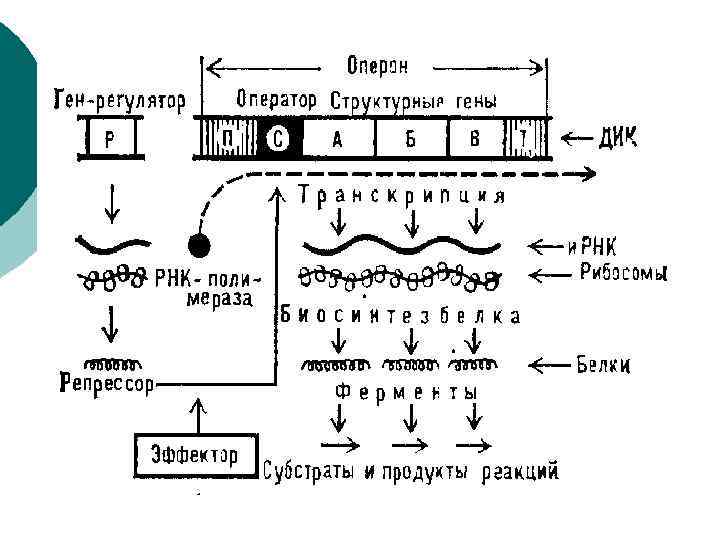
 Регуляция экспрессии генов у прокариот ¡ Негативная регуляция – связывание регуляторного белка с оператором репрессирует работу оперона ¡ Позитивная регуляция – связывание регуляторного белка с оператором активирует оперон ¡ Кроме того известны вещества – индукторы и репрессоры, которые также могут влиять на регуляторный белок (репрессировать или стимулировать) 92
Регуляция экспрессии генов у прокариот ¡ Негативная регуляция – связывание регуляторного белка с оператором репрессирует работу оперона ¡ Позитивная регуляция – связывание регуляторного белка с оператором активирует оперон ¡ Кроме того известны вещества – индукторы и репрессоры, которые также могут влиять на регуляторный белок (репрессировать или стимулировать) 92
 ¡ ¡ ¡ Характерным примером оперонной организации генома прокариот является лактозный оперон, триптофановый, пиримидиновый и bgl опероны у Escherichia coli Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор.
¡ ¡ ¡ Характерным примером оперонной организации генома прокариот является лактозный оперон, триптофановый, пиримидиновый и bgl опероны у Escherichia coli Начинается и заканчивается оперон регуляторными областями — промотором в начале и терминатором в конце, кроме этого, каждый отдельный цистрон может иметь в своей структуре собственный промотор и/или терминатор.
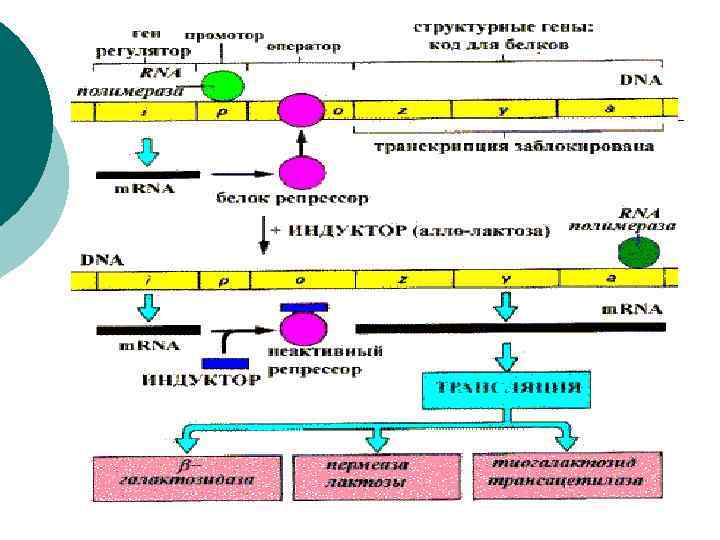
 ¡ ¡ На основании наблюдений была предложена схема регуляции оперона по механизму индукции: 1. В отсутствие лактозы активный белокрепрессор связывается с оператором и блокирует синтез м. РНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются. 2. Если лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНКполимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать м. РНК. Таким образом, лактоза является индуктором транскрипции.
¡ ¡ На основании наблюдений была предложена схема регуляции оперона по механизму индукции: 1. В отсутствие лактозы активный белокрепрессор связывается с оператором и блокирует синтез м. РНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются. 2. Если лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНКполимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать м. РНК. Таким образом, лактоза является индуктором транскрипции.
 Регуляция экспрессии генов у эукариот 1 2 П Э И Э И Т 1 2 1 – энхансеры; 2 – сайленсеры; П – промотор; Э – экзоны; И – интроны; Т – терминатор Специфическая регуляция проявляется в активации или инактивации транскрипции с отдельных генов с участием промоторов, энхансеров и сайленсеров Промоторы – участки длиной 30 -60 п. н. , для связывания с ДНК факторов транскрипции, образования комплекса ДНК-РНК полимеразы для запуска синтеза РНК 96
Регуляция экспрессии генов у эукариот 1 2 П Э И Э И Т 1 2 1 – энхансеры; 2 – сайленсеры; П – промотор; Э – экзоны; И – интроны; Т – терминатор Специфическая регуляция проявляется в активации или инактивации транскрипции с отдельных генов с участием промоторов, энхансеров и сайленсеров Промоторы – участки длиной 30 -60 п. н. , для связывания с ДНК факторов транскрипции, образования комплекса ДНК-РНК полимеразы для запуска синтеза РНК 96
 Регуляция экспрессии генов у эукариот Энхансеры – усилители транскрипции: n Могут находится как в концевых областях, так и в интронах и даже на значительных расстояниях от промоторов n Активируют гены независимо от ориентации n Один энхансер может активировать различные гены n Действие их может быть ткане- и видоспецифичным n Энхансеры доступны действию различных белков, в том числе и гормонов Сайленсеры – ослабители транскрипции 97
Регуляция экспрессии генов у эукариот Энхансеры – усилители транскрипции: n Могут находится как в концевых областях, так и в интронах и даже на значительных расстояниях от промоторов n Активируют гены независимо от ориентации n Один энхансер может активировать различные гены n Действие их может быть ткане- и видоспецифичным n Энхансеры доступны действию различных белков, в том числе и гормонов Сайленсеры – ослабители транскрипции 97
 Регуляция экспрессии генов у эукариот n Одна и та же последовательность ДНК может выступать и в роли энхансера, и в роли сайленсера в зависимости от типа клеток n Эти участки представляют собой короткие участки ДНК (100 -200 п. н. ), являющиеся местом прикрепления регуляторных белков n Взаимодействие регуляторных протеинов с этими участками изменяет активность генов вследствие изменения конформации определенного участка ДНК 98
Регуляция экспрессии генов у эукариот n Одна и та же последовательность ДНК может выступать и в роли энхансера, и в роли сайленсера в зависимости от типа клеток n Эти участки представляют собой короткие участки ДНК (100 -200 п. н. ), являющиеся местом прикрепления регуляторных белков n Взаимодействие регуляторных протеинов с этими участками изменяет активность генов вследствие изменения конформации определенного участка ДНК 98
 Регуляция экспрессии генов у эукариот Неспецифическая регуляция экспрессии n Регуляция транскрипции может осуществляться также путем конденсации-деконденсации хроматина n Регуляция экспрессии происходит на уровне процессинга – феномен альтернатиного сплайсинга n Контроль на уровне транспортировки РНК – примерно половина гя -РНК полностью распадается в ядре, не выходя за его пределы 99
Регуляция экспрессии генов у эукариот Неспецифическая регуляция экспрессии n Регуляция транскрипции может осуществляться также путем конденсации-деконденсации хроматина n Регуляция экспрессии происходит на уровне процессинга – феномен альтернатиного сплайсинга n Контроль на уровне транспортировки РНК – примерно половина гя -РНК полностью распадается в ядре, не выходя за его пределы 99
 Регуляция экспрессии генов у эукариот Неспецифическая регуляция экспрессии n Контроль на уровне стабилизации м-РНК – избирательная стабилизация определенных типов м-РНК в цитоплазме, которые не подвергаются распаду после трансляции n Контроль на уровне трансляции – отбор определенных м-РНК для трансляции на рибосомах n Контроль на уровне пострансляционной модификации белка 100
Регуляция экспрессии генов у эукариот Неспецифическая регуляция экспрессии n Контроль на уровне стабилизации м-РНК – избирательная стабилизация определенных типов м-РНК в цитоплазме, которые не подвергаются распаду после трансляции n Контроль на уровне трансляции – отбор определенных м-РНК для трансляции на рибосомах n Контроль на уровне пострансляционной модификации белка 100
 Генети ческий код ¡ ¡ свойственный всем живым организмам способ кодирования аминокислотно йпоследовательности белков при помощи последовательности нуклеотидов. В ДНК используется четыре азотистых основания — аденин (А), гуанин (G), цитозин (С), тимин (T), которые в русскоязычной литературе обозначаются буквами А, Г, Ц и Т. Эти буквы составляют алфавит генетическогокода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом — урацилом, который обозначается буквой U (У в русскоязычной литературе).
Генети ческий код ¡ ¡ свойственный всем живым организмам способ кодирования аминокислотно йпоследовательности белков при помощи последовательности нуклеотидов. В ДНК используется четыре азотистых основания — аденин (А), гуанин (G), цитозин (С), тимин (T), которые в русскоязычной литературе обозначаются буквами А, Г, Ц и Т. Эти буквы составляют алфавит генетическогокода. В РНК используются те же нуклеотиды, за исключением тимина, который заменён похожим нуклеотидом — урацилом, который обозначается буквой U (У в русскоязычной литературе).
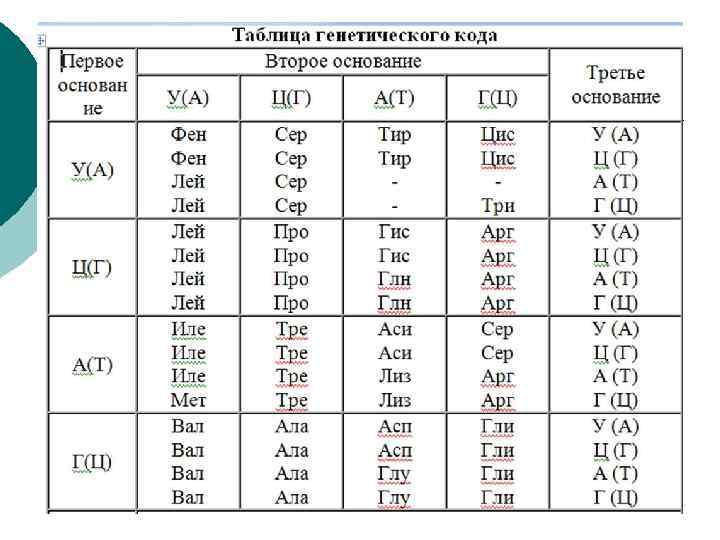
 Свойства ¡ ¡ Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий ибактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1]
Свойства ¡ ¡ Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий ибактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1]
 — одной и той же аминокислоте может соответствовать несколько") ¡ ¡ ¡ Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже). Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
¡ ¡ ¡ Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже). Помехоустойчивость — мутации замен нуклеотидов, не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.
 Генная инженерия
Генная инженерия
 Генетическая инжене рия (генная инженерия ¡ ¡ совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы. Генетическая инженерия не является наукой в широком смысле, но является инструментом биотехнологии, используя методы таких биологических наук, как молекулярная и клеточная биология, цитология, генетика, микробиология, вирусология.
Генетическая инжене рия (генная инженерия ¡ ¡ совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами и введения их в другие организмы. Генетическая инженерия не является наукой в широком смысле, но является инструментом биотехнологии, используя методы таких биологических наук, как молекулярная и клеточная биология, цитология, генетика, микробиология, вирусология.
 ¡ Генная инженерия служит для получения желаемых качеств изменяемого или генетически модифицированного организма. В отличие от традиционной селекции, в ходе которой генотип подвергается изменениям лишь косвенно, генная инженерия позволяет непосредственно вмешиваться в генетический аппарат, применяя технику молекулярного клонирования.
¡ Генная инженерия служит для получения желаемых качеств изменяемого или генетически модифицированного организма. В отличие от традиционной селекции, в ходе которой генотип подвергается изменениям лишь косвенно, генная инженерия позволяет непосредственно вмешиваться в генетический аппарат, применяя технику молекулярного клонирования.
 ¡ Примерами применения генной инженерии являются получение новых генетически модифицированных сортов зерновых культур, производство человеческого инсулина путём использования генномодифицированных бактерий, производство эритропоэтина в культуре клеток или новых пород экспериментальных мышей для научных исследований.
¡ Примерами применения генной инженерии являются получение новых генетически модифицированных сортов зерновых культур, производство человеческого инсулина путём использования генномодифицированных бактерий, производство эритропоэтина в культуре клеток или новых пород экспериментальных мышей для научных исследований.
 ¡ Основные этапы решения генноинженерной задачи следующие: l l l 1. Получение изолированного гена. 2. Введение гена в вектор для переноса в организм. 3. Перенос вектора с геном в модифицируемый организм. 4. Преобразование клеток организма. 5. Отбор генетически модифицированных организмов (ГМО) и устранение тех, которые не были успешно модифицированы.
¡ Основные этапы решения генноинженерной задачи следующие: l l l 1. Получение изолированного гена. 2. Введение гена в вектор для переноса в организм. 3. Перенос вектора с геном в модифицируемый организм. 4. Преобразование клеток организма. 5. Отбор генетически модифицированных организмов (ГМО) и устранение тех, которые не были успешно модифицированы.
 — стойкое (то есть такое, которое") Мутации ¡ Мута ция (лат. mutatio — изменение) — стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) изменение генотипа, происходящее под влиянием внешней или внутренней среды. Термин предложен Гуго де Фризом. Процесс возникновения мутаций получил название мутагенеза.
Мутации ¡ Мута ция (лат. mutatio — изменение) — стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) изменение генотипа, происходящее под влиянием внешней или внутренней среды. Термин предложен Гуго де Фризом. Процесс возникновения мутаций получил название мутагенеза.

 Классификации мутаций ¡ ¡ В современной учебной литературе используется формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций: геномные; хромосомные; генные.
Классификации мутаций ¡ ¡ В современной учебной литературе используется формальная классификация, основанная на характере изменения структуры отдельных генов, хромосом и генома в целом. В рамках этой классификации различают следующие виды мутаций: геномные; хромосомные; генные.
 ¡ Геномные: — полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3 n, 4 n, 6 n и т. д. ) наборами хромосом) и анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору
¡ Геномные: — полиплоидизация (образование организмов или клеток, геном которых представлен более чем двумя (3 n, 4 n, 6 n и т. д. ) наборами хромосом) и анеуплоидия (гетероплоидия) — изменение числа хромосом, не кратное гаплоидному набору
 ¡ При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация)
¡ При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация)
 ¡ На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях
¡ На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точечных мутациях
 Последствия мутаций для клетки и организма ¡ ¡ Мутации, которые ухудшают деятельность клетки в многоклеточном организме, часто приводят к уничтожению клетки (в частности, к программируемой смерти клетки, — апоптозу). Если внутри- и внеклеточные защитные механизмы не распознали мутацию и клетка прошла деление, то мутантный ген передастся всем потомкам клетки. Мутация в соматической клетке сложного многоклеточного организма может привести к злокачественным или доброкачественным новообразованиям, мутация в половой клетке — к изменению свойств всего организма-потомка.
Последствия мутаций для клетки и организма ¡ ¡ Мутации, которые ухудшают деятельность клетки в многоклеточном организме, часто приводят к уничтожению клетки (в частности, к программируемой смерти клетки, — апоптозу). Если внутри- и внеклеточные защитные механизмы не распознали мутацию и клетка прошла деление, то мутантный ген передастся всем потомкам клетки. Мутация в соматической клетке сложного многоклеточного организма может привести к злокачественным или доброкачественным новообразованиям, мутация в половой клетке — к изменению свойств всего организма-потомка.
 Роль мутаций в эволюции ¡ При существенном изменении условий существования те мутации, которые раньше были вредными, могут оказаться полезными. Таким образом, мутации являются материалом для естественного отбора.
Роль мутаций в эволюции ¡ При существенном изменении условий существования те мутации, которые раньше были вредными, могут оказаться полезными. Таким образом, мутации являются материалом для естественного отбора.
