

5974a95168b019f4c5905e4bc2af1b5a.ppt
- Количество слайдов: 117
 ENZİMLER
ENZİMLER
 GİRİŞ VE TANIM İlk enzim çalışmaları sindirime ilişkin enzimlerin araştırılmaya başlandığı 17601825 yılları arasındadır. Enzimler biyolojik sistemlerde oluşan tepkimelerde etkili olan biyolojik katalizörler olarak tanımlanmaktadır. Termodinamik yönden oluşabilen bir tepkimenin hızını katalizörler, tepkimenin denge sabitini değiştirmeden 1011 kat veya daha fazla arttırmaktadır.
GİRİŞ VE TANIM İlk enzim çalışmaları sindirime ilişkin enzimlerin araştırılmaya başlandığı 17601825 yılları arasındadır. Enzimler biyolojik sistemlerde oluşan tepkimelerde etkili olan biyolojik katalizörler olarak tanımlanmaktadır. Termodinamik yönden oluşabilen bir tepkimenin hızını katalizörler, tepkimenin denge sabitini değiştirmeden 1011 kat veya daha fazla arttırmaktadır.
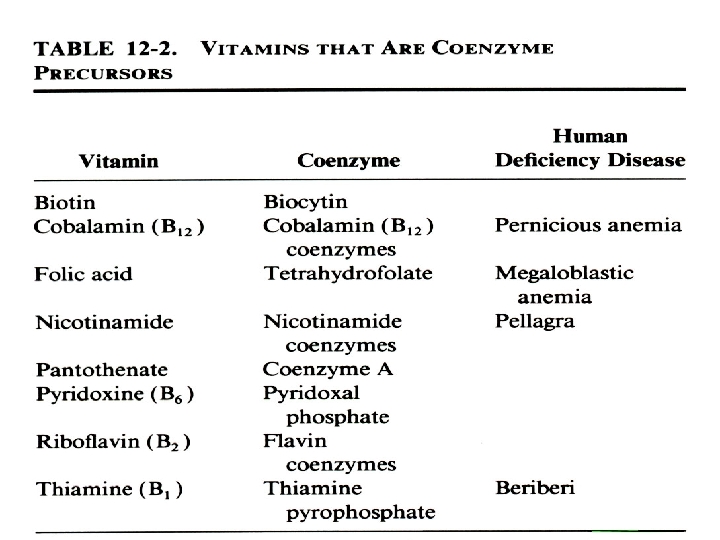
 Kofaktörler: Bazı enzimler, enzimatik reaksiyon için gerekli olan bir non protein faktörle birleşir. Bu faktörler arasında metal iyonlar (Zn+2, Fe+2) ve koenzim olarak bilinen, genellikle vitamin türevi olan (NAD+, FAD, Co. A) organik moleküller yer alır. Holoenzim, kofaktörü ile birlikte enzimi ifade eder. Apoenzim holoenzimin protein kısmını ifade eder. KOENZİM APOENZİM HOLOENZİM
Kofaktörler: Bazı enzimler, enzimatik reaksiyon için gerekli olan bir non protein faktörle birleşir. Bu faktörler arasında metal iyonlar (Zn+2, Fe+2) ve koenzim olarak bilinen, genellikle vitamin türevi olan (NAD+, FAD, Co. A) organik moleküller yer alır. Holoenzim, kofaktörü ile birlikte enzimi ifade eder. Apoenzim holoenzimin protein kısmını ifade eder. KOENZİM APOENZİM HOLOENZİM
 Apoenzim Koenzim Holoenzim Kofaktör, ya Fe 2+, Mg 2+, Mn 2+, Zn 2+ gibi bir veya daha fazla inorganik iyon ya da koenzim denen kompleks bir moleküldür
Apoenzim Koenzim Holoenzim Kofaktör, ya Fe 2+, Mg 2+, Mn 2+, Zn 2+ gibi bir veya daha fazla inorganik iyon ya da koenzim denen kompleks bir moleküldür
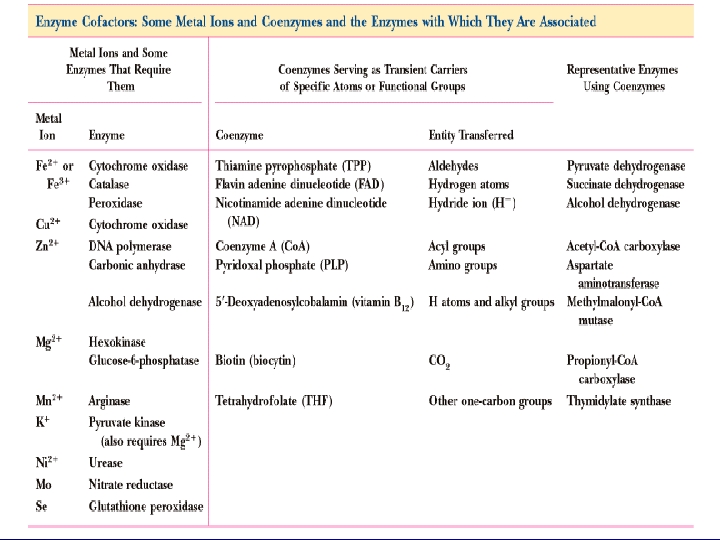
, zimojeni aktifleştiren maddeye de kinaz adı") Holoenzim= Apoenzim +Koenzim Enzimin inaktif şekline zimojen (proenzim), zimojeni aktifleştiren maddeye de kinaz adı verilir. Birden fazla şekilde bulunan enzimlere izoenzim denir. Prostetik grup enzimden ayrılamayan sıkıca bağlı bir koenzimdir. (Ör: Karboksilazların biyotini) Bir reaksiyonu hızlandıran fakat kendisi reaksiyondan değişmeksizin çıkan maddeye ‘‘KATALİZÖR’’ denir. Enzimlerle reaksiyon veren maddelere ‘‘SUBSTRAT’’ denir.
Holoenzim= Apoenzim +Koenzim Enzimin inaktif şekline zimojen (proenzim), zimojeni aktifleştiren maddeye de kinaz adı verilir. Birden fazla şekilde bulunan enzimlere izoenzim denir. Prostetik grup enzimden ayrılamayan sıkıca bağlı bir koenzimdir. (Ör: Karboksilazların biyotini) Bir reaksiyonu hızlandıran fakat kendisi reaksiyondan değişmeksizin çıkan maddeye ‘‘KATALİZÖR’’ denir. Enzimlerle reaksiyon veren maddelere ‘‘SUBSTRAT’’ denir.
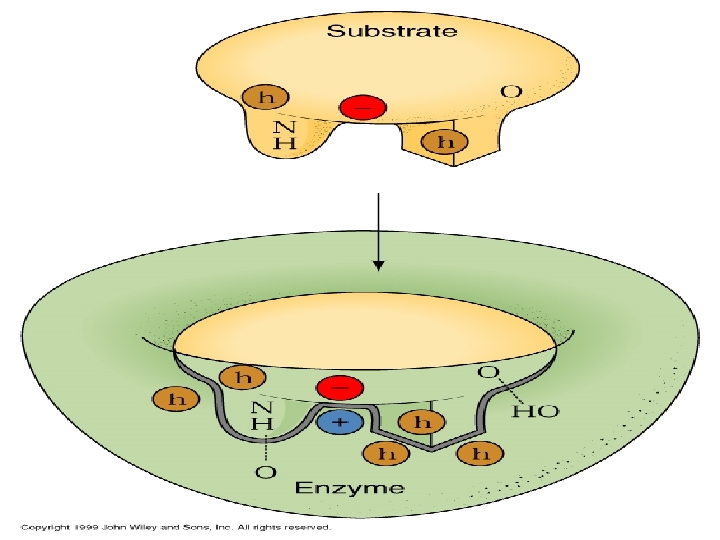
 Enzim-Substrat SUBSTRAT ENZİM SUBSTRAT KOMPLEKSİ
Enzim-Substrat SUBSTRAT ENZİM SUBSTRAT KOMPLEKSİ
 Substrat Değişken durum X Ürün Enzim substrata bağlanırsa reaksiyon daha ileri gitmez. . Enzim hem substratı seçer hem de değişken durumun oluşumunu teşvik eder.
Substrat Değişken durum X Ürün Enzim substrata bağlanırsa reaksiyon daha ileri gitmez. . Enzim hem substratı seçer hem de değişken durumun oluşumunu teşvik eder.
 ENZİMLERİN ÖZELLİKLERİ Enzimler, protein tabiatındadırlar. Hücre içinde ve dışında bulunurlar. Biyolojik maddeleri katalizlerler. Enerji açığa çıkaran maddelerdir. Üzerinde aktif merkez ile bağlanma yeri vardır. Kinetik olarak çalışırlar. Enzimler spesifiktir.
ENZİMLERİN ÖZELLİKLERİ Enzimler, protein tabiatındadırlar. Hücre içinde ve dışında bulunurlar. Biyolojik maddeleri katalizlerler. Enerji açığa çıkaran maddelerdir. Üzerinde aktif merkez ile bağlanma yeri vardır. Kinetik olarak çalışırlar. Enzimler spesifiktir.
’nın ‘‘Nomenculture Committe’’ tarafından tavsiye edilen sınıflandırma şöyledir;") ENZİMLERİN ADLANDIRILMASI International Union of Biochemistry (IUB)’nın ‘‘Nomenculture Committe’’ tarafından tavsiye edilen sınıflandırma şöyledir; (1984) Enzim sınıflandırma ve isimlendirilmesinde iki yol izlenir; 1 -Sistematik Yol 2 -Çalışma veya trivial yoldur. Sistematik yol, enzimin muhtemel fonksiyonu aynı olacak şekilde isimlendirilmesidir. Enzim katalitik olarak tanımlanır. Trivial yol, kısa ve genel kullanımdan sıkça bahsedilen enzimlerin sistematikte yer almayan şekli verilir.
ENZİMLERİN ADLANDIRILMASI International Union of Biochemistry (IUB)’nın ‘‘Nomenculture Committe’’ tarafından tavsiye edilen sınıflandırma şöyledir; (1984) Enzim sınıflandırma ve isimlendirilmesinde iki yol izlenir; 1 -Sistematik Yol 2 -Çalışma veya trivial yoldur. Sistematik yol, enzimin muhtemel fonksiyonu aynı olacak şekilde isimlendirilmesidir. Enzim katalitik olarak tanımlanır. Trivial yol, kısa ve genel kullanımdan sıkça bahsedilen enzimlerin sistematikte yer almayan şekli verilir.
 Geleneksel Adlandırma: Enzimler etkili oldukları substratın sonuna –az eki getirilerek (ÜREAZ, AMİLAZ, ARJİNAZ,") A-) Geleneksel Adlandırma: Enzimler etkili oldukları substratın sonuna –az eki getirilerek (ÜREAZ, AMİLAZ, ARJİNAZ, PROTEAZ, LİPAZ) veya katalizledikleri tepkimeyi tanımlayan (Laktat Dehidrogenaz, Adenilat Siklaz) isimleri kullanılarak adlandırılmıştır.
A-) Geleneksel Adlandırma: Enzimler etkili oldukları substratın sonuna –az eki getirilerek (ÜREAZ, AMİLAZ, ARJİNAZ, PROTEAZ, LİPAZ) veya katalizledikleri tepkimeyi tanımlayan (Laktat Dehidrogenaz, Adenilat Siklaz) isimleri kullanılarak adlandırılmıştır.
 Sistematik Adlandırma: Uluslar arası biyokimya ve moleküler biyoloji birliğinin (IUBMB) göre; enzim adlandırmaları,") B-) Sistematik Adlandırma: Uluslar arası biyokimya ve moleküler biyoloji birliğinin (IUBMB) göre; enzim adlandırmaları, enzimin etkilediği tepkimenin türüne ve mekanizmasına göre yapılmaktadır. Tepkimeler ve bu tepkimeleri katalize eden enzimler 6 ana gruba ayrılmıştır. Bu 6 grubun 4 -13 arasında değişen alt sınıfları bulunmaktadır. Her enzime 4 sayı ile belirlenen bir kod numarası verilmiştir.
B-) Sistematik Adlandırma: Uluslar arası biyokimya ve moleküler biyoloji birliğinin (IUBMB) göre; enzim adlandırmaları, enzimin etkilediği tepkimenin türüne ve mekanizmasına göre yapılmaktadır. Tepkimeler ve bu tepkimeleri katalize eden enzimler 6 ana gruba ayrılmıştır. Bu 6 grubun 4 -13 arasında değişen alt sınıfları bulunmaktadır. Her enzime 4 sayı ile belirlenen bir kod numarası verilmiştir.
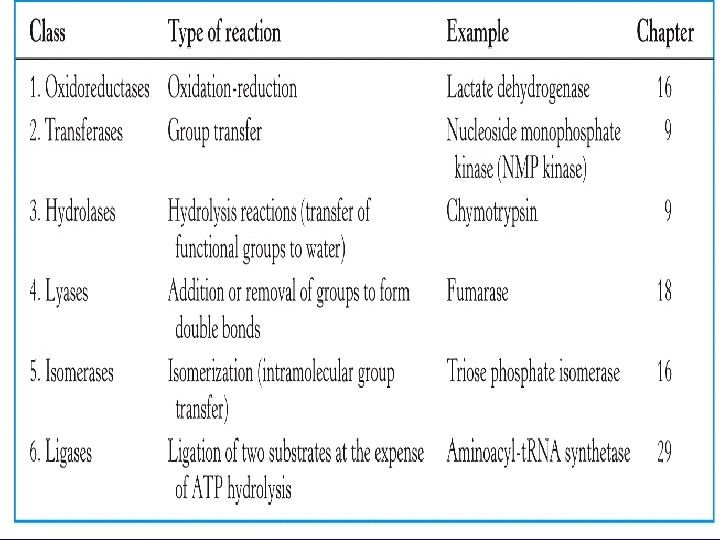
 Oksidoredüktazlar 2 -) Transferazlar 3 -) Hidrolazlar 4 -) Liyazlar") ENZİMLERİN SINIFLANDIRILMASI 1 -) Oksidoredüktazlar 2 -) Transferazlar 3 -) Hidrolazlar 4 -) Liyazlar OTH ALİL 5 -) İzomerazlar 6 -) Ligazlar v NOT: Sınıflandırmada baş harflerini birleştirirsek ‘‘OTHALİL’’ elde ederiz böylelikle enzimlerin sırasını karıştırmamış oluruz.
ENZİMLERİN SINIFLANDIRILMASI 1 -) Oksidoredüktazlar 2 -) Transferazlar 3 -) Hidrolazlar 4 -) Liyazlar OTH ALİL 5 -) İzomerazlar 6 -) Ligazlar v NOT: Sınıflandırmada baş harflerini birleştirirsek ‘‘OTHALİL’’ elde ederiz böylelikle enzimlerin sırasını karıştırmamış oluruz.
 İSİMLENDİRME VE NUMARALANDIRMA 1961 yılında enzim komisyonu, kod numaralı sistemi geliştirdi. 4 element bazı farklarla ayrılarak işlem yapıldı. A-)Birinci numara altı ana bölümden birisini simgeler. B-)İkinci numara subklası gösterir. C-)Üçüncü numara subsubklası ifade eder. D-)Dördüncü numara ise subsubklastaki enzimin seri numarasını gösterir.
İSİMLENDİRME VE NUMARALANDIRMA 1961 yılında enzim komisyonu, kod numaralı sistemi geliştirdi. 4 element bazı farklarla ayrılarak işlem yapıldı. A-)Birinci numara altı ana bölümden birisini simgeler. B-)İkinci numara subklası gösterir. C-)Üçüncü numara subsubklası ifade eder. D-)Dördüncü numara ise subsubklastaki enzimin seri numarasını gösterir.
 Örnek verecek olursak; 1. 1 abcd a: Ana sınıf b: Subklas c: Subsubklas d: Subsubklas seri no
Örnek verecek olursak; 1. 1 abcd a: Ana sınıf b: Subklas c: Subsubklas d: Subsubklas seri no
 Tepkimenin tipini birinci sayı, vericinin etkilediği grubu ikinci sayı, alıcı olarak yararlanılan grubu üçüncü sayı ve adlandırılan enzimi dördüncü sayı belirlemektedir. Örnek : EC(Enzim komisyonu). 2. 7. 1. 1 2 Transferaz türü tepkimeyi 7 Fosfat transferini 1 Fosfat alıcısının bir alkol olduğunu gösterir, 1 ATP molekülünden glikozun altıncı karbonundaki hidroksil grubuna fosfat transferi yapan enzimi, ATP-DHeksozaltıfosfotransferazı (hekzokinaz) tanımlamaktadır.
Tepkimenin tipini birinci sayı, vericinin etkilediği grubu ikinci sayı, alıcı olarak yararlanılan grubu üçüncü sayı ve adlandırılan enzimi dördüncü sayı belirlemektedir. Örnek : EC(Enzim komisyonu). 2. 7. 1. 1 2 Transferaz türü tepkimeyi 7 Fosfat transferini 1 Fosfat alıcısının bir alkol olduğunu gösterir, 1 ATP molekülünden glikozun altıncı karbonundaki hidroksil grubuna fosfat transferi yapan enzimi, ATP-DHeksozaltıfosfotransferazı (hekzokinaz) tanımlamaktadır.
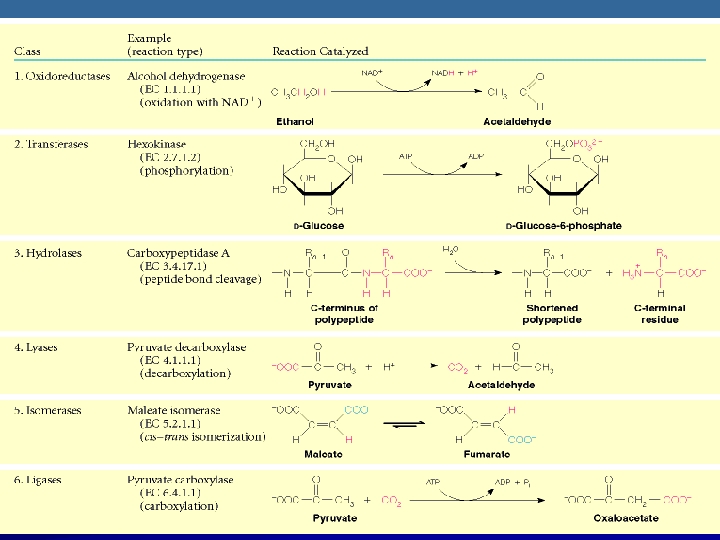
 OKSİDOREDÜKTAZLAR: ( Oxidoreductases) Bu sınıftaki enzimler oksido- redüksiyon reaksiyonlarını katalizlerler. R-CH 2") 1 -) OKSİDOREDÜKTAZLAR: ( Oxidoreductases) Bu sınıftaki enzimler oksido- redüksiyon reaksiyonlarını katalizlerler. R-CH 2 OH+NAD+ (Primer Alkol) R-CHO+NADH+H+ (Aldehid) Enzim : EC. 1. 1, Alkol NAD+ oksidoredüktaz
1 -) OKSİDOREDÜKTAZLAR: ( Oxidoreductases) Bu sınıftaki enzimler oksido- redüksiyon reaksiyonlarını katalizlerler. R-CH 2 OH+NAD+ (Primer Alkol) R-CHO+NADH+H+ (Aldehid) Enzim : EC. 1. 1, Alkol NAD+ oksidoredüktaz
 DEHİDROJENAZLAR: (Redüktazlar) Uygun bir H+ alıcısının mevcudiyeti halinde substrattan hidrojeni ayırırlar. 1.") 1. a-) DEHİDROJENAZLAR: (Redüktazlar) Uygun bir H+ alıcısının mevcudiyeti halinde substrattan hidrojeni ayırırlar. 1. b-) OKSİDAZLAR: H+ alıcısı olarak oksijene sahiptirler. Aldehit oksidaz gibi. Aldehit + H 2 O +O 2 Asit + H 2 O 2
1. a-) DEHİDROJENAZLAR: (Redüktazlar) Uygun bir H+ alıcısının mevcudiyeti halinde substrattan hidrojeni ayırırlar. 1. b-) OKSİDAZLAR: H+ alıcısı olarak oksijene sahiptirler. Aldehit oksidaz gibi. Aldehit + H 2 O +O 2 Asit + H 2 O 2
 1: Oksidoredüktaz Oksidaz Dehidrogenaz - -
1: Oksidoredüktaz Oksidaz Dehidrogenaz - -
 1: Oxidoredüktaz – Oksijenaz
1: Oxidoredüktaz – Oksijenaz
 TRANSFERAZLAR: Bu gruptaki enzimler bir fonksiyonel grubu bir molekülden diğerine aktarmaktadırlar. CH") 2 -) TRANSFERAZLAR: Bu gruptaki enzimler bir fonksiyonel grubu bir molekülden diğerine aktarmaktadırlar. CH 3 CO-Co. A + HOCH 2 N+(CH 3)3 Asetil Co. A Kolin CH 3 CO-O-CH 2 N+(CH 3)3 + Co. A (O-Astilkolin) Enzim. EC. 2. 3. 1. 6, Asetil Co. A: Kolin-O-asetil transferaz
2 -) TRANSFERAZLAR: Bu gruptaki enzimler bir fonksiyonel grubu bir molekülden diğerine aktarmaktadırlar. CH 3 CO-Co. A + HOCH 2 N+(CH 3)3 Asetil Co. A Kolin CH 3 CO-O-CH 2 N+(CH 3)3 + Co. A (O-Astilkolin) Enzim. EC. 2. 3. 1. 6, Asetil Co. A: Kolin-O-asetil transferaz
 2: Transferaz - Kinaz
2: Transferaz - Kinaz
 1 C’lu grupları transfer ederler(Metil transferaz) 2. b-) Aldehitten keton transfer ederler") 2. a-) 1 C’lu grupları transfer ederler(Metil transferaz) 2. b-) Aldehitten keton transfer ederler (Transketolaz) 2. c-) Diğerleri, Alkil, N, P, S’lü grupları transfer ederler. 3 -)HİDROLAZLAR: Bu enzimler, su katılması ile bağların parçalandığı hidroliz tepkimelerini katalizlemektedir. H 2 N-CO-NH 2+H 2 O 2 NH 3+CO 2
2. a-) 1 C’lu grupları transfer ederler(Metil transferaz) 2. b-) Aldehitten keton transfer ederler (Transketolaz) 2. c-) Diğerleri, Alkil, N, P, S’lü grupları transfer ederler. 3 -)HİDROLAZLAR: Bu enzimler, su katılması ile bağların parçalandığı hidroliz tepkimelerini katalizlemektedir. H 2 N-CO-NH 2+H 2 O 2 NH 3+CO 2
 Basit esterazlar 3. g-) Selülaz") Enzim. EC. 3. 5. 1. 5, Üreaz 3. a-) Basit esterazlar 3. g-) Selülaz 3. b-) Lipazlar 3. h-) İnulaz 3. c-) Fosfatazlar 3. i-) Glikozidaz 3. d-) Kolinesterazlar 3. j-) Üreaz 3. e-) Peptid hidrolazlar 3. k-) Asparajinaz 3. f-) Nükleazlar 3. l-) Arjinaz
Enzim. EC. 3. 5. 1. 5, Üreaz 3. a-) Basit esterazlar 3. g-) Selülaz 3. b-) Lipazlar 3. h-) İnulaz 3. c-) Fosfatazlar 3. i-) Glikozidaz 3. d-) Kolinesterazlar 3. j-) Üreaz 3. e-) Peptid hidrolazlar 3. k-) Asparajinaz 3. f-) Nükleazlar 3. l-) Arjinaz
 LİYAZLAR: Bu gruptaki enzimler hidrolizden başka mekanizmalarla C-C, C-S ve bazı C-N") 4 -) LİYAZLAR: Bu gruptaki enzimler hidrolizden başka mekanizmalarla C-C, C-S ve bazı C-N bağlarının parçalanması tepkimelerinde görev yapmaktadırlar. CH 3 -CO-COO CH 3 -CH 0+CO 2 (PİRUVAT) (ASETALDEHİT) Enzim. EC. 4. 1. 1. 1, Piruvat dekarboksilaz 4. a-)Dekarboksilazlar. Okzaloasetat karboksilaz gibi. 4. b-)Karbonik Anhidraz 4. c-)Aspartat amonyak liyaz 4. d-)Sistein desülfhidraz
4 -) LİYAZLAR: Bu gruptaki enzimler hidrolizden başka mekanizmalarla C-C, C-S ve bazı C-N bağlarının parçalanması tepkimelerinde görev yapmaktadırlar. CH 3 -CO-COO CH 3 -CH 0+CO 2 (PİRUVAT) (ASETALDEHİT) Enzim. EC. 4. 1. 1. 1, Piruvat dekarboksilaz 4. a-)Dekarboksilazlar. Okzaloasetat karboksilaz gibi. 4. b-)Karbonik Anhidraz 4. c-)Aspartat amonyak liyaz 4. d-)Sistein desülfhidraz
 4: Liyaz - Fumaraz
4: Liyaz - Fumaraz
İZOMERAZLAR: Optik veya geometrik izomerlerin rasemizasyonu tepkimelerini katalize eden enzim grubudur. L-Alanin D-Alanin") 5 -)İZOMERAZLAR: Optik veya geometrik izomerlerin rasemizasyonu tepkimelerini katalize eden enzim grubudur. L-Alanin D-Alanin Enzim. EC. 5. 1. 1. 1, Alanin Rasemaz 5. a-) Rasemazlar ve epimerazlar 5. b-) Cis transizomerazlar 5. c-) İntramoleküler oksidoredüktazlar 5. d-) İntramoleküler transferazlar
5 -)İZOMERAZLAR: Optik veya geometrik izomerlerin rasemizasyonu tepkimelerini katalize eden enzim grubudur. L-Alanin D-Alanin Enzim. EC. 5. 1. 1. 1, Alanin Rasemaz 5. a-) Rasemazlar ve epimerazlar 5. b-) Cis transizomerazlar 5. c-) İntramoleküler oksidoredüktazlar 5. d-) İntramoleküler transferazlar
 5: Isomeraz Epimeraz ve Rasemaz Mutaz
5: Isomeraz Epimeraz ve Rasemaz Mutaz
LİGAZLAR: C ve O, S, N arasında yeni bağ oluşumunu katalize eden enzimlerdir.") 6 -)LİGAZLAR: C ve O, S, N arasında yeni bağ oluşumunu katalize eden enzimlerdir. Tepkimelerde gerekli enerji, yüksek enerjili bir fosfat bileşiğinin hidrolizi ile sağlanmaktadır. -OOC-CH CH -COO-+Co. A+GTP 2 2 (Süksinat) -OOC-CH CH -CO-Co. A+GDP+Pi 2 2 Co. A) (Süksinil
6 -)LİGAZLAR: C ve O, S, N arasında yeni bağ oluşumunu katalize eden enzimlerdir. Tepkimelerde gerekli enerji, yüksek enerjili bir fosfat bileşiğinin hidrolizi ile sağlanmaktadır. -OOC-CH CH -COO-+Co. A+GTP 2 2 (Süksinat) -OOC-CH CH -CO-Co. A+GDP+Pi 2 2 Co. A) (Süksinil
 6: Ligaz - Piruvatkarboksilaz
6: Ligaz - Piruvatkarboksilaz
 Aktif Bölgeler: Enzim moleküllerinde aktif bölge denilen özel bir cep ya") ENZİMLERİN ÖZELLİKLERİ A-) Aktif Bölgeler: Enzim moleküllerinde aktif bölge denilen özel bir cep ya da yuva bulunur. Aktif bölge, substrata komplementer olan üç boyutlu bir yüzey yaratan amino asit yan zincirleri içerir. Aktif bölge, substratı bağlayarak ES kompleksi meydana getirir. ES, sonradan enzim ve ürüne parçalanan EP’ye dönüşür. Aktif merkez için E-S bağlanmasını açıklayan iki model öne sürülmektedir.
ENZİMLERİN ÖZELLİKLERİ A-) Aktif Bölgeler: Enzim moleküllerinde aktif bölge denilen özel bir cep ya da yuva bulunur. Aktif bölge, substrata komplementer olan üç boyutlu bir yüzey yaratan amino asit yan zincirleri içerir. Aktif bölge, substratı bağlayarak ES kompleksi meydana getirir. ES, sonradan enzim ve ürüne parçalanan EP’ye dönüşür. Aktif merkez için E-S bağlanmasını açıklayan iki model öne sürülmektedir.
 Enzimle katalizlenen bir reaksiyonun ayırt edici özelliği, enzim üzerinde aktif merkez denen bir cep sınırları içinde meydana gelmesidir.
Enzimle katalizlenen bir reaksiyonun ayırt edici özelliği, enzim üzerinde aktif merkez denen bir cep sınırları içinde meydana gelmesidir.
 Anahtar-Kilit Modeli(Fischer Modeli): Bu modelde, substrat ve enzimin aktif yerinin birbirine uyacak şekilde") a-) Anahtar-Kilit Modeli(Fischer Modeli): Bu modelde, substrat ve enzimin aktif yerinin birbirine uyacak şekilde önceden belirlenmiş olduğu varsayılmaktadır. S Aktif Bölge ES Kompleksi ES E
a-) Anahtar-Kilit Modeli(Fischer Modeli): Bu modelde, substrat ve enzimin aktif yerinin birbirine uyacak şekilde önceden belirlenmiş olduğu varsayılmaktadır. S Aktif Bölge ES Kompleksi ES E
 Uyum oluşturma modeli / El eldiven modeli (Koshland Modeli): Ø Bu modelde aktif") b-) Uyum oluşturma modeli / El eldiven modeli (Koshland Modeli): Ø Bu modelde aktif merkez esnek yapıdadır. Proteinin tersiyer yapısında oluşan bir değişiklik ile enzim, substratını katalize uygun ve en doğru biçimde bağlayacak şekilde biçimsel bir değişikliğe uğramaktadır.
b-) Uyum oluşturma modeli / El eldiven modeli (Koshland Modeli): Ø Bu modelde aktif merkez esnek yapıdadır. Proteinin tersiyer yapısında oluşan bir değişiklik ile enzim, substratını katalize uygun ve en doğru biçimde bağlayacak şekilde biçimsel bir değişikliğe uğramaktadır.
 Substrat ES Kompleksi Enzim
Substrat ES Kompleksi Enzim
 Katalitik Etkinlik: Enzimle katalizlenen reaksiyonların çoğu katalizlenmeyen reaksiyonlara göre 10 -3 ile 108") B-) Katalitik Etkinlik: Enzimle katalizlenen reaksiyonların çoğu katalizlenmeyen reaksiyonlara göre 10 -3 ile 108 kere daha hızlı olarak oldukça etkindir. Her enzim molekülü saniyede 100 -1000 substrat molekülünü ürüne çevirme yeteneğine sahiptir. C-)Spesifiklik: Enzimler bir veya birkaç substratla etkileşerek ve sadece tek tip kimyasal reaksiyonu katalizleyerek oldukça spesifiktir.
B-) Katalitik Etkinlik: Enzimle katalizlenen reaksiyonların çoğu katalizlenmeyen reaksiyonlara göre 10 -3 ile 108 kere daha hızlı olarak oldukça etkindir. Her enzim molekülü saniyede 100 -1000 substrat molekülünü ürüne çevirme yeteneğine sahiptir. C-)Spesifiklik: Enzimler bir veya birkaç substratla etkileşerek ve sadece tek tip kimyasal reaksiyonu katalizleyerek oldukça spesifiktir.
 Kofaktörler: Bazı enzimler, enzimatik reaksiyon için gerekli olan bir non protein faktörle birleşir.") D-) Kofaktörler: Bazı enzimler, enzimatik reaksiyon için gerekli olan bir non protein faktörle birleşir. Bu faktörler arasında metal iyonlar (Zn+2, Fe+2) ve koenzim olarak bilinen, genellikle vitamin türevi olan (NAD+, FAD, Co. A) organik moleküller yer alır. Holoenzim, kofaktörü ile birlikte enzimi ifade eder. Apoenzim holoenzimin protein kısmını ifade eder. Prostetik grup enzimden ayrılamayan sıkıca bağlı bir koenzimdir. (Ör: Karboksilazların biyotini)
D-) Kofaktörler: Bazı enzimler, enzimatik reaksiyon için gerekli olan bir non protein faktörle birleşir. Bu faktörler arasında metal iyonlar (Zn+2, Fe+2) ve koenzim olarak bilinen, genellikle vitamin türevi olan (NAD+, FAD, Co. A) organik moleküller yer alır. Holoenzim, kofaktörü ile birlikte enzimi ifade eder. Apoenzim holoenzimin protein kısmını ifade eder. Prostetik grup enzimden ayrılamayan sıkıca bağlı bir koenzimdir. (Ör: Karboksilazların biyotini)
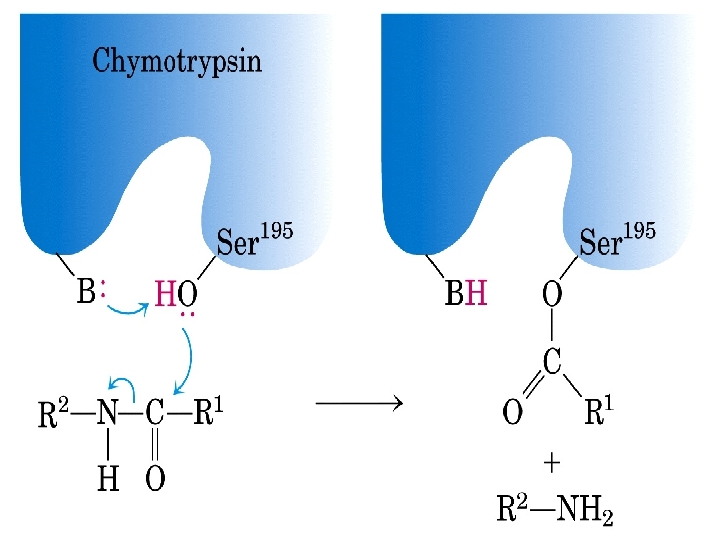
 Chymotrypsin
Chymotrypsin
 Chymotrypsin
Chymotrypsin
 Hekzokinaz
Hekzokinaz
 Serinproteaz
Serinproteaz
 Allosterik Enzim wenig aktiv Aktiver Enzym. Substratkomplex
Allosterik Enzim wenig aktiv Aktiver Enzym. Substratkomplex
 Models Allosteric site") Mechanism and Example of Allosteric Effect Kinetics R = Relax (active) Models Allosteric site vo (+) Allosteric site [S] vo X T = Tense (inactive) Cooperation A (+) Homotropic (+) Concerted Heterotropic (+) Sequential (+) [S] I vo (-) [S] X (-) X Heterotropic (-) Concerted Juang RH (2004) BCbasics
Mechanism and Example of Allosteric Effect Kinetics R = Relax (active) Models Allosteric site vo (+) Allosteric site [S] vo X T = Tense (inactive) Cooperation A (+) Homotropic (+) Concerted Heterotropic (+) Sequential (+) [S] I vo (-) [S] X (-) X Heterotropic (-) Concerted Juang RH (2004) BCbasics
 ENZİM KİNETİĞİ Enzim kinetiği, enzimlerin katalize ettikleri reaksiyonların hızlarını inceler. Isı yükseldikçe enzim aktivitesi artmaktadır. Ateşli hastalıklarda metabolizma hızlanmaktadır.
ENZİM KİNETİĞİ Enzim kinetiği, enzimlerin katalize ettikleri reaksiyonların hızlarını inceler. Isı yükseldikçe enzim aktivitesi artmaktadır. Ateşli hastalıklarda metabolizma hızlanmaktadır.
 S E k 1 k-1 S E k 2 P -Km, Vmax 0. 5 Vmax Km Eric Niederhoffer SIU-SOM
S E k 1 k-1 S E k 2 P -Km, Vmax 0. 5 Vmax Km Eric Niederhoffer SIU-SOM
 Enerji Değişimi EST S ES EP Reaksiyon Yönü T = Transition state P Enerji Düşüşü Enerji İhtiyacı ST
Enerji Değişimi EST S ES EP Reaksiyon Yönü T = Transition state P Enerji Düşüşü Enerji İhtiyacı ST
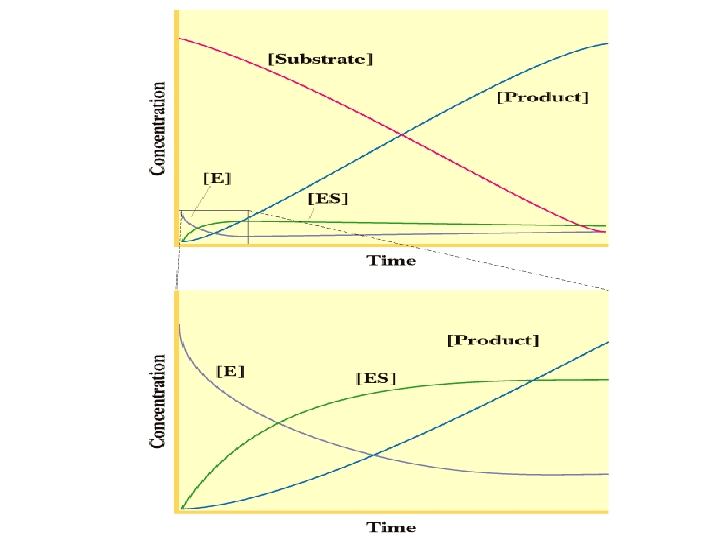
 Enzim Kinetiği Steady State Teorisi E + S E +P In steady state, the production and consumption of the transition state proceed at the same rate. So the concentration of transition state keeps a constant.
Enzim Kinetiği Steady State Teorisi E + S E +P In steady state, the production and consumption of the transition state proceed at the same rate. So the concentration of transition state keeps a constant.
 ENZİM REAKSİYONUNUN HIZINA ETKİ EDEN FAKTÖRLER Uygulamada bir enzimin etkinliği, verilen bir zamanda ve belli başlangıç koşullarında, enzimin bilinen miktarının etkisi altında değişime uğrayan substratın miktarı ile ölçülür. Bir enzimin etkinliğinin, özellikle şu faktörlerle değiştiği görülür; 1 - SICAKLIK 2 - ZAMAN 3 - p. H 4 - ENZİM KONSANTRASYONU 5 - SUBSTRAT KONSANTRASYONU 6 - DİĞER FAKTÖRLER
ENZİM REAKSİYONUNUN HIZINA ETKİ EDEN FAKTÖRLER Uygulamada bir enzimin etkinliği, verilen bir zamanda ve belli başlangıç koşullarında, enzimin bilinen miktarının etkisi altında değişime uğrayan substratın miktarı ile ölçülür. Bir enzimin etkinliğinin, özellikle şu faktörlerle değiştiği görülür; 1 - SICAKLIK 2 - ZAMAN 3 - p. H 4 - ENZİM KONSANTRASYONU 5 - SUBSTRAT KONSANTRASYONU 6 - DİĞER FAKTÖRLER
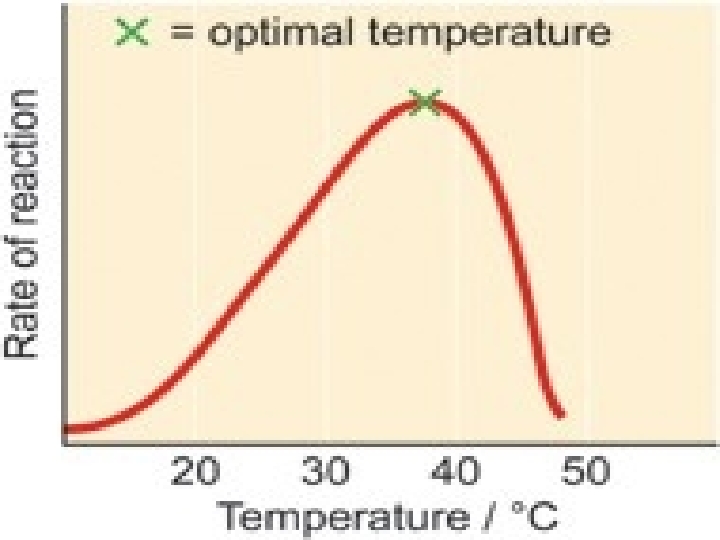
 SICAKLIK: Enzimatik tepkimelerde sıcaklığın artması ile moleküller arası, çarpışmanın artışına bağlı olarak") 1 -) SICAKLIK: Enzimatik tepkimelerde sıcaklığın artması ile moleküller arası, çarpışmanın artışına bağlı olarak genellikle tepkimenin hızı artmaktadır. Ancak maksimum bir hıza ulaşıldıktan sonra sıcaklığın artışı ile hızda tekrar bir azalma gözlenmektedir. Maksimum hıza ulaşıldığındaki sıcaklığa optimal sıcaklık adı verilmektedir. Enzimler protein yapısında oldukları için bu sıcaklık üzerinde yapıyı oluşturan bağlar parçalanmakta ve molekül denatürasyona uğramaktadır. ES oluşumu bozulduğu için tepkimenin hızı azalmaktadır. Optimum sıcaklık insanlar için 37 o C, bazı mikroorganizmalar için daha yüksek olabilmektedir.
1 -) SICAKLIK: Enzimatik tepkimelerde sıcaklığın artması ile moleküller arası, çarpışmanın artışına bağlı olarak genellikle tepkimenin hızı artmaktadır. Ancak maksimum bir hıza ulaşıldıktan sonra sıcaklığın artışı ile hızda tekrar bir azalma gözlenmektedir. Maksimum hıza ulaşıldığındaki sıcaklığa optimal sıcaklık adı verilmektedir. Enzimler protein yapısında oldukları için bu sıcaklık üzerinde yapıyı oluşturan bağlar parçalanmakta ve molekül denatürasyona uğramaktadır. ES oluşumu bozulduğu için tepkimenin hızı azalmaktadır. Optimum sıcaklık insanlar için 37 o C, bazı mikroorganizmalar için daha yüksek olabilmektedir.
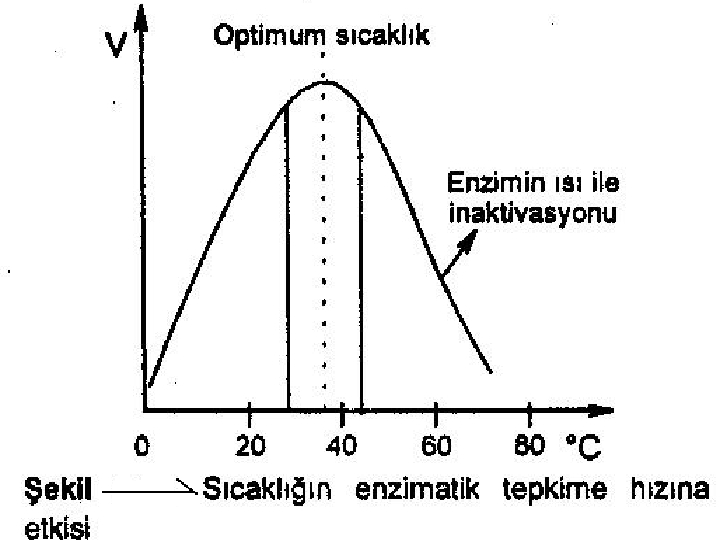
 ZAMAN: Enzim reaksiyonun hızı zamanla artar, çünkü reaksiyon ürünleri zamanla denatüre olur,") 2 -) ZAMAN: Enzim reaksiyonun hızı zamanla artar, çünkü reaksiyon ürünleri zamanla denatüre olur, substrat azalır veya tükenebilir. Bunun için enzim ölçümleri genellikle yaklaşık %10 nun kullanıldığı başlangıç zamanına rastlar. Hız Substrat konstrasyonun azaldığı yer 0 10 20 30 40 (Zaman)
2 -) ZAMAN: Enzim reaksiyonun hızı zamanla artar, çünkü reaksiyon ürünleri zamanla denatüre olur, substrat azalır veya tükenebilir. Bunun için enzim ölçümleri genellikle yaklaşık %10 nun kullanıldığı başlangıç zamanına rastlar. Hız Substrat konstrasyonun azaldığı yer 0 10 20 30 40 (Zaman)
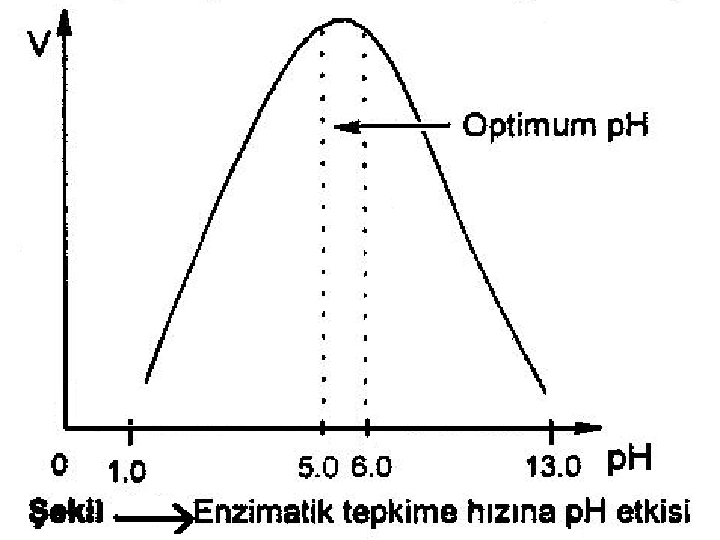
 p. H: enzimlerin aktiviteleri, ortamın hidrojen iyon konsantrasyonuna bağlı olarak değişmektedir. Enzimatik") 3 -) p. H: enzimlerin aktiviteleri, ortamın hidrojen iyon konsantrasyonuna bağlı olarak değişmektedir. Enzimatik tepkimenin hızının optimal olduğu ve her enzim için değişik olan bir optimum p. H değeri bulunmaktadır. Sıcaklık artışıyla enzimatik reaksiyonun hızındaki artış, sıcaklık belli bir değere yükselinceye kadar devam eder; daha yüksek sıcaklıklarda enzim denatüre olarak aktivitesini kaybeder ve reaksiyon hızı azalır
3 -) p. H: enzimlerin aktiviteleri, ortamın hidrojen iyon konsantrasyonuna bağlı olarak değişmektedir. Enzimatik tepkimenin hızının optimal olduğu ve her enzim için değişik olan bir optimum p. H değeri bulunmaktadır. Sıcaklık artışıyla enzimatik reaksiyonun hızındaki artış, sıcaklık belli bir değere yükselinceye kadar devam eder; daha yüksek sıcaklıklarda enzim denatüre olarak aktivitesini kaybeder ve reaksiyon hızı azalır

 Rate of an enzymatic reaction as a function of temperature and p. H
Rate of an enzymatic reaction as a function of temperature and p. H
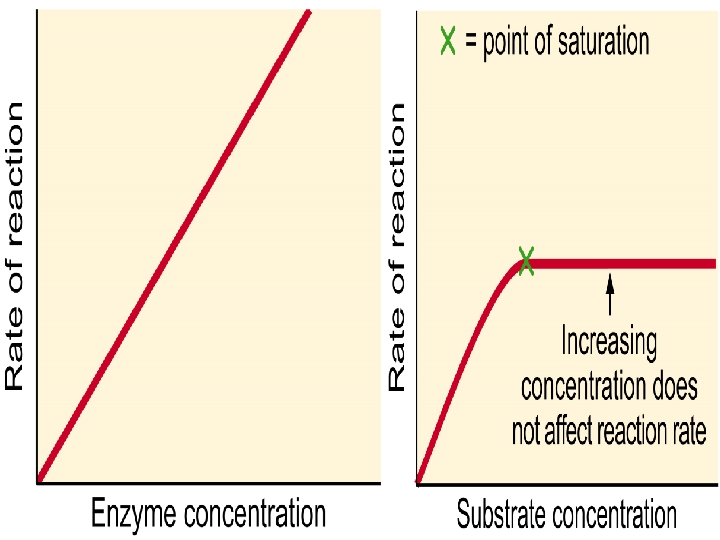
 Belirli bir p. H alanında enzimin etkinliği en fazladır. Bu p. H’ya o enzimin optimum Ph’sı denir. Örneğin: p. H: 1 -2 de en kuvvetli etki gösteren Pepsin, nötral veya alkali ortamda etkin değildir. 4 -) ENZİM KONSANTRASYONU: Enzimatik bir tepkime ortamında fazla miktarda substrat bulunması halinde tepkimenin hızı, enzimin konsantrasyonu ile orantılı olarak artmaktadır. Denge halinde sağa doğru olan hızı (k 1), sola doğru olan hızına(k 2) eşittir
Belirli bir p. H alanında enzimin etkinliği en fazladır. Bu p. H’ya o enzimin optimum Ph’sı denir. Örneğin: p. H: 1 -2 de en kuvvetli etki gösteren Pepsin, nötral veya alkali ortamda etkin değildir. 4 -) ENZİM KONSANTRASYONU: Enzimatik bir tepkime ortamında fazla miktarda substrat bulunması halinde tepkimenin hızı, enzimin konsantrasyonu ile orantılı olarak artmaktadır. Denge halinde sağa doğru olan hızı (k 1), sola doğru olan hızına(k 2) eşittir

![V 1=k 1. [E]. [S] V 2=k 2. [E]. [P] k 1. [E]. [S]=k](https://present5.com/presentation/5974a95168b019f4c5905e4bc2af1b5a/image-67.jpg "V 1=k 1. [E]. [S] V 2=k 2. [E]. [P] k 1. [E]. [S]=k") V 1=k 1. [E]. [S] V 2=k 2. [E]. [P] k 1. [E]. [S]=k 2. [E]. [P] V 1=V 2 Kdenge= k 1/k 2= [P]/ [S] Denge halinde enzim konsantrasyonun denge sabiti üzerine etkisi yoktur. Tepkime hızını etkileyen enzimler, hız ve denge sabitlerini değiştirmemektedir. Bir tepkimenin denge sabitini, ortamda bulunan enzim etkilememektedir.
V 1=k 1. [E]. [S] V 2=k 2. [E]. [P] k 1. [E]. [S]=k 2. [E]. [P] V 1=V 2 Kdenge= k 1/k 2= [P]/ [S] Denge halinde enzim konsantrasyonun denge sabiti üzerine etkisi yoktur. Tepkime hızını etkileyen enzimler, hız ve denge sabitlerini değiştirmemektedir. Bir tepkimenin denge sabitini, ortamda bulunan enzim etkilememektedir.
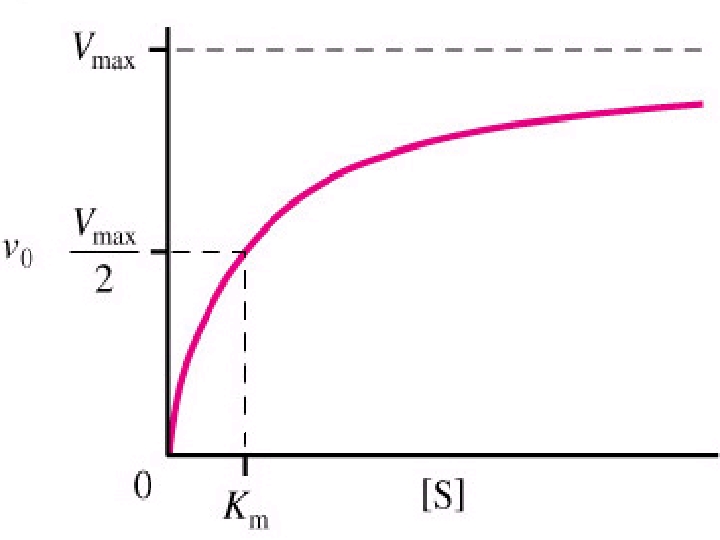
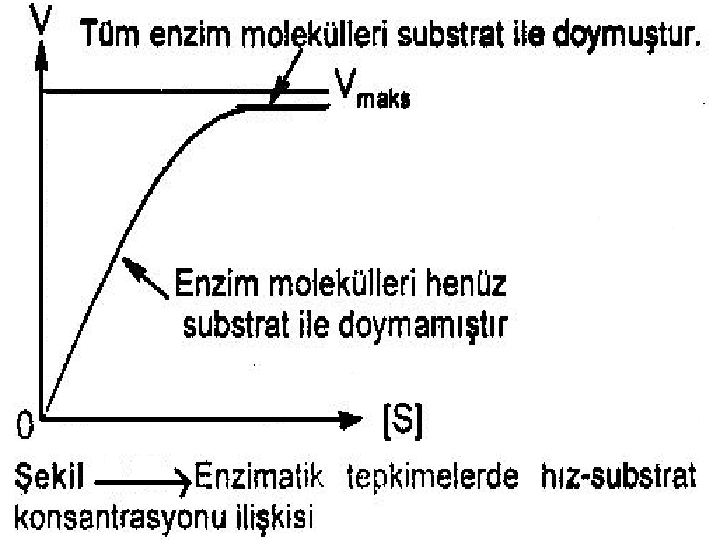
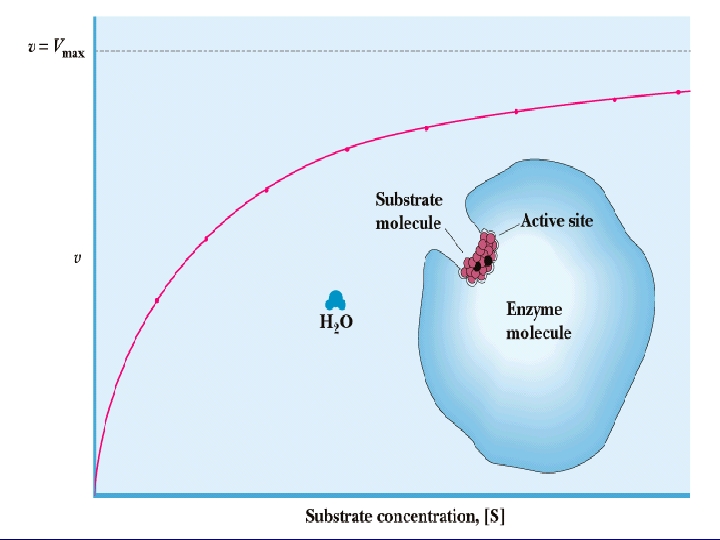
 SUBSTRAT KONSANTRASYONU: Sabit enzim konsantrasyonunda, enzim reaksiyonunun hızı belirli bir noktaya kadar") 5 -) SUBSTRAT KONSANTRASYONU: Sabit enzim konsantrasyonunda, enzim reaksiyonunun hızı belirli bir noktaya kadar substrat konsantrasyonu ile artar. Bundan sonra substrat konsantrasyonunun artması ile artık reaksiyon hızı değişmez. Başlangıçta artan [S], enzim molekülleri ile bağlanarak [ES] kompleksi oluşturmaktadır. Maksimum kapasiteye ulaşıldığında enzimde substrat bağlayacak tüm boş yerler dolduğundan, eklenen substrat artık [ES] kompleksi oluşturamayacağı için enzimatik tepkimenin hızı Vmax değişmemektedir.
5 -) SUBSTRAT KONSANTRASYONU: Sabit enzim konsantrasyonunda, enzim reaksiyonunun hızı belirli bir noktaya kadar substrat konsantrasyonu ile artar. Bundan sonra substrat konsantrasyonunun artması ile artık reaksiyon hızı değişmez. Başlangıçta artan [S], enzim molekülleri ile bağlanarak [ES] kompleksi oluşturmaktadır. Maksimum kapasiteye ulaşıldığında enzimde substrat bağlayacak tüm boş yerler dolduğundan, eklenen substrat artık [ES] kompleksi oluşturamayacağı için enzimatik tepkimenin hızı Vmax değişmemektedir.
 DİĞER FAKTÖRLER: Bunlar reaksiyon sonunda oluşan ürün, ortamdaki iyonların tabiatı, konsantrasyonları, allosterik") 6 -) DİĞER FAKTÖRLER: Bunlar reaksiyon sonunda oluşan ürün, ortamdaki iyonların tabiatı, konsantrasyonları, allosterik etki, ışık ve diğer fiziksel faktörler, hormonlar ve diğer biyokimyasal faktörler olarak söylenebilir.
6 -) DİĞER FAKTÖRLER: Bunlar reaksiyon sonunda oluşan ürün, ortamdaki iyonların tabiatı, konsantrasyonları, allosterik etki, ışık ve diğer fiziksel faktörler, hormonlar ve diğer biyokimyasal faktörler olarak söylenebilir.
 AKTİVATÖRLER VE İNHİBİTÖRLER: Çoğu enzimlerin maksimum aktivite için spesifik iyonlara ihtiyacı vardır. Heksokinaz gibi bütün fosfat transfer eden enzimlerin Mg+2 iyonlarına ihtiyacı vardır. Diğer metal iyon aktiviteleri de Mn+2 , Ca+2, Zn+2, Fe+2 , K+ dir. Bazı enzimlerinde maksimum aktivite için birkaç iyona birden ihtiyaçları vardır. Her durumda aktivatörün optimal konsantrasyonu, substrat konsantrasyonu da optimal olduğu zaman belirlenmelidir.
AKTİVATÖRLER VE İNHİBİTÖRLER: Çoğu enzimlerin maksimum aktivite için spesifik iyonlara ihtiyacı vardır. Heksokinaz gibi bütün fosfat transfer eden enzimlerin Mg+2 iyonlarına ihtiyacı vardır. Diğer metal iyon aktiviteleri de Mn+2 , Ca+2, Zn+2, Fe+2 , K+ dir. Bazı enzimlerinde maksimum aktivite için birkaç iyona birden ihtiyaçları vardır. Her durumda aktivatörün optimal konsantrasyonu, substrat konsantrasyonu da optimal olduğu zaman belirlenmelidir.
 Regulation of Enzyme Activity Inhibitor Proteolysis or proteolysis inhibitor Feedback regulation Phosophorylation P R R regulator effector (+) P Signal transduction or (-) Regulatory subunit + c. AMP or calmodulin Juang RH (2004) BCbasics phosphorylation
Regulation of Enzyme Activity Inhibitor Proteolysis or proteolysis inhibitor Feedback regulation Phosophorylation P R R regulator effector (+) P Signal transduction or (-) Regulatory subunit + c. AMP or calmodulin Juang RH (2004) BCbasics phosphorylation
 İnhibitörler bir enzimin katalitik aktivitesini azaltan maddelerdir. Bir çok inhibitör tipi ve birkaç inhibisyon sınıfı vardır. İnhibitörler, kelat teşkil ederek bir enzimin aktivitesini ortadan kaldırabilirler. Ör; Ca+2 ve Mg+2 nin etkisini EDTA ortadan kaldırır. Hekzokinazın inhibisyonu da okzalat tarafından gerçekleştirilir. Bunlar etkilerini substratla yarışarak aktif yere bağlanarak gösterebildikleri gibi, enzim aktivitesinde etkili olan allosterik yer gibi bir yerde kompleks oluşturarak ta gösterebilirler.
İnhibitörler bir enzimin katalitik aktivitesini azaltan maddelerdir. Bir çok inhibitör tipi ve birkaç inhibisyon sınıfı vardır. İnhibitörler, kelat teşkil ederek bir enzimin aktivitesini ortadan kaldırabilirler. Ör; Ca+2 ve Mg+2 nin etkisini EDTA ortadan kaldırır. Hekzokinazın inhibisyonu da okzalat tarafından gerçekleştirilir. Bunlar etkilerini substratla yarışarak aktif yere bağlanarak gösterebildikleri gibi, enzim aktivitesinde etkili olan allosterik yer gibi bir yerde kompleks oluşturarak ta gösterebilirler.
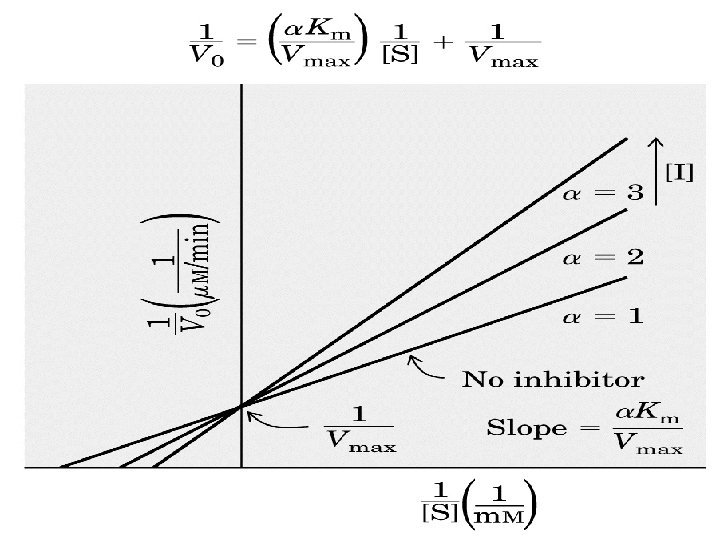
Kompetatif inhibitörler 2 -)Kompetatif olmayan inhibitörler(Unkompetative) 3 -)Non") İnhibitörler, başlıca 3 gruba ayrılırlar; 1 -)Kompetatif inhibitörler 2 -)Kompetatif olmayan inhibitörler(Unkompetative) 3 -)Non kompetatif inhibitörler 1 -)KOMPETATİF İNHİBİTÖRLER: I aktif yere bağlanmada S ile yarışır. Reversible dır. Artan [S] azaltılabilir. Vmax etkilenmez çünkü reversibledır. Km substrat etkilendiğinden artar.
İnhibitörler, başlıca 3 gruba ayrılırlar; 1 -)Kompetatif inhibitörler 2 -)Kompetatif olmayan inhibitörler(Unkompetative) 3 -)Non kompetatif inhibitörler 1 -)KOMPETATİF İNHİBİTÖRLER: I aktif yere bağlanmada S ile yarışır. Reversible dır. Artan [S] azaltılabilir. Vmax etkilenmez çünkü reversibledır. Km substrat etkilendiğinden artar.
 Aynı Bağlanma Yeri S I B-) Sterik Engelleme: I S") A-) Aynı Bağlanma Yeri S I B-) Sterik Engelleme: I S
A-) Aynı Bağlanma Yeri S I B-) Sterik Engelleme: I S
 Aynı bağlanma yerini paylaşma: I D-) Overlapping: S") C-) Aynı bağlanma yerini paylaşma: I D-) Overlapping: S
C-) Aynı bağlanma yerini paylaşma: I D-) Overlapping: S
 Baskılama Bağlanma yerini yapısal değiştirme
Baskılama Bağlanma yerini yapısal değiştirme
 Kompetatif İnhibitörler
Kompetatif İnhibitörler
 Competitive Inhibition Ürün Substrat Suksinat C-OO- Competitive Inhibitor Glutarat C-OO- C-H H-C-H C-OO- Malonat C-OOH-C-H C-OO- Süksinat dehidrogenaz Oxalat C-OO-
Competitive Inhibition Ürün Substrat Suksinat C-OO- Competitive Inhibitor Glutarat C-OO- C-H H-C-H C-OO- Malonat C-OOH-C-H C-OO- Süksinat dehidrogenaz Oxalat C-OO-
 Para-aminobenzoic acid (PABA) H 2 N- -COOH") Sulfa Drug Is Competitive Inhibitor Domagk (1939) Para-aminobenzoic acid (PABA) H 2 N- -COOH Bacteria needs PABA for the biosynthesis of folic acid Folic acid Precursor H 2 N- -SONH 2 Sulfanilamide Sulfa drug (anti-inflammation) Tetrahydrofolic acid Sulfa drugs has similar structure with PABA, and inhibit bacteria growth.
Sulfa Drug Is Competitive Inhibitor Domagk (1939) Para-aminobenzoic acid (PABA) H 2 N- -COOH Bacteria needs PABA for the biosynthesis of folic acid Folic acid Precursor H 2 N- -SONH 2 Sulfanilamide Sulfa drug (anti-inflammation) Tetrahydrofolic acid Sulfa drugs has similar structure with PABA, and inhibit bacteria growth.
 Competitive Inhibition EI Competitive S CI E Kic S+E + I ES E+P Kmapp/Vmax Kmapp -1/Kmapp
Competitive Inhibition EI Competitive S CI E Kic S+E + I ES E+P Kmapp/Vmax Kmapp -1/Kmapp
 Kompetatif İnhibitörler PABA Sulfanilamide PABA precursor to folic acid in bacteria O 2 C-CH 2 -CO 2 -------> O 2 C-CH=CH-CO 2 succinate fumarate Succinate dehydrogenase O 2 C-CH 2 -CO 2 Malonate
Kompetatif İnhibitörler PABA Sulfanilamide PABA precursor to folic acid in bacteria O 2 C-CH 2 -CO 2 -------> O 2 C-CH=CH-CO 2 succinate fumarate Succinate dehydrogenase O 2 C-CH 2 -CO 2 Malonate
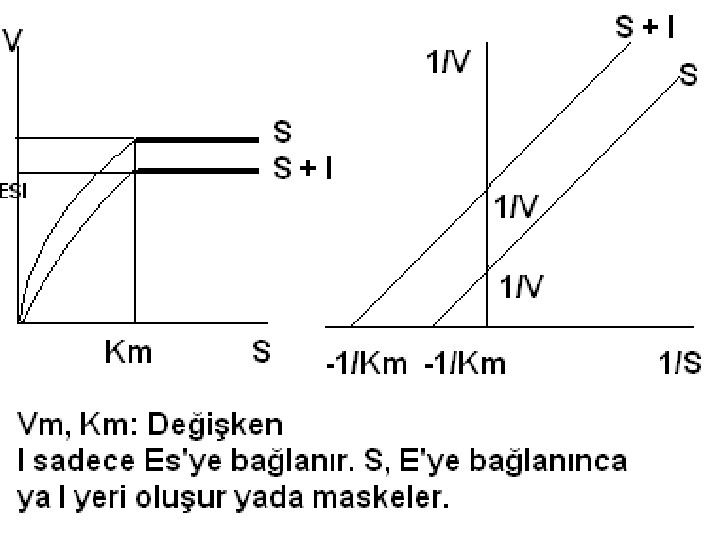
 Kompetatif Olmayan İnhibitörler (Unkompetatif) Serbest enzime bağlanma yoktur Ortama fazla S ilave edilince") 2) Kompetatif Olmayan İnhibitörler (Unkompetatif) Serbest enzime bağlanma yoktur Ortama fazla S ilave edilince inhibisyon artacaktır. Çünkü daha fazla ES oluşacak ve buda I ile ESI oluşacaktır. Burada Vmax azalır, Km düşer.
2) Kompetatif Olmayan İnhibitörler (Unkompetatif) Serbest enzime bağlanma yoktur Ortama fazla S ilave edilince inhibisyon artacaktır. Çünkü daha fazla ES oluşacak ve buda I ile ESI oluşacaktır. Burada Vmax azalır, Km düşer.
 Unkompetatif İnhibitörler
Unkompetatif İnhibitörler
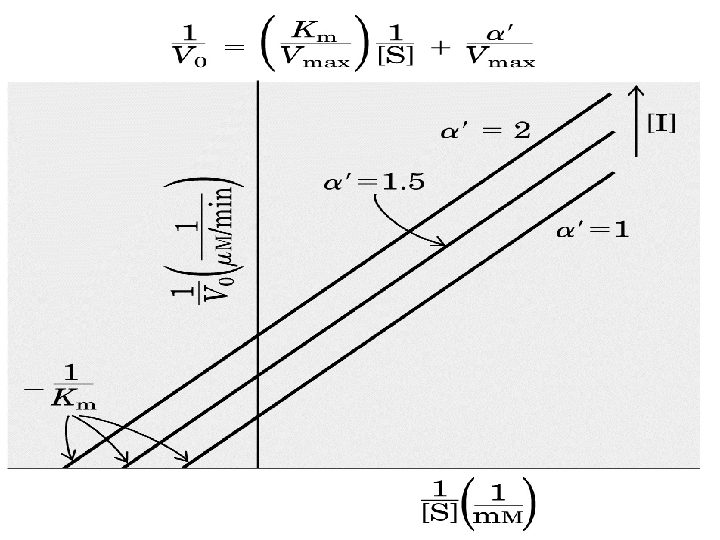
 S E UCI S+E ESI Kiu ES + I E+P") Uncompetitive Inhibition Uncompetitive (catalytic) S E UCI S+E ESI Kiu ES + I E+P 1/Vmaxapp 0. 5 Vmax Kmapp/Vmaxapp Kmapp -1/Kmapp
Uncompetitive Inhibition Uncompetitive (catalytic) S E UCI S+E ESI Kiu ES + I E+P 1/Vmaxapp 0. 5 Vmax Kmapp/Vmaxapp Kmapp -1/Kmapp
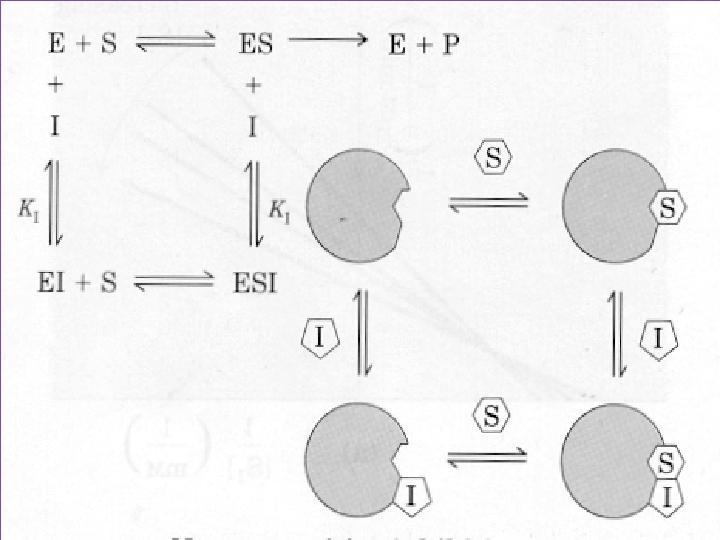
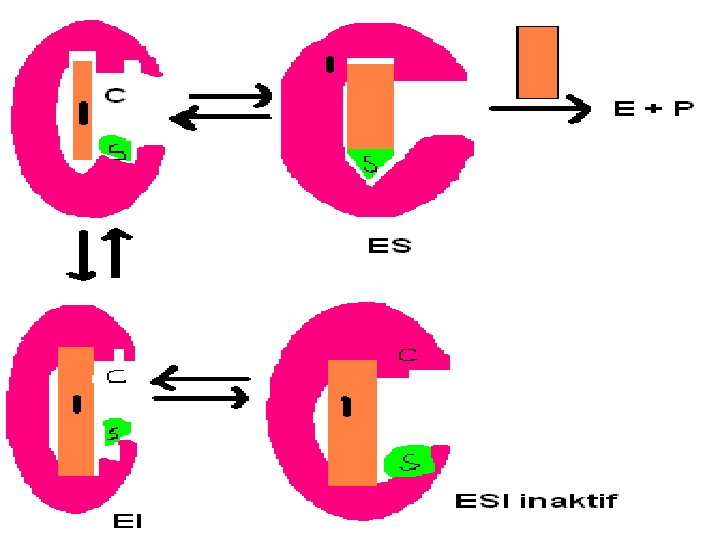
 Yarışmasız İnhibisyon (Nonkompetatif): Allosterik veya regülatör yere bağlanır. S ilavesi ile reverzibl olmaz.") 3) Yarışmasız İnhibisyon (Nonkompetatif): Allosterik veya regülatör yere bağlanır. S ilavesi ile reverzibl olmaz. Çünkü enzim üzerinde farklı yere bağlanır. I ile S arasında fark yoktur. Km etkilenmez. Ortamdaki E ve ES azalacağı için Vm azalır. E+S + I EI + S ESI E+P Vm: Değişken Km: Sabit
3) Yarışmasız İnhibisyon (Nonkompetatif): Allosterik veya regülatör yere bağlanır. S ilavesi ile reverzibl olmaz. Çünkü enzim üzerinde farklı yere bağlanır. I ile S arasında fark yoktur. Km etkilenmez. Ortamdaki E ve ES azalacağı için Vm azalır. E+S + I EI + S ESI E+P Vm: Değişken Km: Sabit
 malathion parathion • Organofosfatlar • Inhibit serin hidrolaz") Irreversible Inhibitörler Diisopropyl fluorophosphate (nerve gas) malathion parathion • Organofosfatlar • Inhibit serin hidrolaz • Asetilkolinesteraz inhibitörleri
Irreversible Inhibitörler Diisopropyl fluorophosphate (nerve gas) malathion parathion • Organofosfatlar • Inhibit serin hidrolaz • Asetilkolinesteraz inhibitörleri
 Non-competitive Inhibitor
Non-competitive Inhibitor
 S S NCI E E NCI EI Kic S+E +") Noncompetitive Inhibition Noncompetitive (mixed-type) S S NCI E E NCI EI Kic S+E + I ESI Kiu ES + I 1/Vmaxapp 0. 5 Vmax Km/Vmaxapp E+P
Noncompetitive Inhibition Noncompetitive (mixed-type) S S NCI E E NCI EI Kic S+E + I ESI Kiu ES + I 1/Vmaxapp 0. 5 Vmax Km/Vmaxapp E+P
 KOMPETATİF İNHİBİTÖRLER
KOMPETATİF İNHİBİTÖRLER
 UNKOMPETATİF İNHİBİTÖRLER NONKOMPETATİF İNHİBİTÖRLER") c) UNKOMPETATİF İNHİBİTÖRLER NONKOMPETATİF İNHİBİTÖRLER
c) UNKOMPETATİF İNHİBİTÖRLER NONKOMPETATİF İNHİBİTÖRLER
 Equation and Description Cartoon Guide Competitive Non-competitive Uncompetitive Substrate Compete for") Enzyme Inhibition (Mechanism) Equation and Description Cartoon Guide Competitive Non-competitive Uncompetitive Substrate Compete for Inhibitor active site E E X Different site E + S ← ES → E + P → + I ↑ ↓ EI E + S ← ES → E + P → + + I I ↑ ↓ ↓↑ EI + S →EIS E + S ← ES → E + P → + I ↑ ↓ E IS [I] binds to free [E] only, and competes with [S]; increasing [S] overcomes Inhibition by [I] binds to free [E] or [ES] complex; Increasing [S] can not overcome [I] inhibition. [I] binds to [ES] complex only, increasing [S] favors the inhibition by [I]. Juang RH (2004) BCbasics
Enzyme Inhibition (Mechanism) Equation and Description Cartoon Guide Competitive Non-competitive Uncompetitive Substrate Compete for Inhibitor active site E E X Different site E + S ← ES → E + P → + I ↑ ↓ EI E + S ← ES → E + P → + + I I ↑ ↓ ↓↑ EI + S →EIS E + S ← ES → E + P → + I ↑ ↓ E IS [I] binds to free [E] only, and competes with [S]; increasing [S] overcomes Inhibition by [I] binds to free [E] or [ES] complex; Increasing [S] can not overcome [I] inhibition. [I] binds to [ES] complex only, increasing [S] favors the inhibition by [I]. Juang RH (2004) BCbasics
 Direct Plots Competitive Non-competitive Uncompetitive Vmax vo vo I Km Km’") Enzyme Inhibition (Plots) Direct Plots Competitive Non-competitive Uncompetitive Vmax vo vo I Km Km’ Double Reciprocal Vmax I [S], m. M Km = Km’ Vmax’ [S], m. M I Km’ Km [S], m. M Vmax unchanged Km increased Vmax decreased Km unchanged Both Vmax & Km decreased 1/vo Intersect at Y axis 1/Km I I I Two parallel lines 1/ Vmax 1/[S] Intersect at X axis 1/Km 1/ Vmax 1/[S] 1/ Vmax 1/Km 1/[S] Juang RH (2004) BCbasics
Enzyme Inhibition (Plots) Direct Plots Competitive Non-competitive Uncompetitive Vmax vo vo I Km Km’ Double Reciprocal Vmax I [S], m. M Km = Km’ Vmax’ [S], m. M I Km’ Km [S], m. M Vmax unchanged Km increased Vmax decreased Km unchanged Both Vmax & Km decreased 1/vo Intersect at Y axis 1/Km I I I Two parallel lines 1/ Vmax 1/[S] Intersect at X axis 1/Km 1/ Vmax 1/[S] 1/ Vmax 1/Km 1/[S] Juang RH (2004) BCbasics
 1 2 3 4 5 6 7 8 80 60 Ürün Substrat Konsantrasyonunun Artışı 0 40 20 0 0 2 4 6 Substrat(mm) 8 S + E ↓ P
1 2 3 4 5 6 7 8 80 60 Ürün Substrat Konsantrasyonunun Artışı 0 40 20 0 0 2 4 6 Substrat(mm) 8 S + E ↓ P
 E E S + S E+S k 1 k-1 ES E k 2 k-2 E+P + P
E E S + S E+S k 1 k-1 ES E k 2 k-2 E+P + P
![E E S + S E+S k 1 ES Rate = k 1 [E]](https://present5.com/presentation/5974a95168b019f4c5905e4bc2af1b5a/image-102.jpg "E E S + S E+S k 1 ES Rate = k 1 [E]") E E S + S E+S k 1 ES Rate = k 1 [E] [S]
E E S + S E+S k 1 ES Rate = k 1 [E] [S]
 E S ES E k 2 ES k-1 P E+P E E S + + S E+S Rate = (k 2 [ES]) + (k-1[ES]) Rate = [ES](k 2 + k-1)
E S ES E k 2 ES k-1 P E+P E E S + + S E+S Rate = (k 2 [ES]) + (k-1[ES]) Rate = [ES](k 2 + k-1)
![Km = [S]. ½ Vmax](https://present5.com/presentation/5974a95168b019f4c5905e4bc2af1b5a/image-104.jpg "Km = [S]. ½ Vmax") Km = [S]. ½ Vmax
Km = [S]. ½ Vmax
 1 k 12 2 k 23 3 E+S ES E+P k 21 k 12: ES’nin oluşması k 23: Es’nin yıkılması k 12 k 23 1. E + S ES E+P k 21 2. V= k 23 [ES] 3. ES= [E] ES= k 12. E. S Substrat sonsuz ise 4. Vm= k 23 [Et] Enzim substrata doyduğu Vm
1 k 12 2 k 23 3 E+S ES E+P k 21 k 12: ES’nin oluşması k 23: Es’nin yıkılması k 12 k 23 1. E + S ES E+P k 21 2. V= k 23 [ES] 3. ES= [E] ES= k 12. E. S Substrat sonsuz ise 4. Vm= k 23 [Et] Enzim substrata doyduğu Vm
 5. V/Vm = ES/Et 6. ES oluşması ES. k 12 = d. ES/dt 7. ES(k 21 + k 23)= d. ES/dt Es’nin yıkımı 8. d. ES/dt= E. S. k 12 - ES(k 21 + k 23) Birikme Es oluşma hızı ile Es yıkılma hızı aynı ise Es oranı sabittir, değişmez. 9. E. S. k 12 = ES(k 21 + k 23) 10. E. S/ES= (k 21 + k 23)/ k 12 =Km=Keq Km= k 21 + k 23/ k 12 Michealis/Menten Sabiti 11. Et= ES + E E= Et – ES
5. V/Vm = ES/Et 6. ES oluşması ES. k 12 = d. ES/dt 7. ES(k 21 + k 23)= d. ES/dt Es’nin yıkımı 8. d. ES/dt= E. S. k 12 - ES(k 21 + k 23) Birikme Es oluşma hızı ile Es yıkılma hızı aynı ise Es oranı sabittir, değişmez. 9. E. S. k 12 = ES(k 21 + k 23) 10. E. S/ES= (k 21 + k 23)/ k 12 =Km=Keq Km= k 21 + k 23/ k 12 Michealis/Menten Sabiti 11. Et= ES + E E= Et – ES
 Michaelis-Menten E+S Vmax Km Kcat/Km k 1 k-1 ES kcat E+P • Vmax • Vo = Vmax [S] • K m Km + [S] • kcat/Km
Michaelis-Menten E+S Vmax Km Kcat/Km k 1 k-1 ES kcat E+P • Vmax • Vo = Vmax [S] • K m Km + [S] • kcat/Km
![12. [Et - ES]. S/ES= Km 13. V= Vm. [S] / [Km] + [S]](https://present5.com/presentation/5974a95168b019f4c5905e4bc2af1b5a/image-108.jpg "12. [Et - ES]. S/ES= Km 13. V= Vm. [S] / [Km] + [S]") 12. [Et - ES]. S/ES= Km 13. V= Vm. [S] / [Km] + [S] 14. 1/V=1/Vm + 1/S. Km/V Lineweaver Burk Denklemi 15. Vm= V. Km/S + V V= Vm – Km. V/S Eadie Hofstee Bağıntısı V/[S]= -V/Km + V/Km
12. [Et - ES]. S/ES= Km 13. V= Vm. [S] / [Km] + [S] 14. 1/V=1/Vm + 1/S. Km/V Lineweaver Burk Denklemi 15. Vm= V. Km/S + V V= Vm – Km. V/S Eadie Hofstee Bağıntısı V/[S]= -V/Km + V/Km
 Lineweaver-Burk Eğrisi
Lineweaver-Burk Eğrisi

 İki substratlı enzimatik tepkimelerde E-S ilişkisi: İki S’nin E’ye bağlanabildiği enzimatik tepkimeler için, iki model öne sürülmüştür; S 1 ve S 2 substrat, P 1 ve P 2 ise ürünleri göstermektedir. E S 1+S 2 P 1+P 2 1 -) Tek substrat tek ürün(uni-uni) reaksiyonunun gösterilişi: S Ü E ES (ES EÜ) E
İki substratlı enzimatik tepkimelerde E-S ilişkisi: İki S’nin E’ye bağlanabildiği enzimatik tepkimeler için, iki model öne sürülmüştür; S 1 ve S 2 substrat, P 1 ve P 2 ise ürünleri göstermektedir. E S 1+S 2 P 1+P 2 1 -) Tek substrat tek ürün(uni-uni) reaksiyonunun gösterilişi: S Ü E ES (ES EÜ) E
İki substrat iki product (ürün) düzensiz (Bi. Bi) reaksiyonunun gösterilişi: S 1 S") 2 -)İki substrat iki product (ürün) düzensiz (Bi. Bi) reaksiyonunun gösterilişi: S 1 S 2 ES 1 Ü 2 EÜ 1 ES 2 E EÜ 2 S 1 Ü 2 (DÜZENSİZ BİNARY KOMPLEKS)
2 -)İki substrat iki product (ürün) düzensiz (Bi. Bi) reaksiyonunun gösterilişi: S 1 S 2 ES 1 Ü 2 EÜ 1 ES 2 E EÜ 2 S 1 Ü 2 (DÜZENSİZ BİNARY KOMPLEKS)
 İki substrat iki product (ürün) düzenli (Bi. Bi) reaksiyonunun gösterilişi: S 1") 3 -) İki substrat iki product (ürün) düzenli (Bi. Bi) reaksiyonunun gösterilişi: S 1 S 2 Ü 1 Ü 2 E ES 1 (ES 1 S 2 EÜ 1 EÜ 2) (DÜZENLİ BİNARY) EÜ 2 E
3 -) İki substrat iki product (ürün) düzenli (Bi. Bi) reaksiyonunun gösterilişi: S 1 S 2 Ü 1 Ü 2 E ES 1 (ES 1 S 2 EÜ 1 EÜ 2) (DÜZENLİ BİNARY) EÜ 2 E
 Ternary (üçlü) kompleks: a-) Gelişi güzel (Randomly (EAB, EPQ)): A EA E") 4 -) Ternary (üçlü) kompleks: a-) Gelişi güzel (Randomly (EAB, EPQ)): A EA E A EAB P Q EQ EPQ E EB B B Q EP P b-) Düzenli (Ordered) (EA, EB, EP, EQ): Q P E+A EA + B EAB EPQ EQ E
4 -) Ternary (üçlü) kompleks: a-) Gelişi güzel (Randomly (EAB, EPQ)): A EA E A EAB P Q EQ EPQ E EB B B Q EP P b-) Düzenli (Ordered) (EA, EB, EP, EQ): Q P E+A EA + B EAB EPQ EQ E
 Bi-Bi, Ping-Pong Mekanizması : A P B E EA FP F FB") 5 -) Bi-Bi, Ping-Pong Mekanizması : A P B E EA FP F FB Q EQ Ping-Pong Reaksiyonu A E P (EA)(FP) B (F) Q (FB)(EQ) E E
5 -) Bi-Bi, Ping-Pong Mekanizması : A P B E EA FP F FB Q EQ Ping-Pong Reaksiyonu A E P (EA)(FP) B (F) Q (FB)(EQ) E E
 E
E
