
Энергетический метаболизм прокариот Метаболизм










, окислительный пентозофосфатный путь, путь")


, служит для обозначения макроэргической связи.")








 • Освобождающаяся припреносе электронов энергия первоначально запасается в форме ΔμН+ • Разрядка")



 аэробы развиваются при наличии в")



energ.metabolizm.ppt
- Размер: 1.7 Мб
- Автор:
- Количество слайдов: 31
Описание презентации Энергетический метаболизм прокариот Метаболизм по слайдам
Энергетический метаболизм прокариот
Метаболизм – совокупность ферментативных процессов, протекающих в клетке и обеспечивающих её энергетические и биосинтетические потребности. Энергетический метаболизм (катаболизм) – поток реакций, сопровождающийся мобилизацией энергии и преобразованием её в электрохимическую или химическую форму, которая затем используется во всех энергозависимых процессах. Конструктивный метаболизм (биосинтез, анаболизм) – поток реакций, в результате которых за счет поступающих извне веществ строится вещество клетки и при этом используется запасённая клеткой энергия.
Классификация микроорганизмов По источнику углерода Автотрофы Гетеротрофы СО 2 органические соединения В зависимости от донора электронов. По источнику энергии Литотрофы Органотрофы. Фототрофы Хемотрофы свет Энергия химических связей питаются мертвым органическим материалом и независимы от других организмов получают питательные вещества от макроорганизма
Факторы роста – вещества, которые прокариоты по каким-либо причинам не могут синтезировать самостоятельно из используемого источника углерода. К ним относятся аминокислоты, пурины , пиримидины, витамины и др. Такие вещества добавляют в питательные среды в готовом виде в небольших количествах. Микроорганизмы, которым в дополнение к основному источнику углерода необходимы факторы роста, называются ауксотрофы. Микроорганизмы, которые синтезируют все необходимые органические соединения из основного источника углерода самостоятельно, называются прототрофы.
Бактерии, изучаемые медицинской микробиологией- хемоорганогетеротрофы Отличительной особенностью этой группы является то, что источник углерода у них является источником энергии.
• В общем виде процессы, способные служить источником энергии для прокариот, можно представить следующим образом Должен существовать энергетический ресурс — исходный субстрат С помощью ферментных систем организм извлекает энергию из этого субстрата в реакциях его ступенчатого окисления
Способность использовать химическую энергию присуща всем без исключения организмам. Особенно многообразны возможности прокариот. Мономеры, образованные при расщеплении биополимеров, должны быть ферментативным путем трансформированы в молекулы, способные включаться в качестве метаболитов в функционирующие клеточные катаболические системы
Основные катаболические системы клетки: гликолиз (путь Эмбдена-Мейергофа-Парнаса), окислительный пентозофосфатный путь, путь Энтнера-Дудорова цикл трикарбоновых кислот (ЦТК) Общее для всех катаболических путей – многоступенчатость процесса окисления исходного субстрата На некоторых этапах окисление субстрата сопряжено с образованием энергии в той форме, в которой она может быть использована клеткой
Схема энергетических процессов у прокариот.
Символ «~», введенный американским биохимиком Ф. Липманом (F. Lipmann), служит для обозначения макроэргической связи.
Брожение – наиболее примитивный способ получения энергии Донор и акцептор электронов – органические вещества, т. е. из субстрата в результате его анаэробного преобразования извлекается лишь незначительная доля той химической энергии, которая в нем содержится. Процесс протекает только в анаэробных условиях Энергия запасается в молекулах АТФ в реакциях субстратного фосфорилирования. Все реакции катализируются растворимыми ферментами. Энергетический выход: при окислении 1 молекулы глюкозы в среднем образуется 2 молекулы АТФ
Продукты брожения глюкозы
Муравьинокислое брожение и семейство Enterobacteriaceae Некоторые микроорганизмы, образующие при брожении кислоты, объединяют в одну физиологическую группу на том основании, что характерным, хотя и не главным продуктом брожения является у них муравьиная кислота. Наряду с муравьиной кислотой такие бактерии выделяют и некоторые другие кислоты; такой тип метаболизма называют поэто му муравьинокислым брожением или брожением смешанного типа. Так как некоторые типичные представители этой группы обитают в кишечнике, все семейство носит название Enterobacteriaceae. Будучи факультативными аэробами, они обладают гемопротеинами (цитохромами и каталазой) и способны получать энергию как в процессе дыхания (в аэробных условиях), так и в процессе брожения (в анаэробных условиях).
1. Спиртовое брожение дрожжи, грибы рода Mucor 2. Молочнокислое брожение. Типичное (гомоферментативное) молочнокислое брожение — процесс расщепления углеводов (лактозы, мальтозы, сахарозы, глюкозы и др. ) с образованием молочной кислоты без побочных продуктов Род Streptococcus, Отдельные представители рода Lactobacillus Нетипичное (гетероферментативное) молочнокислое брожение происходит с образованием молочной кислоты, уксусной кислоты, этилового спирта и газообразных продуктов — углекислоты. Отдельные представители рода Lactobacillus 3. Пропионовокислое Propionibacterium spp. , Veillonella alcalescens, Clostridium propionicum 4. Маслянокислое Бактерии рода Clostridium 5. Муравьинокислое смешанного типа C емейство Enterobacteriaceae
В процессе дыхания происходит окисление восстановленных веществ с относительно низким окислительно-восстановительным потенциалом, образующихся в реакциях метаболизма или являющихся исходными субстратами (NADH 2, сукцинат, лактат и др. ) Окисление происходит в результате переноса электронов от донора к акцептору по градиенту редокс-потенциала через ряд последовательно функционирующих переносчиков, встроенных в мембрану – дыхательную электронотранспортную цепь
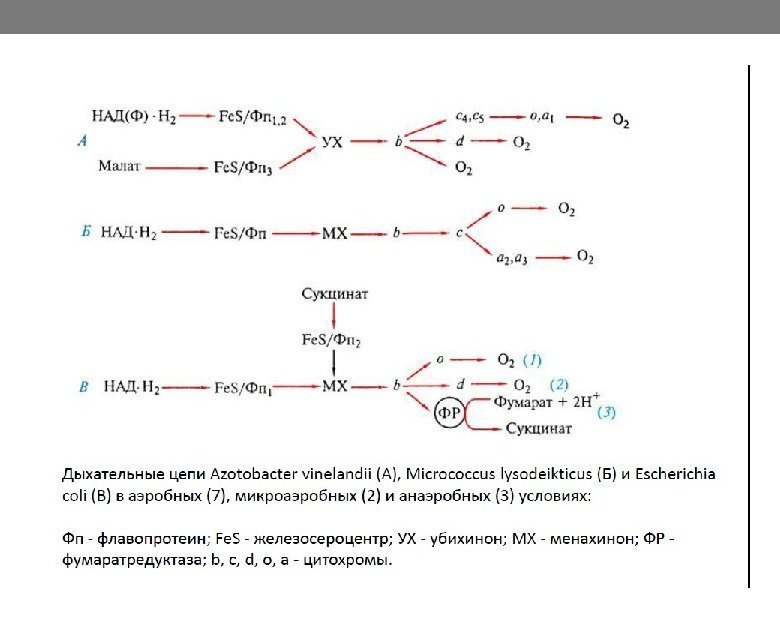
Организация дыхательной цепи
Расположение переносчиков электронов в ЦПМ прокариот таково, что при работе любой электронотранспортной цепи (фотосинтетической или дыхательной) во внешней среде происходит накопление ионов водорода (протонов), приводящее к подкислению среды, а в клеточной цитоплазме — их уменьшение, сопровождающееся ее подщелачиванием, таким образом при переносе электронов на ЦПМ возникает трансмембранный электрохимический градиент ионов водорода, обозначаемый символом ΔμН+
Дыхание (продолжение) • Освобождающаяся припреносе электронов энергия первоначально запасается в форме ΔμН+ • Разрядка ΔμН+ происходит с участием протонного АТФсинтазного комплекса • Локализованная в мембране АТФсинтаза катализирует реакции синтеза и гидролиза АТФ в соответствии с уравнением • • Реакция, протекающая слева направо, сопряжена с транспортом H+ по градиенту ΔμH+, при этом выделяется энергия, что приводит к разрядке градиента и синтезу АТФ. • Протекающая в противоположном направлении реакция гидролиза АТФ сопровождается выделением энергии и приводит к переносу Н+ против градиента, что приводит к образованию (или возрастанию) ΔμH+ на мембране. • Таким образом, АТФ-синтазный ферментный комплекс служит механизмом, обеспечивающим взаимное превращение двух форм клеточной энергии ( ΔμH+ и АТФ), устройством, сопрягающим процессы окислительной природы с фосфорилированием. .
Особенности дыхательных цепей прокариот • Дыхательная цепь бактерий: Расположена в цитоплазматической мембране Имеет непостоянный состав переносчиков ( возможна замена одних переносчиков другими; добавление или удаление, например , цитохрома с) Большинство бактерий имеют разветвленные дыхательные цепи на уровне цитохромов из-за большого потока электронов при избытке субстрата, чтобы не образовывалась электронная пробка Количество переносимых протонов на одну пару электронов варьирует у разных бактерий Может иметь вместо кислорода другие конечные акцепторы электронов.
Брожение Процесс протекает в цитоплазме Донор и акцептор электронов – органические метаболиты Анаэробный процесс Синтез АТФ в реакциях субстратного фосфорилирования Низкий энергетический выход Неполное использование энергии химических связей – конечные продукты: спирты, кислоты, кетоны Дыхание Дыхательная цепь локализована в ЦПМ Необходим кислород или другие внешние акцепторы электрона Протекает в аэробных или анаэробных условиях Синтез АТФ за счет трансмембранного градиента протонов Высокий энергетический выход
Типы анаэробного дыхания у эубактерий Энергетический процесс Конечный акцептор электронов Продукты восстановления Нитратное дыхание и денитрификация NO 3 – , NO 2 – , NO, N 2 Сульфатное и серное дыхание SO 4 2– , S 0 H 2 S Карбонатное дыхание CO 2 ацетат Фумаратное дыхание фумарат сукцинат. Различия дыхательных цепей аэробных и анаэробных бактерий – в конечном акцепторе электрона
Классификация прокариот по отношению к молекулярному кислороду Облигатные (строгие) аэробы развиваются при наличии в атмосфере 20% кислорода ( Pseudomonas aeruginosa, Vibrio cholera , Mycobacterium tuberculosis) Микроаэрофилы нуждаются в значительно меньшем количестве кислорода, его высокая концентрация хотя и не убивает бактерии, но задерживает их рост (Helicobacter pylori ), некоторые микробы нуждаются в повышенном содержании углекислого газа ( капнофилы — Neisseria )
Облигатные анаэробы — бактерии, для которых наличие молекулярного кислорода является губительным (Bacteroides, Clostridium) Факультативные анаэробы могут размножаться как в присутствии, так и в отсутствие кислорода (большинство патогенных и сапрофитных микробов) (E. coli) Аэротолерантные бактерии способны расти в присутствии кислорода, но не использовать его в качестве источника энергии. Энергию они получают исключительно с помощью брожения (Streptococcus) Классификация прокариот по отношению к молекулярному кислороду
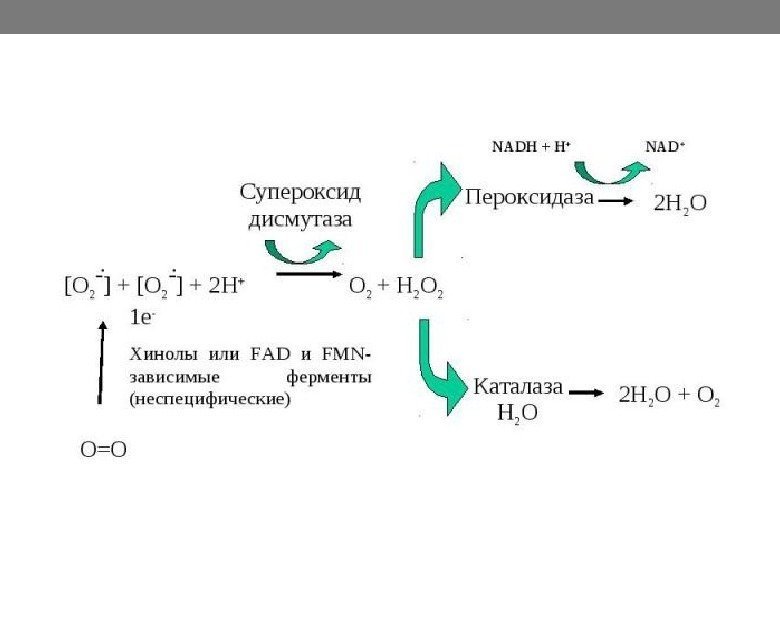
Молекулярный кислород. Токсичность Как фактор внешней среды О 2 воздействует на организмы двояко: с одной стороны, он может быть абсолютно необходимым, с другой — с молекулярным кислородом и его производными связаны токсические эффекты для клеток. Наибольшей токсичностью для клетки обладают синглетный кислород и гидроксидные радикалы. Определенный вклад в общий отрицательный кислородный эффект вносят и другие производные О 2.
Защитные механизмы клетки от кислорода и его токсических форм. Для нейтрализации токсических форм О 2 существующие прокариоты выработали различные защитные механизмы: деятельность специальных ферментов, для которых разложение токсических форм О 2 является основной и в ряде случаев единственной функцией ( супероксиддисмутаза (СОД), каталаза (КТ) и пероксидаза) способность пигментов или некоторых метаболитов выступать в качестве антиоксидантов