эндомембраны грибной клетки Лекция 5 2012.ppt
- Количество слайдов: 64



Эндомембранная система грибов

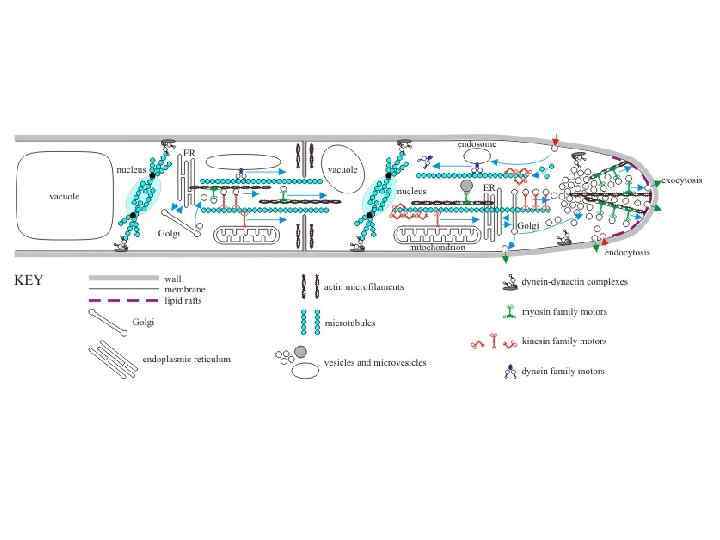
Эукариотическая мембранная система может быть определена как комплекс органелл, осуществляющих эндоцитоз и секреторные пути (экзоцитоз) и включающий: • эндоплазматический ретикулум (ЭР), • аппарат Гольджи, • эндосомы, мультивезикулярные тела, лизосомы, вакуоли, • плазматическую мембрану • и транспортные интермедиаты, такие как везикулы и микровезикулы, в том числе и апикальное тельце. Поляризованный рост обеспечивается органеллами эндомембранной системы
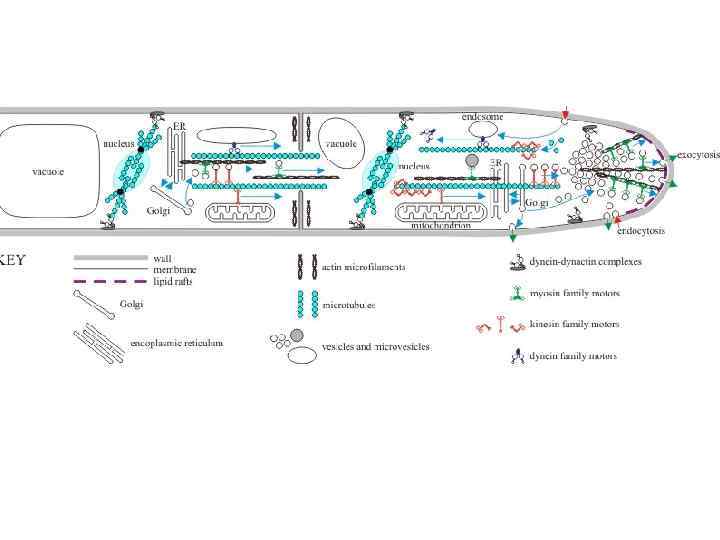

• Одним из достижений в эволюции от прокариот к эукариотам является развитие клеточной компартментализации, возможной благодаря размножению внутренних мембран. Это усовершенствование внутренних мембран позволило перевести многие биохимические процессы внутрь клетки, те что протекали на поверхности клетки у прокариот. Этот поворот позволил развитию больших клеток с безмерно развитым пространством – средний размер эукариотической клетки в 102 -103 раз больше объема прокариот.

• Эндомембранной системе грибов свойственные специфические черты организации, отличающие ее от других высших эукариот. Например, аппарат Гольджи не имеет стопки цистерн и не разделяется во время митоза. Также нет доказательства существования клатрин-одетых везикул, векторы которых отвечают за эндоцитоз и транспорт на транс-Гольджи конце.
, аппарат Гольджи и везикулы принимают участие в")
Внуриклеточные мембраны клеток грибов Эндоплазматический ретикулум (ЭР), аппарат Гольджи и везикулы принимают участие в секреторных путях (экзоцитоз). В эндоцитозе приниают участие: • Эндосомы • Вакуоли и лизосомы • Мультивезикулярные тела • Везикулы это относительно маленькие внутриклеточные органоиды, которые запасают или транспортируют питательные вещества. Они могут возникать из разных клеточных мембран (ядерной мембраны, мембран ЭПР, аппарата Гольджи, ЦПМ). Филасомы – микровезикулы с фибриллярным актиновым содержимым (встречаются в районе аппрессория, септ).
 транспортная сеть в клетке, которая сортирует молекулы белков и липидов, предназначенные")
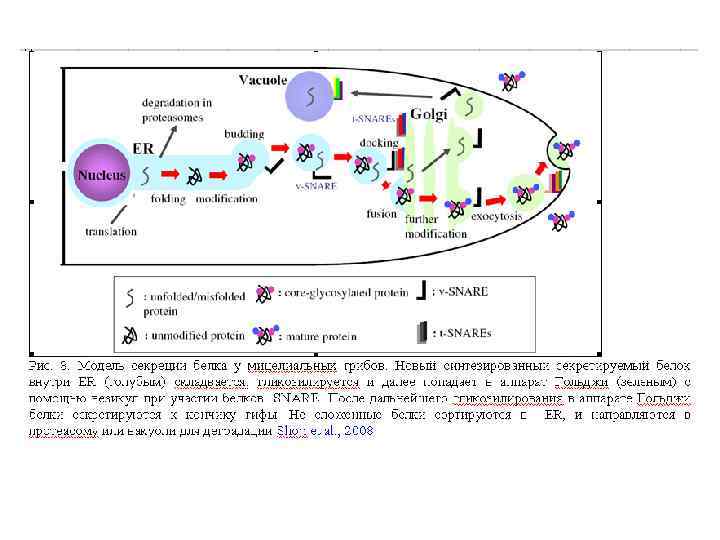
Эндоплазматический ретикулум (ЭР) транспортная сеть в клетке, которая сортирует молекулы белков и липидов, предназначенные для специфических модификаций. ЭПР это система ограниченных мембраной канальцев и мешочков (названных цистернами), связанных с двухслойной ядерной мембраной, который делает, обрабатывает и транспортируют химические компоненты для использования по клетке, и осуществляет связь между ядром и цитоплазмой. Сеть ЭР (a) Конфокальная микроскопия для выявления ЭР у гифы Aspergillus oryzae и б) Saccharomyces cerevisiae (неветвящиеся трубочки). Наблюдается экспрессия гибридного белка Bip. A-GFP (маркер ЭР с GFP). Масштабный отрезок 5 мкм. (Maruyama and Kitamoto, 2007).

Функции ЭР ЭР выполняет особую роль в перемещении липидов и белков, так как место их биосинтеза находится в цитоплазме около всех клеточных мембран, число трансмембранных белков-переносчиков ничтожно по сравнению с разнообразием этих молекул. Белки, предназначенные для модификации в аппарате Гольджи и вакуолях, первоначально котрансляционно освобождаются в люмен ЭР. Дополнительно, люмен ЭР служит для запасания ионов кальция с последующим освобождением в цитозоль при индукции через соответствующие сигналы.

Подвижность мембран ЭР Было выявлено на примере Ustilago maydis, что для подвижности ЭР необходимы цитоплазматический динеин и микротрубочки.

Распределение ЭР в клетках мицелия Aspergillus oryseae

Эндоплазматический ретикулум у U. maydis.

Реорганизация ЭР ЭР может претерпевать структурную реорганизацию. Так, перед прорастанием спор грибов, сопровождающимся их гидратацией, объем ЭР сильно увеличен. У фитопатогенного гриба Uromyces appendiculatus ЭР претерпевает структурную реорганизацию в процессе патогенеза. Так, в гаустории этого биотрофного гриба в ответ на сигналы хозяина происходит формирование трубчато-везикулярной сети с преимущественным развитием трубочек.

Реорганизация ЭР происходит и при формировании атрактосом и симплехосом у хищных грибов. Атрактосомы формируются из цистерны ЭР; цистерна закручивается в пределах ограниченного пространства и формирует глобулярный компартмент, атрактосому. В течение этого процесса обычно по периферии атрактосомы добавляется электронно-плотный материал.

Симплехосомы и атрактосомы у хищных грибов
 Agaricus bisporus")
ЭР (шероховатый и гладкий) Agaricus bisporus
 вокруг септы шампиньона")
Шапочка из ЭР (гладкого) вокруг септы шампиньона
 — возможно наименее понятная органелла, которая составляет грибную эндомембрнную систему.")
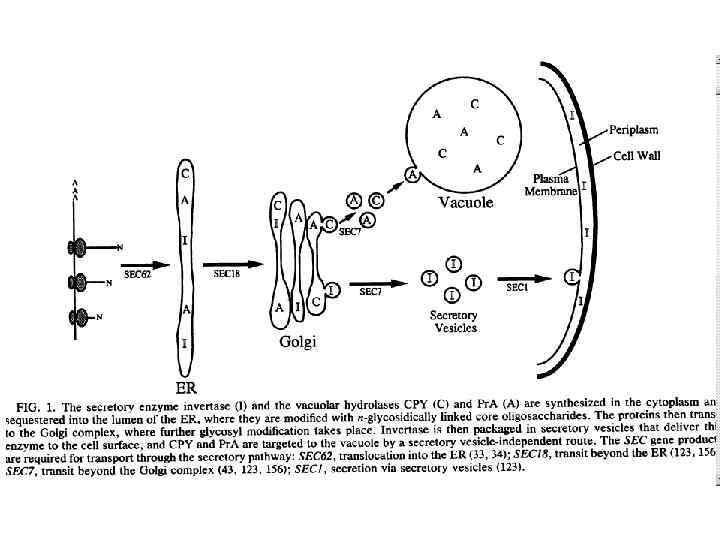
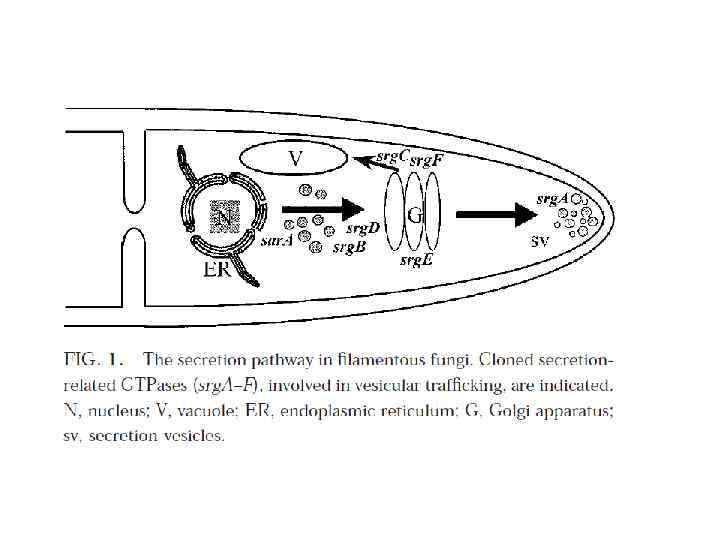
Аппарат Гольджи (комплекс Гольджи) — возможно наименее понятная органелла, которая составляет грибную эндомембрнную систему. Это место, где продукты секреции - белки (плазматической мембраны) упаковываются в транспортные вакуолярные компартменты. Гольджи может также быть конечной точкой локализации материала, который входит в эндоцитозный путь. Структура аппарата Гольджи у грибов сильно отличается от высших растений и животных. Самое большое различие – отсутствие стопок цистерн или диктиосом у настоящих грибов. Так как у грибов нет типичных диктиосом (за исключением дрожжей), выбор терминологии для родственных Гольджи структур находится в процессе. У грибов термин аппарат Гольджи описывает общую сумму всех цистерн в клетке. У большинства грибов он имеет атипичное строение и представлен отдельными цистернами или группой цистерн, рассеянных в цитоплазме и не связанных с зоной локализации ядра. Цистерны Гольджи – эквивалент Гольджи – индивидуальные органеллы, состоящие из продырявленной пластины с трубчатыми расширениями на конце. Ширина трубочек цистерн примерно одинакова, но может варьировать у отдельных клеток. В растительных клетках и клетках грибоподобных протистов (Oomycetes) обнаруживается ряд отдельных стопок (диктиосом), в животных клетках часто содержится одна большая или несколько соединённых трубками стопок. Аппарат Гольджи представляет собой продолжение ЭР.
 – это индивидуальные органеллы, состоящие из")
«Цистерна Гольджи» = эквивалент Гольджи (Howard 1981) – это индивидуальные органеллы, состоящие из продырявленной пластины с трубчатыми ответвлениями, граничащие друг с другом
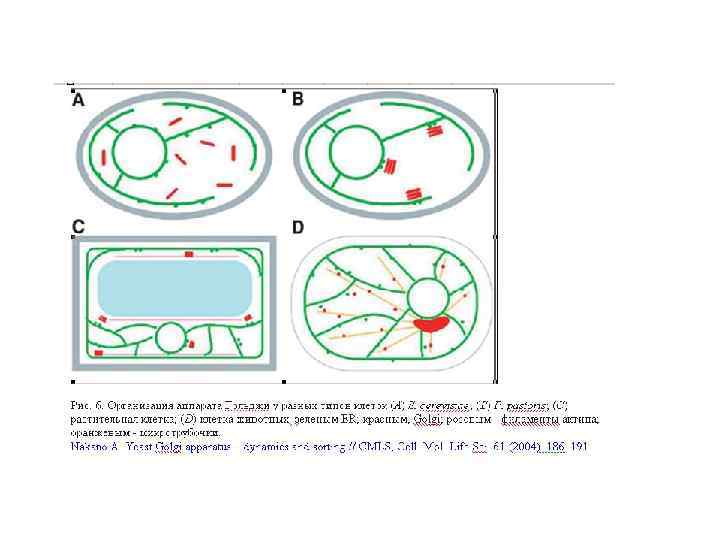

Аппарат Гольджи у дрожжей и мицелиальных грибов имеет слабые структурные различия А. Aspergillus nidulans В-Е. Pichia pastoris F. Trichoderma viride G-J. Helicobasidium mompa


Везикулы являются распространенной органеллой как в секреции, так и в транспорте белков между компартментами донора и воспринимающими компартментами эндомембранной системы. Транспорт белка ассоциированный с ядрами и митохондриями происходит без участия везикул.

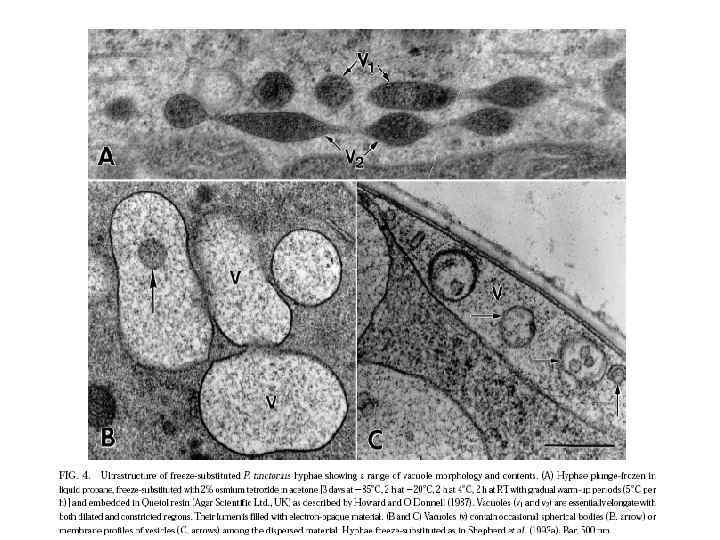
Вакуоль

Функции вакуолей Грибные вакуоли высоко подвижные плейоморфные структуры, имеющие, по-видимому, несколько функций. - они вовлечены в запасание (транспорт) катионов и метаболитов (в том числе белков), таким образом, регулируя цитоплазматический р. Н и ионный гомеостаз, - работают как литический компартмент, содержащий различные гидролитические ферменты - участвуют в детоксикации. Токсины могут либо спонтанно реагировать с SH-группами глутатиона или формировать глутатион-S-коньюганты через глутатион-S-трансферазы

Последние исследования показали, что вакуоли грибов представлены в большом диапазоне морфологии для адаптации к требованиям экологических ниш специфических видов. Они участвуют непосредственно в транспорте питательных веществ на длинные дистанции через мицелий, в регуляции растяжения и ветвления гиф, в сроках клеточного цикла и через автофагию участвуют в индукции жизненных морфогенетических процессов, таких как формирование аппрессориев и псевдогифальный рост.

Транспорт мембран включающий сортировку белка, эндоцитоз, аутофагию и наследование вакуолей у почкующихся дрожжей. Показан поток вакуолярных везикул, которые формируют отдельные вакуоли с подвижностью благодаря актиновому цитоскелету Myo 2 -actin
 дрожжи: Saccharomyces cerevisiae и Candida")
Типы и распределение вакуолей (Veses et al , 2008) дрожжи: Saccharomyces cerevisiae и Candida albicans гифа C. albicans типичный мицелиальный аскомицет патоген растений Ustilago maydis Basidiobolus ranarum трубчатая вакуолярная система типичная для Phanerochaete, Pisolithus и многих других грибов

а-в Ustilago maydis c-d-e Pisolythus tinctorius f аутофагическая конидия, атакующая аппресорию M. oryzae g-h Candida albicans


Вакуолярная система у Pisolithus tinctorius морфология и подвижность трубчатой вакуолярной системы у Pis. tinctorius зависит от микротрубочек.
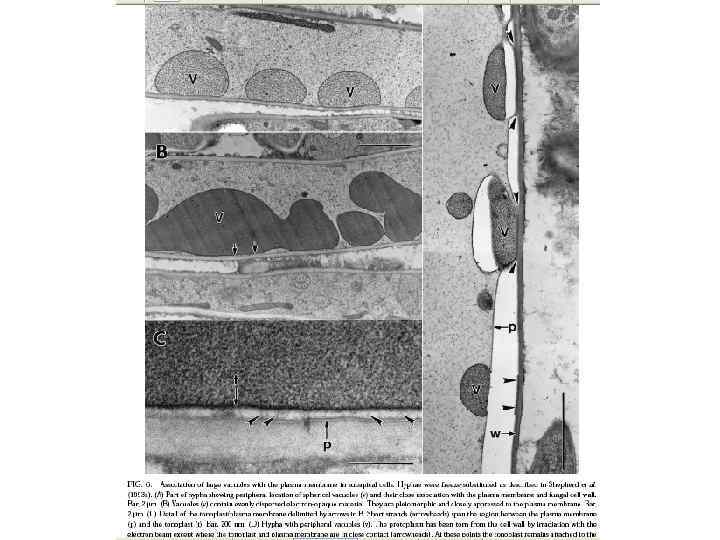

Вакуоли у арбускулярно-микоризного гриба Gigaspora margarita контактирующие трубочки

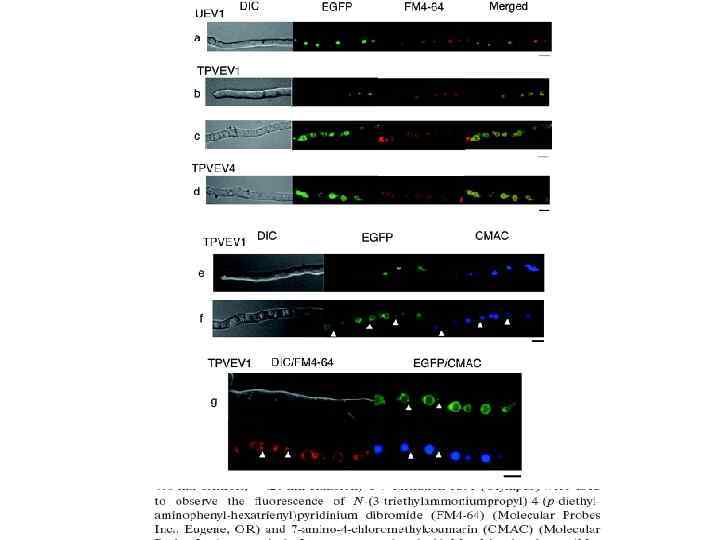

• Было показано, что вакуолярная система у Aspergillus oryzae представлена плейоморфной сетью вакуолей, включающей крупные сферические вакуоли, трубчатые вакуоли и превакуоли (поздние эндосомы). Крупные вакуоли были обнаружены в гифах, контактирующих с субстратом, что указывает на участие их в процессах аутофагии. Гифы не контактирующие с субстратом имели прерывистую систему трубчатых вакуолей, образующую ретикулум подобную сеть.

Вакуоли в прорастающих конидиях Aspergillus orizae
 и субстратном (f-i) мицелии")
Вакуоли в воздушном (а-е) и субстратном (f-i) мицелии


Phanerochaeta velutina

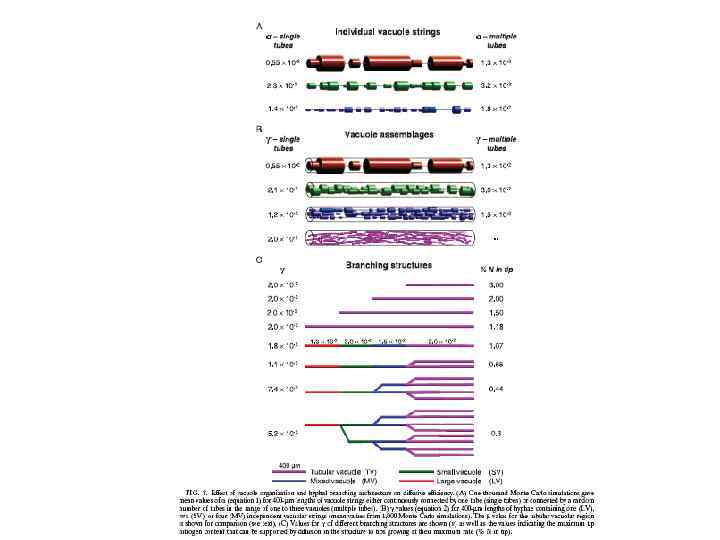

Скорость передвижения фрагментов вакуолей в гифе Neurospora crassa соответствует митохондриям 1, 8± 0, 8 мкм в сек В слиянии и делении мембран вакуолей дрожжей вовлечен динамин (дрожжи) и кинезин (Ustilago maydis) Кинезиновый и динеиновый моторы участвуют в движении вакуолей и локализованы на +концах микротрубочек Морфология и подвижность трубчатой вакуолярной системы у Pis. tinctorius зависит от микротрубочек. А у дрожжей Sac cereviviae, движение вакуолей в развивающуюся почку основано на актин-зависимых механизмах.
у Sac. cerevisiae и")
Содержимое вакуолей и р. Н • полифосфаты • азотсодержащие аминокислоты (аргинин)у Sac. cerevisiae и N. crassa • белки, включая гидролитические ферменты • р. Н вакуолей ниже (р. Н 6, 28± 0, 10) у клеток Sac. cerevisiae в стационарной фазе роста по сравнению с клетками в логарифмической стадии роста (р. Н 6, 58± 0, 14) и по сравнению с цитоплазматическим р. Н 7, 19± 0, 05 и 6, 97± 0, 12
, -микровезикулы (филасомы), -эндосомы (ранние и поздние),")
Органеллы эндоцитозного пути: -цитоплазматическая мембрана (инвагинация и транспортеры-пермеазы), -микровезикулы (филасомы), -эндосомы (ранние и поздние), -мультивезикулярные тела и вакуоли

Инвагинации ЦПМ и филасомы Органеллы эндоццитоза у мицелиальных грибов –A, B Gilbertella persicaria и Magnaporthe grisea, соответственно –опосредован микровезикулами называемыми филасомами (стрелки), а не clathrin-coated везикулами (C). Филасомы также обнаружены у дрожжей Pichia. Иммуноцитохимия демонстрирует что покров филасом содержит актин у M. grisea (D) и P. pastoris (E). Дополнительно к филасомам плазматическая мембрана дрожжей Pichia имеет инвагинации (F) которые, поверхностно напоминают caveoli клеток животных. G Продольные срезы относительно плазматической мембраны предполагают, что эти инвагинации могут быть вытянутыми и менее четко трубчатыми. All specimens were prepared by freeze substitution. cw Cell wall. Bars 250 nm, except A (500 nm) and B, F (100 nm) Bourett et al. , 2007

Ранние эндосомы у мицелиальных грибов были впервые описаны у диморфного гриба Ustilago maydis. Совсем недавно сходные структуры были обнаружены у A. nidulans и A. oryzae в ходе наблюдения эндоцитозного пути. Эти везикулярные структуры (ранние эндосомы) перемещаются на длинную дистанцию (50 мкм) вдоль оси гифы со скоростью 5 мкм/с зависимым от микротрубочек образом в отличие от дрожжей. Белки плазматической мембраны и внеклеточные молекулы упаковываются в везикулы путем инвагинации мембраны и последовательно транспортируются к ранним эндосомам.

Dynein, dynactin, Lis 1 on moving early endosomes in Ustilago maydis

Ранние эндосомы у Aspergillus nidulans

Эндоцитозный путь у дрожжей Positively charged Nanogold can be used as an endocytic marker. Wild-type and end 3 mutant strains were grown overnight, converted to spheroplasts, and incubated under various conditions The samples were fixed and embedded and thin sections were cut. (A) Sections from wild-type cells incubated with positively charged Nanogold were enhanced with HQ Silver. (B) Sections from end 3 mutant cells incubated with positively charged Nanogold and enhanced.

Мультивезикулярные тела, вероятно, промежуточные компартменты между ранними и поздними эндосомами и могут быть обильными в гифе (стрелки) как это проиллюстрировано у Gilbertella persicaria. Они разбросаны с микротрубочками и везикулами, которые совершают движение в двух направлениях. Масштабный отрезок 500 нм Bourett et al. , 2007

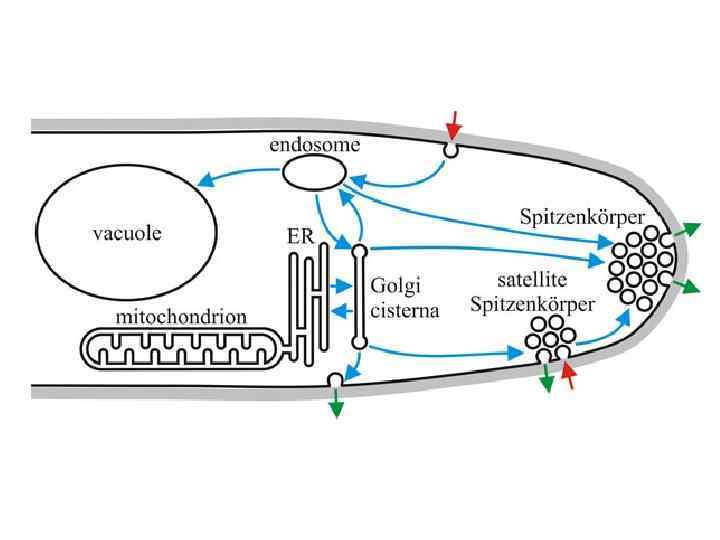
Эндоцитоз у аспергилла занимает 40 минут Схема пространственного распределения органелл и везикулярного транспорта у мицелиальных грибов. Окрашенные символы представляют органеллы и клеточные компоненты: ядро - красным, ER - светло голубым, аппарат Гольджи - оранжевым, клеточная стенка – коричневым, ранняя эндосома – фиолетовым, поздняя эндосома – серым, вакуоль – зеленым. ЭПР в виде кольца окружает ядро или в виде трубчатых структур которые показывают высокий градиент в кончике, но некоторый ЭПР также присутствует около плазматической мембраны и септы. Аппарат Гольджи имеет тенденцию к распространению в апикальных районах, а развитые вакуоли в дистальных районах. Поздние эндосомы локализованы около вакуолей, а ранние – по всей гифе и двигаются вдоль оси гифы. Красные стрелки показывают пути секреции белка, синие – эндоцитозный путь Shoji et al. , 2008

Автофагия В природе, организмы возвращают обратно свои внутриклеточные компоненты через вакуоли/лизосомы и процесс автофагии. Термин автофагия на самом деле покрывает множество процессов: (i) не-селективная (или макро-) автофагия, которая вовлекает случайно захваченные порции цитоплазмы (цитозоль и органеллы) в вакуоль/лизосому для повторного использования (ii) селективная деградация старых/лишних органелл таких как пероксисомы (микропексофагия) и митохондрии (митофагия). (iii) специфический лизис белков в цитоплазме

1 3 аминопептидаза 2 а, б

Первый процесс имеет особое значение для клеточных организмов при истощении питательных веществ. В процессе макроаутофагии у Saccharomyces cerevisiae, двойная мембрана окружает порцию цитоплазмы. Это приводит к формированию аутофагососмы. Позднее наружный слой аутофагосомы сливается с мембраной вакуоли. В заключении одномембранная структура в вакуолярном люмене, сейчас упоминают, как аутофагическое тело деградирует вакуолярными гидролазами для пополнения питательных запасов.

Второй - родственный автофагии процесс вовлекает селективную деградацию пероксисом и митохондрий

Третий процесс также селективный, основополагающий процесс для сортировки основных резидентных гидролаз к вакуоли. Этот процесс обозначен как cytoplasm to vacuole targeting (Cvt) путь и не так давно был описан для S. cerevisiae. В процессе Cvt пути, формируются ферменты: аминопептидаза I (Ape 1) и a‑маннозидазу (Ams 1), которые встраиваются в двухмембранную структуру Cvt везикулу. Так как в макроаутофагии, двухмембранная везикула сливается с вакуолью и приводит к формированию одномембранного Cvt тела в вакуолярном люмене. После лизиса Cvt гидролазами вакуолей зимогены созревают и активируются.

Идентификацию генов вовлеченных в макроавтофагию, Cvt путь, макро- и микропексофагия проводят на видах дрожжей. Эти исследования привели к изоляции и функциональной характеристике 29 ATG генов.

Таким образом, из изложенного выше материала хорошо видно, что эндомембранная система у грибов выполняет чрезвычайно важные функции (эндо-, экзоцитоз), имеет ряд специфических черт, но пока остается во многом слабо изученной системой у грибов как на структурном, так и на молекулярном уровнях.
эндомембраны грибной клетки Лекция 5 2012.ppt