Лекция 4. Энергетический обмен прокариот.ppt
- Количество слайдов: 41



Энергетический метаболизм прокариот.

Энергия и работа. Энергия – способность совершать работу. Химическая работа – способность синтеза сложных молекул. Транспортная работа – поглощение питательных частиц, поддержание концентрации ионов, удаление отходов. Механическая работа – подвижность клетки и ее структур. d. G = d. H – T*d. S Свободная энергия системы (d. G) может быть использована для производства полезной работы. Энтальпия (d. H) – энергия, которая может быть переведена в теплоту. Энтропия (S) мера неупорядоченности системы. Отрицательная свободная энергия в экзотермических реакциях, положительная в эндотермических.

АТФ. Общие черты метаболизма: Использование АТФ в качестве хранилища энергии экзотермических реакций, используется для проведения эндотермических реакций АТФ + H 2 O = АДФ + Ф. Организация метаболических реакций в пути и циклы. Катализ реакций с помощью ферментов. Главную роль играют реакции окисления и восстановления.

Реакции первого типа – перенос фосфатной группы от молекулы-донора на АДФ. Пример – субстратное фосфорилирование на пути анаэробного превращения сахаров. Реакции второго типа – перенос энергии на промежуточный продукт реакции, сопряженный с образованием молекулы АТФ.
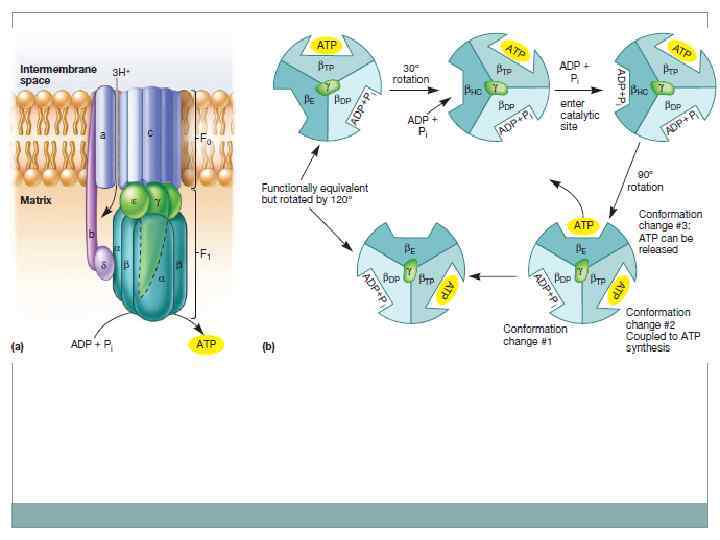

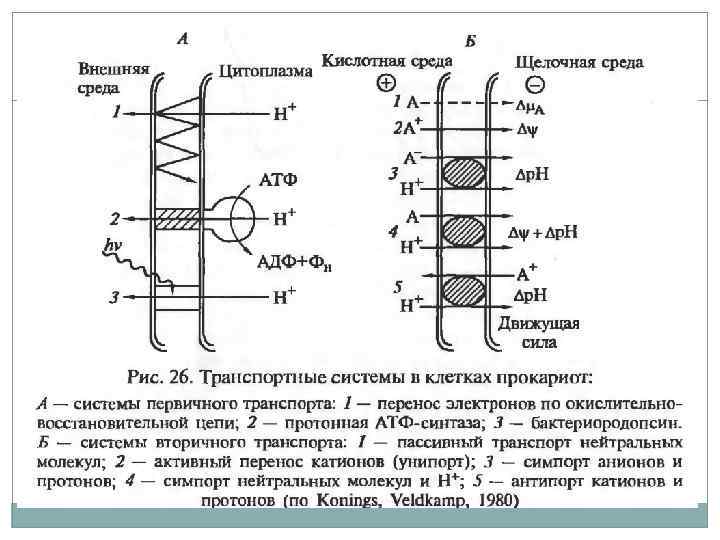
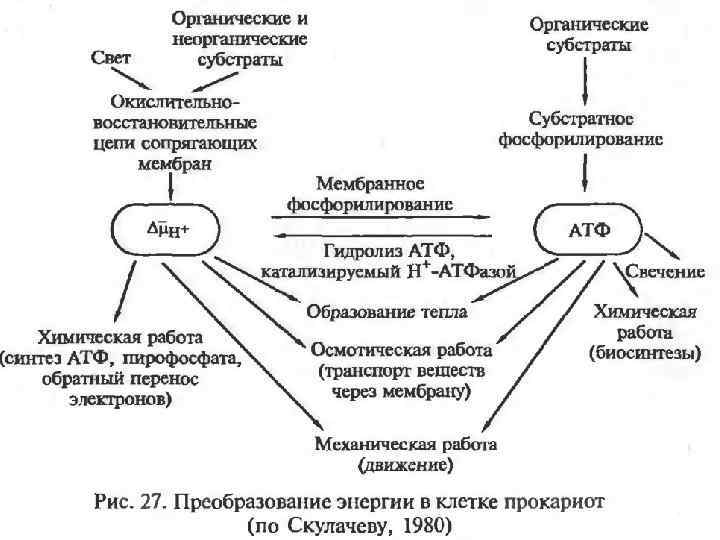
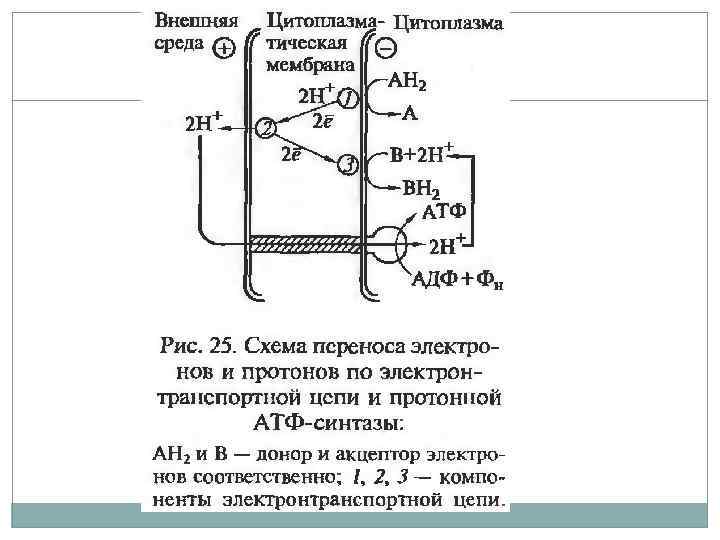
Электрохимический градиент. В 60 -х гг. английский биохимик Питер Митчелл разработал хемиосмотическую теорию энергетического сопряжения, преобразующуюся в энергию фосфатной связи. При переносе электронов по цепи ферментов происходит неравномерное распределение протонов по обе стороны мембраны. Во внешней среде происходит накопление избыточного положительного заряда – трансмембранный электрохимический градиент ионов водорода. На сопрягающих мембранах прокариот разность потенциалов достигает 200 – 250 м. В.

Хемиосмотическая теория.
, энергия химических связей (Хемотрофия). Источник углерода: органические")
Типы жизни прокариот. Источник энергии: свет (Фототрофия), энергия химических связей (Хемотрофия). Источник углерода: органические вещества (Гетеротрофы), углекислый газ (Автотрофы). Донор электронов: неорганические вещества (водород, сероводород, аммиак, железо) (Литотрофы), органические вещества (Органотрофы). Большая часть организмов относиться к хемоорганогетеротрофам. Облигатные организмы могут существовать используя только один тип питания, факультативные – осуществление данного типа не является облигатным (обязательным). Миксотрофы – способные использовать два источника углерода.
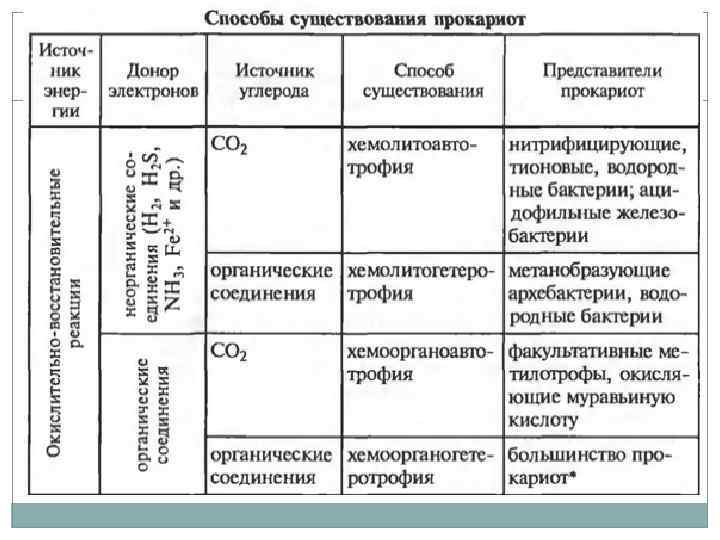
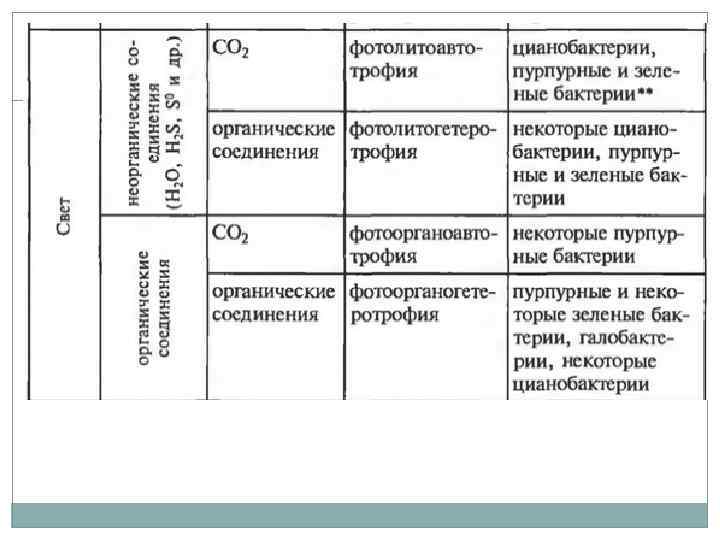
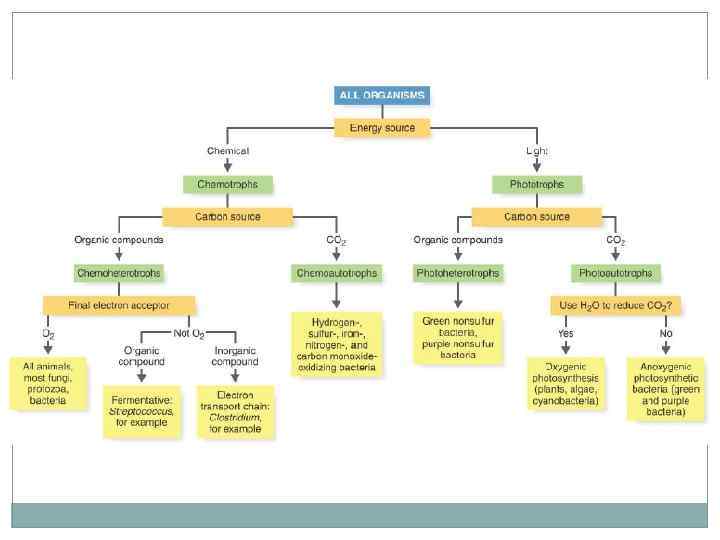

Питание микроорганизмов. Общие черты метаболизма: Использование АТФ в качестве хранилища энергии экзотермических реакций, используется для проведения эндотермических реакций АТФ + H 2 O = АДФ + Ф. Организация метаболических реакций в пути и циклы. Катализ реакций с помощью ферментов. Главную роль играют реакции окисления и восстановления.

Хемоорганотрофы. Основным способом существования является окисление органики и запасание энергии в виде АТФ. Электроны полученные при окислении органики переносятся по цепям транспорт на конечный акцептор. Если имеется внешний акцептор, такой процесс называют дыханием. Если акцептор один из метаболитов, такой процесс называется брожением. Аэробное дыхание – акцептор кислород. Анаэробное – акцептор нитрат-ион, сульфат-ион, углекислый газ, железо, селенит-ион. Потенциальная энергия протонов на мембране используется для синтеза АТФ. Эндогенный акцептор (например, пируват) используется для окисления органического субстрата, в этом случае АТФ синтезируется только в ходе субстратного фосфорилирования.
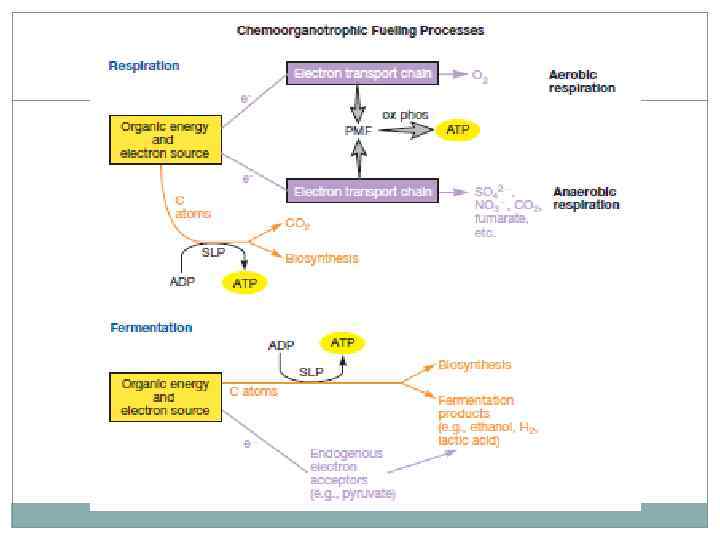

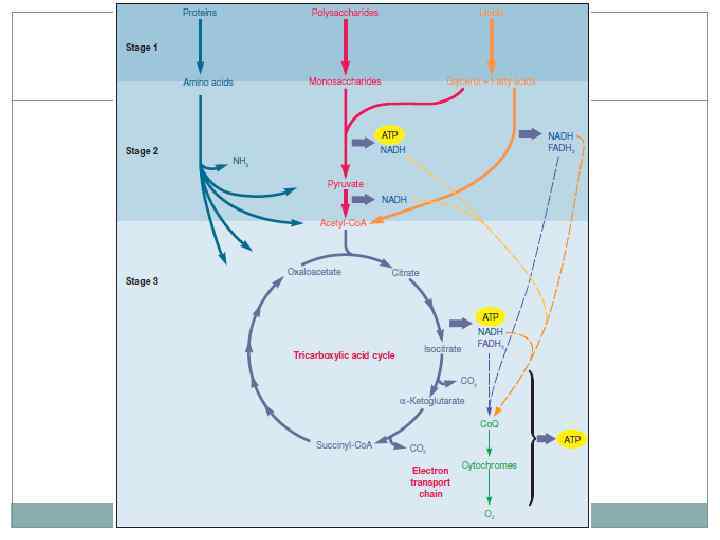
Аэробное дыхание. Процесс аэробного дыхания можно условно разделить на 3 стадии: 1. Гидролиз больших молекул питательных веществ. 2. Аминокислоты, моносахариды, жирные кислоты, глицерол формируют малые молекулы метаболитов – ацетил-Коа, пируват и др. 3. В цикле трикарбоновых кислот происходит окисление до углекислого газа, сопряженное с образованием АТФ, НАДН, ФАДН 2. Эффективность микроорганизмов достигается сведением большого числа питательных субстратов к небольшому числу метаболитов и нескольким путям их окисления. Многие пути являются амфиболическими и поставляют материал для анаболизма.
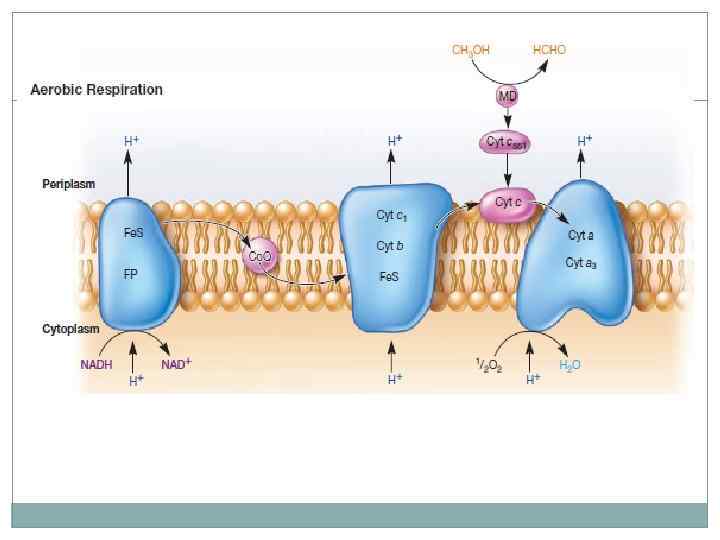
, пентозофосфатный путь и цикл трикарбоновых кислот.")
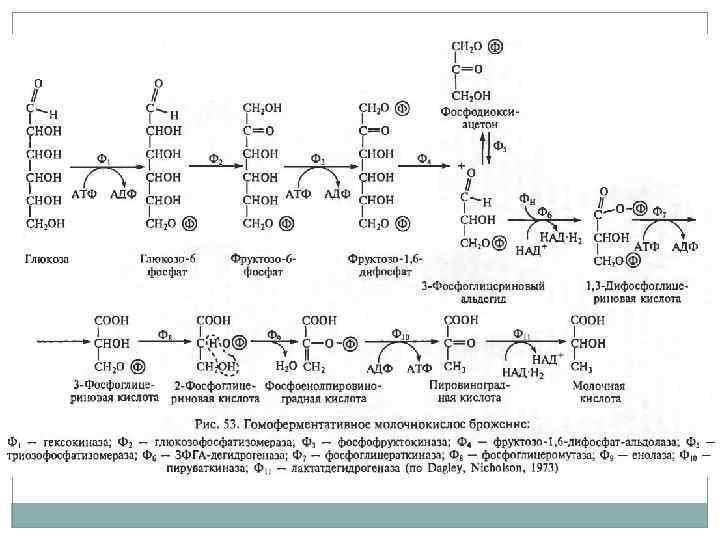
Аэробное дыхание. Наиболее важными являются пути Эмбдена-Мейерхофа (гликолиз), пентозофосфатный путь и цикл трикарбоновых кислот. Путь гликолиза можно разделить на 2 стадии: затрата 2 молекул АТФ с образованием фруктозо-1, 6 -бифосфата, далее фруктоза распадается на 2 молекулы глицеральдегид-3 фосфата, который окисляется до пирувата с образованием 4 молекул АТФ и 2 молекул НАД*Н. Процесс синтеза АТФ – субстратное фосфорилирование.
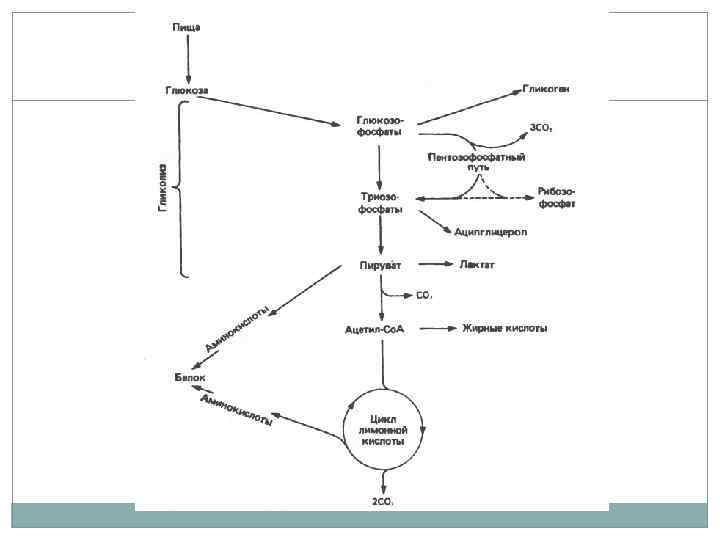

Гликолиз.

Гликолиз.

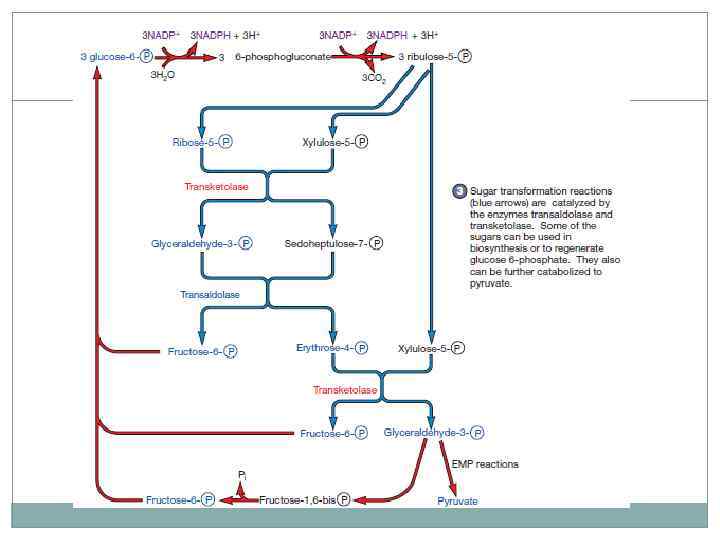
Пентозофосфатный путь встречается в аэробном и анаэробном метаболизме и участвует в процессе анаболизма и катаболизма. Окисление глюкозо-6 -фосфата до 6 -фосфоглюконата продолжается окислением до пентозы с выделением молекулы СО 2 с образованием 6 молекул НАДФ*Н. 2 молекулы пентозы затем образуют глюкозу и эритрозу. Присоединяя еще молекулу пентозы, регенирируется молекула фруктозы и глицеральдегида.

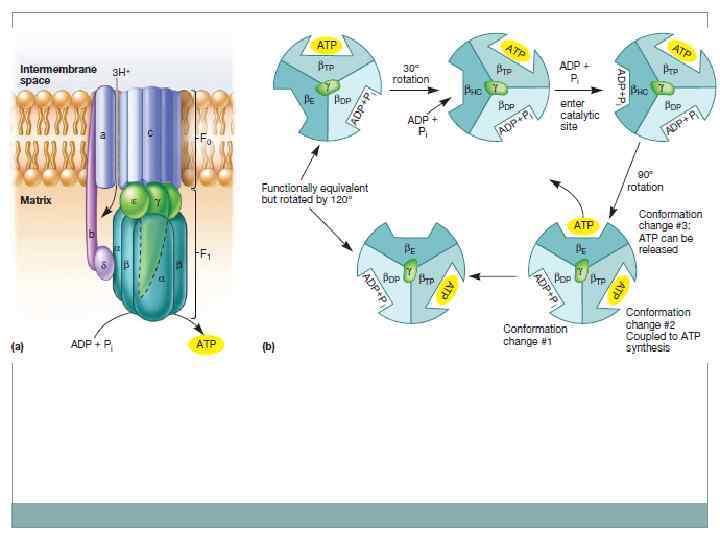
Цикл трикарбоновых кислот. Окисление молекул пирувата до углекислого газа. В отличии от гликолиза энергия переноситься молекулами НАД*Н которые передают электроны белкам электроно транспортной цепи. Электронный транспорт сопряжен с переносом протонов через мембрану с образованием электрохимического градиента. В конце цепи электрон попадает на молекулу кислорода с образованием воды. В процессе окислительного фосфорилирования образуются молекулы АТФ.
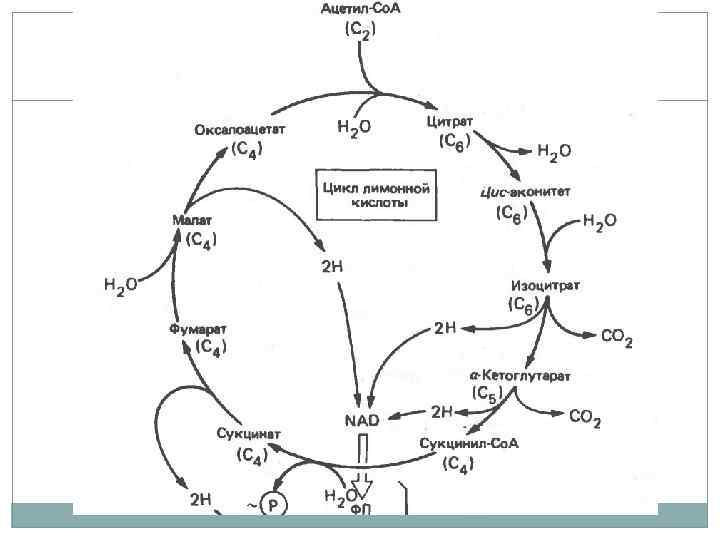
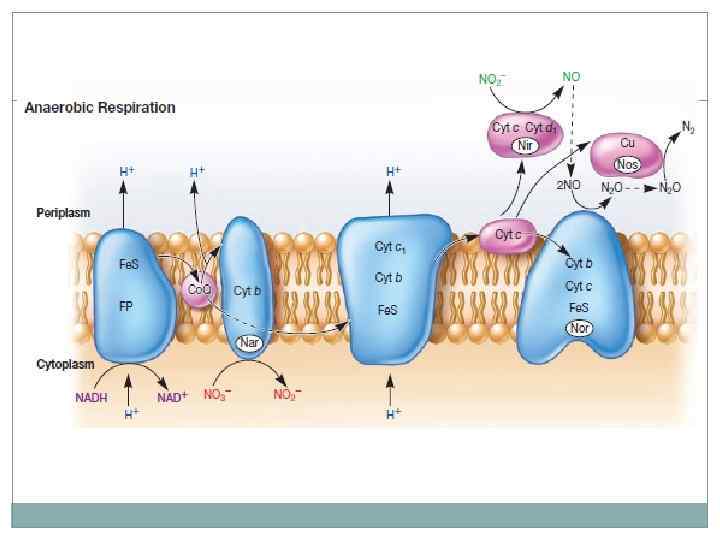

Кроме кислорода возможны другие акцепторы электронов, нитраты, сульфаты, СО 2. Многие бактерии могут использовать оба типа дыхания. Нитрат редукторы и денитрификаторы восстанавливают азот, метаногены восстанавливают СО 2, Desulfovibrio восстанавливают сульфаты с образованием сероводорода.

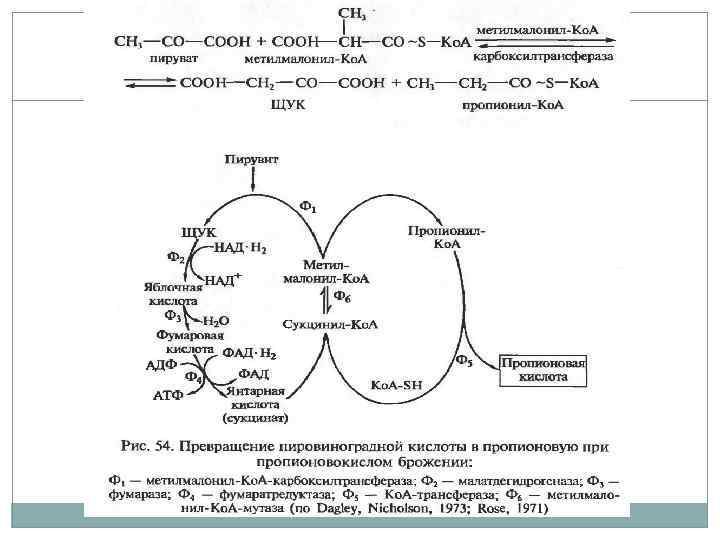
Брожение. Многие организмы лишены компонентов электронотранспортной цепи или находятся в анаэробных условиях. Гликолиз приводит к образованию восстановленного НАД, если нет возможности реализовать кислородный путь, клетки используют пути брожения, восстанавливая пируват до лактата или других продуктов. АТФ формируется только путем субстратного фосфорилирования. Алкогольное – этанол и СО 2. Молочнокислое – лактат. Смешанное кислотное – окисление муравьиной кислоты до СО 2 и Н 2.

Брожение.

Хемолитотрофы. Источником электронов для электронотранспортной цепи служат неорганические молекулы. Обычно акцептор кислород, но встречаются нитраты и сульфаты. Выход АТФ в сравнении с окислением глюкозы невелик и организмам приходится окислять большой объем неорганического материала. Некоторые способны окислять водород. Нитрифицирующие бактерии окисляют аммиак (Nitrosomonas).

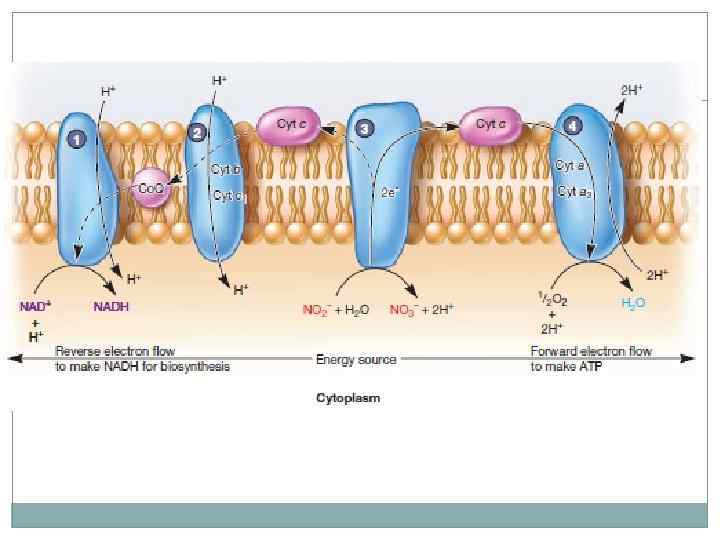
Хемолитотрофы. Многие хемолитотрофы не способны восстанавливать НАД напрямую, поскольку восстановительный потенциал реакций неорганического окисления ниже в сравнении с молекулой НАД. Это привело к использованию обратного потока электронов в транспортной цепи для восстановления НАД*Н для биосинтетических целей.
. Окисляют серу, сероводород и тиосульфат, используя субстратное и окислительное")
Хемолитотрофы. Организмы, окисляющие серу (Thiobacillus). Окисляют серу, сероводород и тиосульфат, используя субстратное и окислительное фосфорилирование. Образуют аденозин-5 фосфосульфат. Бактерии Sulfolobus способны аэробно окислять серу, в анаэробных условиях переходить к анаэробному дыханию.


Лекция 4. Энергетический обмен прокариот.ppt