

Этапы реализации генетической информации.ppt
- Количество слайдов: 32
 Этапы реализации генетической информации в клетке
Этапы реализации генетической информации в клетке
 как в пределах одной") Важным свойством генетической информации является ее способность к переносу (передаче) как в пределах одной клетки, так и от родительской клетки к дочерним либо между клетками разных индивидуумов в процессах клеточного деления и размножения организмов
Важным свойством генетической информации является ее способность к переносу (передаче) как в пределах одной клетки, так и от родительской клетки к дочерним либо между клетками разных индивидуумов в процессах клеточного деления и размножения организмов
 § репликация молекул ДНК,") направления внутриклеточного переноса генетической информации ( в случае ДНКсодержащих организмов) § репликация молекул ДНК, т. е. копирование информации § синтез молекул РНК (транскрипция ) § образование полипептидов (трансляция) каждый из указанных процессов осуществляется на основе принципов матричности и комплементарности.
направления внутриклеточного переноса генетической информации ( в случае ДНКсодержащих организмов) § репликация молекул ДНК, т. е. копирование информации § синтез молекул РНК (транскрипция ) § образование полипептидов (трансляция) каждый из указанных процессов осуществляется на основе принципов матричности и комплементарности.
 Репликация ДНК прокариот и эукариот
Репликация ДНК прокариот и эукариот
 Основные направления внутриклеточного переноса генетической информации.
Основные направления внутриклеточного переноса генетической информации.
 ДНК → РНК→ белок «центральная догма» молекулярной биологии «общий перенос»
ДНК → РНК→ белок «центральная догма» молекулярной биологии «общий перенос»
 «специализированный перенос» у РНК-содержащих вирусов. В этом случае наблюдается процесс, получивший название обратной транскрипции, при котором первичный генетический материал (вирусная РНК), проникший в клетку-хозяина, служит матрицей для синтеза комплементарной ДНК с помощью фермента обратной транскриптазы (ревертазы), кодируемой вирусным геномом. В дальнейшем возможна реализация информации синтезированной вирусной ДНК в обычном направлении. Специализированный перенос генетической информации осуществляется по схеме РНК→ДНК→ РНК → белок.
«специализированный перенос» у РНК-содержащих вирусов. В этом случае наблюдается процесс, получивший название обратной транскрипции, при котором первичный генетический материал (вирусная РНК), проникший в клетку-хозяина, служит матрицей для синтеза комплементарной ДНК с помощью фермента обратной транскриптазы (ревертазы), кодируемой вирусным геномом. В дальнейшем возможна реализация информации синтезированной вирусной ДНК в обычном направлении. Специализированный перенос генетической информации осуществляется по схеме РНК→ДНК→ РНК → белок.
 Транскрипция является первым этапом общего переноса генетической информации и представляет собой процесс биосинтеза молекул РНК по программе ДНК. Принципиальный смысл этого процесса состоит в том, что информация структурного гена (либо нескольких расположенных рядом генов), записанная в форме нуклеотидной последовательности кодирующей нити ДНК в ориентации 3'→ 5', переписывается (транскрибируется) в нуклеотидную последовательность молекулы РНК, синтезируемой в направлении 5'→ 3' на основе комплементарного соответствия дезоксирибонуклеотидов матричной нити ДНК рибонуклеотидам РНК (А-У, Г-Ц, Т-А, Ц-Г). В качестве продуктов транскрипции (транскриптов) можно рассматривать все типы молекул РНК, участвующих в биосинтезе белков в клетке, — матричные (информационные) РНК (м. РНК, или и. РНК), рибосомные РНК (р. РНК), транспортные РНК (т. РНК), малые ядерные РНК (мя. РНК).
Транскрипция является первым этапом общего переноса генетической информации и представляет собой процесс биосинтеза молекул РНК по программе ДНК. Принципиальный смысл этого процесса состоит в том, что информация структурного гена (либо нескольких расположенных рядом генов), записанная в форме нуклеотидной последовательности кодирующей нити ДНК в ориентации 3'→ 5', переписывается (транскрибируется) в нуклеотидную последовательность молекулы РНК, синтезируемой в направлении 5'→ 3' на основе комплементарного соответствия дезоксирибонуклеотидов матричной нити ДНК рибонуклеотидам РНК (А-У, Г-Ц, Т-А, Ц-Г). В качестве продуктов транскрипции (транскриптов) можно рассматривать все типы молекул РНК, участвующих в биосинтезе белков в клетке, — матричные (информационные) РНК (м. РНК, или и. РНК), рибосомные РНК (р. РНК), транспортные РНК (т. РНК), малые ядерные РНК (мя. РНК).
 Синтез молекулы РНК на матричной нити ДНК. Стрелкой показано направление, в котором идет рост цепи РНК
Синтез молекулы РНК на матричной нити ДНК. Стрелкой показано направление, в котором идет рост цепи РНК
 § § § Процесс транскрипции обеспечивается комплексным действием ряда ферментов, к числу которых относится РНК-полимераза, представляющая собой сложный белок, состоящий из нескольких субъединиц и способный выполнять несколько функций. В отличие от прокариот (бактерий), в клетках которых имеется РНК-полимераза лишь одного типа, обеспечивающая синтез разных молекул РНК, у эукариот установлено наличие ядерных РНК-полимераз трех типов (I, П, III), а также РНК-полимераз клеточных органелл, содержащих ДНК (митохондрий, пластид). РНК-полимераза I находится в ядрышке и участвует в синтезе большинства молекул р. РНК, РНК-полимераза II обеспечивает синтез м. РНК и мя. РНК, РНК-полимераза III осуществляет синтез т. РНК и одного варианта молекул р. РНК.
§ § § Процесс транскрипции обеспечивается комплексным действием ряда ферментов, к числу которых относится РНК-полимераза, представляющая собой сложный белок, состоящий из нескольких субъединиц и способный выполнять несколько функций. В отличие от прокариот (бактерий), в клетках которых имеется РНК-полимераза лишь одного типа, обеспечивающая синтез разных молекул РНК, у эукариот установлено наличие ядерных РНК-полимераз трех типов (I, П, III), а также РНК-полимераз клеточных органелл, содержащих ДНК (митохондрий, пластид). РНК-полимераза I находится в ядрышке и участвует в синтезе большинства молекул р. РНК, РНК-полимераза II обеспечивает синтез м. РНК и мя. РНК, РНК-полимераза III осуществляет синтез т. РНК и одного варианта молекул р. РНК.
 Транскрипция РНК.
Транскрипция РНК.
, • элонгацию (удлинение") Транскрипция подразделяется на три основные стадии: • инициацию (начало синтеза РНК), • элонгацию (удлинение полинуклеотидной цепочки), • терминацию (окончание процесса).
Транскрипция подразделяется на три основные стадии: • инициацию (начало синтеза РНК), • элонгацию (удлинение полинуклеотидной цепочки), • терминацию (окончание процесса).
 Инициация транскрипции зависит от предварительного специфического связывания РНК-полимеразы с узнаваемой ею короткой нуклеотидной последовательностью в участке молекулы ДНК (промоторе), расположенном перед стартовой точкой структурного гена, с которой начинается синтез РНК. Промоторы разных структурных генов могут быть идентичными либо содержат отличающиеся друг от друга последовательности нуклеотидов, что, вероятно, определяет эффективность транскрибирования отдельных генов и возможности регуляции самого процесса транскрипции. Промоторы многих генов прокариот имеют в своем составе универсальную последовательность 5'ТАТААТ-З' (блок Прибнова), которая располагается перед стартовой точкой на расстоянии порядка 10 нуклеотидов и распознается РНК-полимеразой. Другая относительно часто встречающаяся узнаваемая последовательность этих организмов (5'-ТТГАЦА-3') обычно обнаруживается на расстоянии примерно 35 нуклеотидов от стартовой точки. В геномах эукариот функцию узнавания для РНК-полимеразы II могут выполнять универсальные последовательности ТАТА (блок Хогнесса), ЦААТ и состоящие из повторяющихся нуклеотидов Г и Ц (ГЦ-мотивы). При этом та или иная промоторная область может содержать либо одну из указанных последовательностей либо комбинацию двух или трех таких последовательностей.
Инициация транскрипции зависит от предварительного специфического связывания РНК-полимеразы с узнаваемой ею короткой нуклеотидной последовательностью в участке молекулы ДНК (промоторе), расположенном перед стартовой точкой структурного гена, с которой начинается синтез РНК. Промоторы разных структурных генов могут быть идентичными либо содержат отличающиеся друг от друга последовательности нуклеотидов, что, вероятно, определяет эффективность транскрибирования отдельных генов и возможности регуляции самого процесса транскрипции. Промоторы многих генов прокариот имеют в своем составе универсальную последовательность 5'ТАТААТ-З' (блок Прибнова), которая располагается перед стартовой точкой на расстоянии порядка 10 нуклеотидов и распознается РНК-полимеразой. Другая относительно часто встречающаяся узнаваемая последовательность этих организмов (5'-ТТГАЦА-3') обычно обнаруживается на расстоянии примерно 35 нуклеотидов от стартовой точки. В геномах эукариот функцию узнавания для РНК-полимеразы II могут выполнять универсальные последовательности ТАТА (блок Хогнесса), ЦААТ и состоящие из повторяющихся нуклеотидов Г и Ц (ГЦ-мотивы). При этом та или иная промоторная область может содержать либо одну из указанных последовательностей либо комбинацию двух или трех таких последовательностей.
 Специфическое прочное связывание РНК-полимеразы с тем или иным узнаваемым ею участком промоторной области позволяет ей начать процесс расплетания молекулы ДНК вплоть до стартовой точки, с которой она начинает осуществлять полимеризацию рибо нуклеотидов с использованием в качестве матрицы однонитевого 3'-5'-фрагмента ДНК. Дальнейшее расплетание ДНК структурного гена сопровождается удлинением синтезируемого полирибонуклеотида (элонгацией нити РНК), продолжающимся вплоть до достижения РНК-полимеразой области терминатора. Последний представляет собой нуклеотидную последовательность ДНК, которая узнается РНК-полимеразой при участии других белковых факторов терминации, что приводит к окончанию синтеза транскрипта и его отсоединению от матрицы. В большинстве случаев терминатор находится в конце структурного гена, обеспечивая синтез одной моногенной молекулы м. РНК. При этом у прокариот возможен синтез полигенной молекулы м. РНК, кодирующей синтез двух и большего числа полипептидных цепочек. Происходит непрерывное транскрибирование нескольких расположенных рядом друг с другом структурных генов, имеющих один общий терминатор. Полигенная м. РНК может содержать в своем составе нетранслирусмые межгенные области (спейсеры), разделяющие кодирующие участки для отдельных полипептидов, что, вероятно, обеспечивает последующее разделение и самих синтезируемых полипептидов. Поскольку структурные гены эукариот имеют прерывистое (мозаичное) строение, то их транскрипция имеет специфические особенности, отличающие ее от транскрипции у прокариот. В случае эукариотического гена, кодирующего синтез полипептида, этот процесс начинается с транскрибирования всей нуклеотидной последовательности, содержащей как экзонные, так и интронные участки ДНК. Образовавшаяся при этом молекула м. РНК, отражающая структуру всего мозаичного гена, которую называют гетерогенной ядерной РНК (гя. РНК) либо проматричной РНК (про-м. РНК), претерпевает затем процесс созревания (процессинг м. РНК).
Специфическое прочное связывание РНК-полимеразы с тем или иным узнаваемым ею участком промоторной области позволяет ей начать процесс расплетания молекулы ДНК вплоть до стартовой точки, с которой она начинает осуществлять полимеризацию рибо нуклеотидов с использованием в качестве матрицы однонитевого 3'-5'-фрагмента ДНК. Дальнейшее расплетание ДНК структурного гена сопровождается удлинением синтезируемого полирибонуклеотида (элонгацией нити РНК), продолжающимся вплоть до достижения РНК-полимеразой области терминатора. Последний представляет собой нуклеотидную последовательность ДНК, которая узнается РНК-полимеразой при участии других белковых факторов терминации, что приводит к окончанию синтеза транскрипта и его отсоединению от матрицы. В большинстве случаев терминатор находится в конце структурного гена, обеспечивая синтез одной моногенной молекулы м. РНК. При этом у прокариот возможен синтез полигенной молекулы м. РНК, кодирующей синтез двух и большего числа полипептидных цепочек. Происходит непрерывное транскрибирование нескольких расположенных рядом друг с другом структурных генов, имеющих один общий терминатор. Полигенная м. РНК может содержать в своем составе нетранслирусмые межгенные области (спейсеры), разделяющие кодирующие участки для отдельных полипептидов, что, вероятно, обеспечивает последующее разделение и самих синтезируемых полипептидов. Поскольку структурные гены эукариот имеют прерывистое (мозаичное) строение, то их транскрипция имеет специфические особенности, отличающие ее от транскрипции у прокариот. В случае эукариотического гена, кодирующего синтез полипептида, этот процесс начинается с транскрибирования всей нуклеотидной последовательности, содержащей как экзонные, так и интронные участки ДНК. Образовавшаяся при этом молекула м. РНК, отражающая структуру всего мозаичного гена, которую называют гетерогенной ядерной РНК (гя. РНК) либо проматричной РНК (про-м. РНК), претерпевает затем процесс созревания (процессинг м. РНК).
 с последующим удалением его интронных") Процессинг состоит в ферментативном разрезании первичного транскрипта (гя. РНК) с последующим удалением его интронных участков и воссоединением (сплайсингом) экзонных участков, формирующих непрерывную кодирующую последовательность зрелой м. РНК, которая в дальнейшем участвует в трансляции генетической информации. Процессинг м. РНК β-глобинового гена человека.
Процессинг состоит в ферментативном разрезании первичного транскрипта (гя. РНК) с последующим удалением его интронных участков и воссоединением (сплайсингом) экзонных участков, формирующих непрерывную кодирующую последовательность зрелой м. РНК, которая в дальнейшем участвует в трансляции генетической информации. Процессинг м. РНК β-глобинового гена человека.
 Роль малой ядерной РНК в процессе транскрипции.
Роль малой ядерной РНК в процессе транскрипции.
 Трансляция как очередной этап реализации генетической информации заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула м. РНК (считывание информации в направлении 5'→ 3'). Следует заметить, что в клетках прокариот, не имеющих настоящего ядра с оболочкой, хромосомный генетический материал (ДНК) практически находится в цитоплазме, что определяет непрерывный характер взаимосвязи процессов транскрипции и трансляции. Иными словами, образовавшийся лидирующий 5'-конец молекулы м. РНК, синтез которой еще не завершен, уже способен вступать в контакт с рибосомой, инициируя синтез полипептида, т. е. транскрипция и трансляция идут одновременно. Что касается эукариот, то процессы транскрипции их ядерной генетической информации и ее трансляции должны быть разделены во времени в связи с процессингом молекул РНК и необходимостью их последующей упаковки и транспортировки из кариоплазмы в цитоплазму с участием специальных транспортных белков.
Трансляция как очередной этап реализации генетической информации заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула м. РНК (считывание информации в направлении 5'→ 3'). Следует заметить, что в клетках прокариот, не имеющих настоящего ядра с оболочкой, хромосомный генетический материал (ДНК) практически находится в цитоплазме, что определяет непрерывный характер взаимосвязи процессов транскрипции и трансляции. Иными словами, образовавшийся лидирующий 5'-конец молекулы м. РНК, синтез которой еще не завершен, уже способен вступать в контакт с рибосомой, инициируя синтез полипептида, т. е. транскрипция и трансляция идут одновременно. Что касается эукариот, то процессы транскрипции их ядерной генетической информации и ее трансляции должны быть разделены во времени в связи с процессингом молекул РНК и необходимостью их последующей упаковки и транспортировки из кариоплазмы в цитоплазму с участием специальных транспортных белков.
 § § § трансляция подразделяется на три основные стадии: инициацию элонгацию терминацию.
§ § § трансляция подразделяется на три основные стадии: инициацию элонгацию терминацию.
, которая может участвовать в синтезе") специфичность структурной организации группы идентичных рибосом (полирибосомы, или полисомы), которая может участвовать в синтезе первичной структуры определенной белковой молекулы (полипептида), кодируемой соответствующей м. РНК. Как известно, отдельная рибосома представляет собой клеточную органеллу, состоящую из молекул р. РНК, которые определяют ее специфичность, и из белков. В составе рибосомы имеются 2 структурные субъединицы (большая и малая), которые можно дифференцировать на основании их способности поразному осаждаться при ультрацентрифугировании препаратов очищенных рибосом из разрушенных клеток, т. е. по коэффициенту седиментации (величине S). При определенных условиях в клетке может происходить разделение (диссоциация) этих двух субъединиц либо их объединение (ассоциация). Рибосомы прокариот, а также митохондрий и хлоропластов состоят из большой и малой субъединиц с величинами 50 S и 30 S соответственно, тогда как у эукариот эти субъединицы имеют другие размеры (60 S и 40 S). Рибосома содержит 2 участка, имеющих прямое отношение к инициации трансляции, обозначенные как А-участок (аминоацильный) и P-участок (пептидильный), специфичность которых определяется сочетанием соответствующих областей субъединиц 50 S и 30 S. При диссоциации субъединиц рибосомы эти участки становятся «недостроенными» , что приводит к изменению их функциональной специфичности.
специфичность структурной организации группы идентичных рибосом (полирибосомы, или полисомы), которая может участвовать в синтезе первичной структуры определенной белковой молекулы (полипептида), кодируемой соответствующей м. РНК. Как известно, отдельная рибосома представляет собой клеточную органеллу, состоящую из молекул р. РНК, которые определяют ее специфичность, и из белков. В составе рибосомы имеются 2 структурные субъединицы (большая и малая), которые можно дифференцировать на основании их способности поразному осаждаться при ультрацентрифугировании препаратов очищенных рибосом из разрушенных клеток, т. е. по коэффициенту седиментации (величине S). При определенных условиях в клетке может происходить разделение (диссоциация) этих двух субъединиц либо их объединение (ассоциация). Рибосомы прокариот, а также митохондрий и хлоропластов состоят из большой и малой субъединиц с величинами 50 S и 30 S соответственно, тогда как у эукариот эти субъединицы имеют другие размеры (60 S и 40 S). Рибосома содержит 2 участка, имеющих прямое отношение к инициации трансляции, обозначенные как А-участок (аминоацильный) и P-участок (пептидильный), специфичность которых определяется сочетанием соответствующих областей субъединиц 50 S и 30 S. При диссоциации субъединиц рибосомы эти участки становятся «недостроенными» , что приводит к изменению их функциональной специфичности.
 Полирибосома.
Полирибосома.
 Строение бактериальной рибосомы: P – пептидильный участок, A – аминоациальный участок.
Строение бактериальной рибосомы: P – пептидильный участок, A – аминоациальный участок.
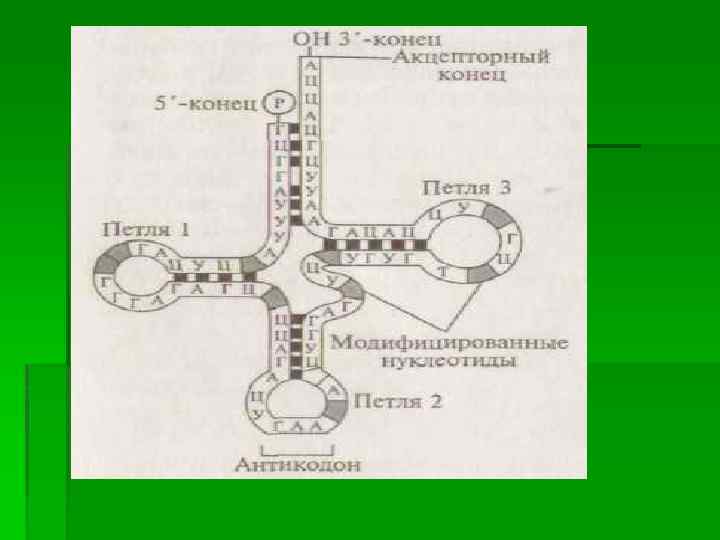
 В процессе трансляции участвуют также молекулы т. РНК, функции которых состоят в транспортировке аминокислот из цитозоля (цитоплазматического раствора) к рибосомам. Молекула т. РНК, имеющая вторичную структуру в форме «клеверного листа» , содержит в своем составе тройку нуклеотидов (антикодон), которая обеспечивает ее комплементарное соединение с соответствующим кодоном (триплетом) молекулы м. РНК, кодирующей синтез полипептида на рибосоме, и акцепторный участок (на 3'конце молекулы), к которому присоединяется определенная аминокислота. Процесс присоединения каждой из 20 аминокислот к акцепторному концу соответствующей т. РНК связан с ее активацией определенным вариантом фермента аминоацил-т. РНК-синтетазы с использованием энергии аденозинтрифосфатов (молекул АТФ). Образовавшийся при этом специфический комплекс т. РНК и аминокислоты, который получил название аминоацил-т. РНК, перемещается затем к рибосоме и участвует в синтезе полипептида.
В процессе трансляции участвуют также молекулы т. РНК, функции которых состоят в транспортировке аминокислот из цитозоля (цитоплазматического раствора) к рибосомам. Молекула т. РНК, имеющая вторичную структуру в форме «клеверного листа» , содержит в своем составе тройку нуклеотидов (антикодон), которая обеспечивает ее комплементарное соединение с соответствующим кодоном (триплетом) молекулы м. РНК, кодирующей синтез полипептида на рибосоме, и акцепторный участок (на 3'конце молекулы), к которому присоединяется определенная аминокислота. Процесс присоединения каждой из 20 аминокислот к акцепторному концу соответствующей т. РНК связан с ее активацией определенным вариантом фермента аминоацил-т. РНК-синтетазы с использованием энергии аденозинтрифосфатов (молекул АТФ). Образовавшийся при этом специфический комплекс т. РНК и аминокислоты, который получил название аминоацил-т. РНК, перемещается затем к рибосоме и участвует в синтезе полипептида.
 Синтез белка.
Синтез белка.
 Инициация трансляции обеспечивается точным соединением лидирующего 5'-конца молекулы м. РНК с определенной областью малой субъединицы диссоциированной рибосомы таким образом, что в «недостроенном» Р-участке оказывается стартовый (инициирующий) кодон АУГ этой молекулы. Функциональная особенность такого Р-участка состоит в том, что он может быть занят только инициирующей аминоацил-т. РНК с антикодоном УАЦ, которая у эукариот несет аминокислоту метионин, а у бактерий формилметионин. Поскольку синтез полипептида всегда начинается с N-конца и нарастает в направлении к С-концу, то все белковые молекулы, синтезируемые в клетках прокариот, должны начинаться с УУ-формилметионина, а у эукариот - с N -метионина. Однако, в дальнейшем эти аминокислоты ферментативно выщепляются во время процессинга белковой молекулы. § После образования инициирующего комплекса в «недостроенном» Р -участке становится возможным воссоединение малой и большой субъединиц рибосомы, что приводит к «достраиванию» Р-участка и А-участка. Лишь после этого следующая аминоацил-т. РНК может занимать А-участок на основе принципа комплементарности ее антикодона соответствующему кодону м. РНК, находящемуся в этом участке.
Инициация трансляции обеспечивается точным соединением лидирующего 5'-конца молекулы м. РНК с определенной областью малой субъединицы диссоциированной рибосомы таким образом, что в «недостроенном» Р-участке оказывается стартовый (инициирующий) кодон АУГ этой молекулы. Функциональная особенность такого Р-участка состоит в том, что он может быть занят только инициирующей аминоацил-т. РНК с антикодоном УАЦ, которая у эукариот несет аминокислоту метионин, а у бактерий формилметионин. Поскольку синтез полипептида всегда начинается с N-конца и нарастает в направлении к С-концу, то все белковые молекулы, синтезируемые в клетках прокариот, должны начинаться с УУ-формилметионина, а у эукариот - с N -метионина. Однако, в дальнейшем эти аминокислоты ферментативно выщепляются во время процессинга белковой молекулы. § После образования инициирующего комплекса в «недостроенном» Р -участке становится возможным воссоединение малой и большой субъединиц рибосомы, что приводит к «достраиванию» Р-участка и А-участка. Лишь после этого следующая аминоацил-т. РНК может занимать А-участок на основе принципа комплементарности ее антикодона соответствующему кодону м. РНК, находящемуся в этом участке.
 начальные этапы трансляции: а – инициирующий комплекс; б – элонгация
начальные этапы трансляции: а – инициирующий комплекс; б – элонгация
 Синтез белка у прокариот и эукариот.
Синтез белка у прокариот и эукариот.
 и последующей") Процесс элонгации начинается с образования пептидной связи между инициирующей (первой в цепочке) и последующей (второй) аминокислотами. Затем происходит перемещение рибосомы на один триплет м. РНК в направлении 5'-» 3', что сопровождается отсоединением инициирующей т. РНК от матрицы (м. РНК), от инициирующей аминокислоты и выходом ее в цитоплазму. При этом вторая по счету аминоацил-т. РНК передвигается из А-участка в Р-участок, а освободившийся А-участок занимается следующей (третьей по счету) аминоацилт. РНК. Процесс последовательного передвижения рибосомы «триплетными шагами» по нити м. РНК повторяется, сопровождаясь освобождением т. РНК, поступающих в Р-участок, и наращиванием аминокислотной последовательности синтезируемого полипептида. Терминация трансляции связана с вхождением одного из трех известных стоп-триплетов м. РНК в А-участок рибосомы. Поскольку такой триплет не несет информации о какой-либо аминокислоте, но узнается соответствующими белками терминации, то процесс синтеза полипептида прекращается и он отсоединяется от матрицы (м. РНК). После выхода из функционирующей рибосомы свободный 5'-конец м. РНК может вступать в контакт со следующей рибосомой полисомной группы, инициируя синтез еще одного (идентичного) полипептида. Следовательно, рассмотренный рибосомный цикл последовательно повторяется с участием нескольких рибосом одной и той же полисомы, в результате чего синтезируется группа идентичных полипептидов.
Процесс элонгации начинается с образования пептидной связи между инициирующей (первой в цепочке) и последующей (второй) аминокислотами. Затем происходит перемещение рибосомы на один триплет м. РНК в направлении 5'-» 3', что сопровождается отсоединением инициирующей т. РНК от матрицы (м. РНК), от инициирующей аминокислоты и выходом ее в цитоплазму. При этом вторая по счету аминоацил-т. РНК передвигается из А-участка в Р-участок, а освободившийся А-участок занимается следующей (третьей по счету) аминоацилт. РНК. Процесс последовательного передвижения рибосомы «триплетными шагами» по нити м. РНК повторяется, сопровождаясь освобождением т. РНК, поступающих в Р-участок, и наращиванием аминокислотной последовательности синтезируемого полипептида. Терминация трансляции связана с вхождением одного из трех известных стоп-триплетов м. РНК в А-участок рибосомы. Поскольку такой триплет не несет информации о какой-либо аминокислоте, но узнается соответствующими белками терминации, то процесс синтеза полипептида прекращается и он отсоединяется от матрицы (м. РНК). После выхода из функционирующей рибосомы свободный 5'-конец м. РНК может вступать в контакт со следующей рибосомой полисомной группы, инициируя синтез еще одного (идентичного) полипептида. Следовательно, рассмотренный рибосомный цикл последовательно повторяется с участием нескольких рибосом одной и той же полисомы, в результате чего синтезируется группа идентичных полипептидов.
 § Посттрансляционная модификация полипептида представляет собой завершающий этап реализации генетической информации в клетке, приводящий к превращению синтезированного полипептида в функционально активную молекулу белка. При этом первичный полипептид может претерпевать процессинг, состоящий в ферментативном удалении инициирующих аминокислот, отщеплении других (ненужных) аминокислотных остатков и в химической модификации отдельных аминокислот. Затем происходит процесс сворачивания линейной структуры полипептида за счет образования дополнительных связей между отдельными аминокислотами и формирование вторичной структуры белковой молекулы. На этой основе формируется еще более сложная третичная структура молекулы.
§ Посттрансляционная модификация полипептида представляет собой завершающий этап реализации генетической информации в клетке, приводящий к превращению синтезированного полипептида в функционально активную молекулу белка. При этом первичный полипептид может претерпевать процессинг, состоящий в ферментативном удалении инициирующих аминокислот, отщеплении других (ненужных) аминокислотных остатков и в химической модификации отдельных аминокислот. Затем происходит процесс сворачивания линейной структуры полипептида за счет образования дополнительных связей между отдельными аминокислотами и формирование вторичной структуры белковой молекулы. На этой основе формируется еще более сложная третичная структура молекулы.
 § Вторичная структура молекулы фермента рибонуклеазы.
§ Вторичная структура молекулы фермента рибонуклеазы.
 § В случае белковых молекул, состоящих более чем из одного полипептида, происходит образование комплексной четвертичной структуры, в которой объединяются третичные структуры отдельных полипептидов. В качестве примера можно рассмотреть модель молекулы гемоглобина человека, состоящей из двух £-цепочек и двух β-цепочек, которые формируют стабильную тетрамерную структуру с помощью водородных связей. Каждая из глобиновых цепочек содержит также молекулу гема, который в комплексе с железом способен связывать молекулы кислорода, обеспечивая их транспортировку эритроцитами крови.
§ В случае белковых молекул, состоящих более чем из одного полипептида, происходит образование комплексной четвертичной структуры, в которой объединяются третичные структуры отдельных полипептидов. В качестве примера можно рассмотреть модель молекулы гемоглобина человека, состоящей из двух £-цепочек и двух β-цепочек, которые формируют стабильную тетрамерную структуру с помощью водородных связей. Каждая из глобиновых цепочек содержит также молекулу гема, который в комплексе с железом способен связывать молекулы кислорода, обеспечивая их транспортировку эритроцитами крови.
 Четвертичная структура молекулы гемоглобина человека.
Четвертичная структура молекулы гемоглобина человека.