Лекция 6-2017 эндомембраны грибной клетки.ppt
- Количество слайдов: 84



Эндомембранная система грибов
 эукариот система представляет собой совокупность органелл, составляющих эндо- и экзосекреторные пути")
Эндомембранная система (ЭС) эукариот система представляет собой совокупность органелл, составляющих эндо- и экзосекреторные пути и включающих: эндоплазматический ретикулум (ЭР), аппарат Гольджи, эндосомы, мультивезикулярные тельца, вакуоли-лизосомы, транспортные вакуоли, плазматическую мембрану и транспортные интермедиаты - везикулы, а также экзосомы Поляризованный рост у грибов обеспечивается органеллами эндомембранной системы
 грибов. Стрелками обозначены")
Объединенная схема, отражающая предположительные связи между различными структурами эндомембранной системы (ЭС) грибов. Стрелками обозначены возможные переходы органелл одна в другую (созревание), слияние органелл или везикулярный транспорт от одной эндомембранной структуры к другой. Пунктирные стрелки – гипотетические переходы. Светлые стрелки (ЭНП) соответствуют эндоцитозному пути и внутреннему транспорту. Темные стрелки (ЭКП) – экзоцитозный путь. Обозначения на рисунке: АГ – актиновые глобулы, входящие в субапикальное кольцо, АТ – апикальное тельце, АФ – автофагосома, В – вакуоль, ВЗ – различные везикулы (первичные эндоцитозные, экзоцитозные, отвечающие за внутренний транспорт), ГЖ – цистерны Гольджи, МВТ – мультивезикулярное тело, ПЭ – поздняя эндосома, РЭ – ранние эндосомы, ЭР – эндоплазматический ретикулум, ЭС – экзосома, Я – ядро. Изображена утрированная схема апекса мицелиального гриба с простой септой (мембранный аппарат септы не отражен)

• Одним из достижений в эволюции от прокариот к эукариотам является развитие клеточной компартментализации, возможной благодаря размножению внутренних мембран. Это усовершенствование внутренних мембран позволило перевести многие биохимические процессы внутрь клетки. Этот поворот позволил развитию больших клеток с безмерно развитым пространством – средний размер эукариотической клетки в 102 -103 раз больше объема прокариот. Компартменты – обособленные области в клетке, как правило, изолированные друг от друга внутриклеточными мембранами.

• Эндомембранной системе грибов свойственны специфические черты организации, отличающие ее от других высших эукариот. Например, аппарат Гольджи не имеет стопки цистерн и не разделяется во время митоза. Также нет доказательства существования клатринодетых везикул у мицелиальных грибов, векторы которых отвечают за эндоцитоз и транспорт на транс-Гольджи конце.
, аппарат")
Внуриклеточные мембраны клеток грибов, принимающие участие в экзо- и эндоцитозе Эндоплазматический ретикулум (ЭР), аппарат Гольджи, везикулы, экзосомы плазмалемма принимают участие в секреторных путях (экзоцитоз). В эндоцитозе приниают участие: • Плазмалемма (ЦПМ) • Везикулы • Эндосомы • Мультивезикулярные тела • Вакуоли
, Аппарат Гольджи, Везикулы, Экзосомы, Плазмалемма")
Органеллы экзоцитоза у грибов Эндоплазматический ретикулум (ЭР), Аппарат Гольджи, Везикулы, Экзосомы, Плазмалемма
 грибов. Стрелками обозначены")
Объединенная схема, отражающая предположительные связи между различными структурами эндомембранной системы (ЭС) грибов. Стрелками обозначены возможные переходы органелл одна в другую (созревание), слияние органелл или везикулярный транспорт от одной эндомембранной структуры к другой. Пунктирные стрелки – гипотетические переходы. Светлые стрелки (ЭНП) соответствуют эндоцитозному пути и внутреннему транспорту. Темные стрелки (ЭКП) – экзоцитозный путь. Обозначения на рисунке: АГ – актиновые глобулы, входящие в субапикальное кольцо, АТ – апикальное тельце, АФ – автофагосома, В – вакуоль, ВЗ – различные везикулы (первичные эндоцитозные, экзоцитозные, отвечающие за внутренний транспорт), ГЖ – цистерны Гольджи, МВТ – мультивезикулярное тело, ПЭ – поздняя эндосома, РЭ – ранние эндосомы, ЭР – эндоплазматический ретикулум, ЭС – экзосома, Я – ядро. Изображена утрированная схема апекса мицелиального гриба с простой септой (мембранный аппарат септы не отражен)
 ЭР - транспортная система ограниченных мембраной канальцев и мешочков (названных цистернами),")
Эндоплазматический ретикулум (ЭР) ЭР - транспортная система ограниченных мембраной канальцев и мешочков (названных цистернами), . связанных с двухслойной ядерной мембраной, которая образует, обрабатывает и транспортирует молекулы белков и липидов для использования по клетке и осуществляет связь между ядром и цитоплазмой Сеть ЭР (a) Конфокальная микроскопия для выявления ЭР у гифы Aspergillus oryzae и б) Saccharomyces cerevisiae (неветвящиеся трубочки). Наблюдается экспрессия гибридного белка Bip. A-GFP (маркер ЭР с GFP). Масштабный отрезок 5 мкм. (Maruyama and Kitamoto, 2007).
 ЭР выполняет особую роль в перемещении липидов и белков, так как")
Эндоплазматический ретикулум (ЭР) ЭР выполняет особую роль в перемещении липидов и белков, так как место их биосинтеза находится в цитоплазме около всех клеточных мембран, число трансмембранных белков-переносчиков ничтожно по сравнению с разнообразием этих молекул. Белки, предназначенные для модификации в аппарате Гольджи и вакуолях, первоначально котрансляционно освобождаются в люмен ЭР. Дополнительно, люмен ЭР служит для запасания ионов кальция с последующим освобождением в цитозоль.

Подвижность мембран ЭР Было выявлено на примере Ustilago maydis, что для подвижности ЭР необходимы цитоплазматический динеин и микротрубочки.

В 2013 году Нобелевскую премию по физиологии и медицине вручили Джеймсу Ротману, Рэнди Шекману и Томасу Зюдофу — «за открытие системы везикулярного транспорта- основной транспортной системы в клетках эукариот» .

Гены везикулярного транспорта: при чем тут дрожжи? Рэнди Шекман, изучавший биохимию под началом Артура Корнберга (лауреата Нобелевской премии 1959 года), решил заняться механизмом молекулярного транспорта с позиций генетики и выбрал в качестве модельного объекта пекарские дрожжи (Saccharomyces cerevisiae). Шекман собрался выявить гены, которые участвуют в процессе везикулярного транспорта, с помощью мутантных форм дрожжей и анализа накапливающихся внутри клетки секреторных ферментов. Первые два открытых гена получили названия sec 1 и sec 2, а более детальное сканирование выявило в сумме 23 гена. Эти белки разделились на три группы, в зависимости от того, блокировался ли везикулярный транспорт на уровне ЭР, аппарата Гольджи или секреции за пределы клеточной мембраны (рис. ). Позже была определена последовательность событий посттрансляционной модификации, которые управляют процессом секреции. Детальное генетическое и морфологическое исследование «отловило» промежуточные стадии транспорта между ЭР и аппаратом Гольджи, и особенно это было заметно на мутантах sec 17 и sec 18, в которых секреторные пузырьки
 и")
Пути секретируемого фермента инвертазы и вакуолярных гидролаз у дрожжей Секретируемый фермент инвертаза (I) и вакуолярные гидролазы CPY (С) и Pr. A (А) синтезируются в цитоплазме и переносятся в люмен ЭР, где они модифицируются гликозидной связью с олигосахаридами. Белки далее переносятся в комплекс Гольджи, где происходит модификация гликозила. Инвертаза затем упаковывается в секреторные везикулы, которые удаляют фермент к клеточной поверхности, а CPY и Pr. A транспортируются к вакуоли без участия везикул. Продукты гена SEC участвуют в транспорте через секреторный путь: SEC 62 транспортируется в ЭР, SEC 18 переносится не через ЭР, SEC 7 переносится не через комплекс Гольджи, SEC 1 секретируется через секреторные везикулы.

Везикулярный транспорт ГТФ-аз у мицелиальных грибов

Распределение ЭР в клетках мицелия Aspergillus oryseae

Эндоплазматический ретикулум у Ustilago maydis дрожжи мицелий

Реорганизация ЭР ЭР может претерпевать структурную реорганизацию. Так, перед прорастанием спор грибов, сопровождающимся их гидратацией, объем ЭР сильно увеличен. У фитопатогенного гриба Uromyces appendiculatus ЭР претерпевает структурную реорганизацию в процессе патогенеза. Так, в гаустории этого биотрофного гриба в ответ на сигналы хозяина происходит формирование трубчато-везикулярной сети с преимущественным развитием трубочек.

Реорганизация ЭР происходит и при формировании атрактосом и симплехосом у хищных грибов. Атрактосомы формируются из цистерны ЭР; цистерна закручивается в пределах ограниченного пространства и формирует округлый компартмент, атрактосому. В течение этого процесса обычно по периферии атрактосомы добавляется электронно-плотный материал.

Симплехосомы и атрактосомы у хищных грибов
 Agaricus bisporus")
ЭР (шероховатый и гладкий) Agaricus bisporus
 вокруг септы шампиньона Аппарат Гольджи")
Шапочка из ЭР (гладкого) вокруг септы шампиньона Аппарат Гольджи
 грибов. Стрелками обозначены")
Объединенная схема, отражающая предположительные связи между различными структурами эндомембранной системы (ЭС) грибов. Стрелками обозначены возможные переходы органелл одна в другую (созревание), слияние органелл или везикулярный транспорт от одной эндомембранной структуры к другой. Пунктирные стрелки – гипотетические переходы. Светлые стрелки (ЭНП) соответствуют эндоцитозному пути и внутреннему транспорту. Темные стрелки (ЭКП) – экзоцитозный путь. Обозначения на рисунке: АГ – актиновые глобулы, входящие в субапикальное кольцо, АТ – апикальное тельце, АФ – автофагосома, В – вакуоль, ВЗ – различные везикулы (первичные эндоцитозные, экзоцитозные, отвечающие за внутренний транспорт), ГЖ – цистерны Гольджи, МВТ – мультивезикулярное тело, ПЭ – поздняя эндосома, РЭ – ранние эндосомы, ЭР – эндоплазматический ретикулум, ЭС – экзосома, Я – ядро. Изображена утрированная схема апекса мицелиального гриба с простой септой (мембранный аппарат септы не отражен)
 — возможно наименее понятная органелла, которая составляет грибную эндомембранную систему.")
Аппарат Гольджи (комплекс Гольджи) — возможно наименее понятная органелла, которая составляет грибную эндомембранную систему. Аппарат Гольджи представляет собой продолжение мембран ЭР и является местом созревания и сортировки белков и липидов. Аппарат Гольджи Структура аппарата Гольджи у грибов сильно отличается от высших растений и животных. Самое большое различие – отсутствие стопок цистерн или диктиосом у настоящих грибов. Так как у грибов нет типичных диктиосом (за исключением дрожжей), выбор терминологии для родственных Гольджи структур находится в процессе. У грибов термин аппарат Гольджи описывает общую сумму всех цистерн в клетке. У большинства грибов он имеет атипичное строение и представлен отдельными цистернами или группой цистерн, рассеянных в цитоплазме и не связанных с зоной локализации ядра. Цистерны Гольджи – эквивалент Гольджи – индивидуальные органеллы, состоящие из трубочек или продырявленной пластины с трубчатыми расширениями на конце. Ширина трубочек цистерн примерно одинакова, но может варьировать у отдельных клеток. В растительных клетках и клетках грибоподобных протистов (Oomycetes) обнаруживается ряд отдельных стопок (диктиосом), в животных клетках часто содержится одна большая или несколько соединённых трубками стопок.
 – это индивидуальные органеллы, состоящие из")
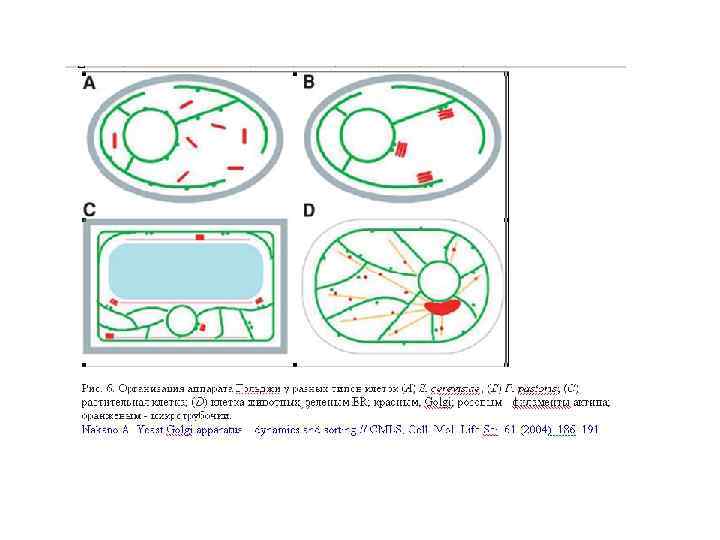
«Цистерна Гольджи» = эквивалент Гольджи (Howard 1981) – это индивидуальные органеллы, состоящие из продырявленной пластины с трубчатыми ответвлениями, граничащие друг с другом

Аппарат Гольджи у дрожжей и мицелиальных грибов имеет слабые структурные различия А. Aspergillus nidulans В-Е. Pichia pastoris F. Trichoderma viride G-J. Helicobasidium mompa

Секреторный аппарат

Везикулы являются распространенной органеллой как в секреции, так и при внутриклеточном транспорте белков между компартментами донора и воспринимающими компартментами эндомембранной системы. Транспорт белка ассоциированный с ядрами и митохондриями происходит без участия везикул.
, которые запасают")
• Везикулы это относительно маленькие внутриклеточные органоиды (от 30 -150 нм), которые запасают или транспортируют различные вещества. Они могут возникать из клеточных мембран: ядерной мембраны, мембран ЭПР, аппарата Гольджи, ЦПМ.

Плазмалемма

Экзосомы Секреторная везикула, прошедшая через плазматическую мембрану, несущая секреторный белок, материал клеточной стенки и др. Может быть локализована в периплазматическом пространстве между плазмалеммой и клеточной стенкой.

Экзосомы Agaricus bisporus

Экзосомы Cryptococcus neoformans

Связь экзоцитоза и эндоцитоза
 Везикулы и микровезикулы Эндосомы Мультивезикулярные")
Органеллы эндоцитоза у грибов • • • Плазмалемма (ЦПМ) Везикулы и микровезикулы Эндосомы Мультивезикулярные тела Вакуоли

Эндоцитоз у аспергилла занимает 40 минут Объединенная схема, отражающая предположительные связи между различными структурами эндомембранной системы (ЭС) грибов. Стрелками обозначены возможные переходы органелл одна в другую (созревание), слияние органелл или везикулярный транспорт от одной эндомембранной структуры к другой. Пунктирные стрелки – гипотетические переходы. Светлые стрелки (ЭНП) соответствуют эндоцитозному пути и внутреннему транспорту. Темные стрелки (ЭКП) – экзоцитозный путь. Обозначения на рисунке: АГ – актиновые глобулы, входящие в субапикальное кольцо, АТ – апикальное тельце, АФ – автофагосома, В – вакуоль, ВЗ – различные везикулы (первичные эндоцитозные, экзоцитозные, отвечающие за внутренний транспорт), ГЖ – цистерны Гольджи, МВТ – мультивезикулярное тело, ПЭ – поздняя эндосома, РЭ – ранние эндосомы, ЭР – эндоплазматический ретикулум, ЭС – экзосома, Я – ядро. Изображена утрированная схема апекса мицелиального гриба с простой септой (мембранный аппарат септы не отражен)

Функции эндоцитоза Эндоцитозный компонент грибов – комплекс связанных мембранных органелл, выполняющих следующие функции: поглощение биомолекул из окружающей среды (питание мицелия), удаление и деградация белков и липидов плазмалеммы, участие в сигнальных путях, удаление белков сигнальных путей, круговорот белков к эквиваленту Гольджи и от него (Lemmon, Traub, 2000).

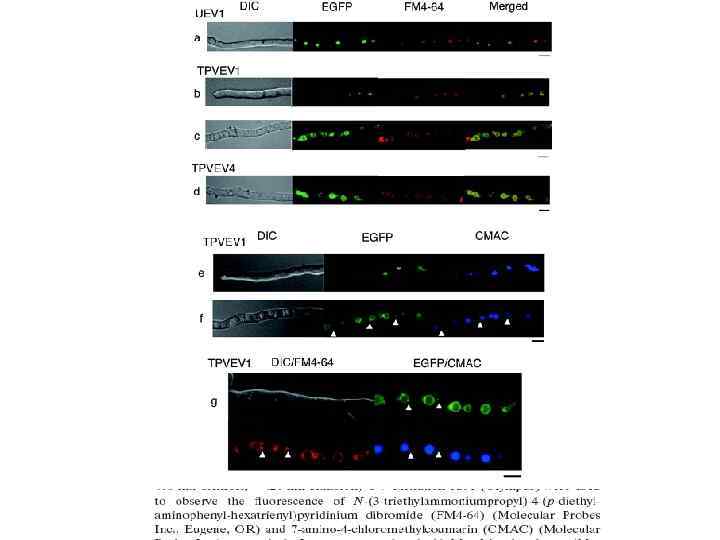
СХЕМА РАБОТЫ И ОРГАНИЗАЦИЯ МАРКЕРА ЭНДОЦИТОЗА AM 4 -64 Обозначения на схеме: КС – клеточная стенка, ЦМ – цитоплазматическая мембрана, МАВ – макровезикулы, МИВ – микровезикулы, РЭ – ранние эндосомы, ПЭ – поздние эндосомы, МВТ – мультивезикулярные тела, ВК – вакуоли.

Метод выявления эндоцитоза у R. solani с помощью флуоресцентного маркера эндоцитоза АМ 4 -64

Динеин, Динактин, белокрегулятор динеина Lis 1 в движении эндосом у Ustilago maydis
 Срезы протопластов клеток")
Эндоцитозный путь у почкующихся дрожжей Наночастицы золота – маркер эндоцитоза. (A) Срезы протопластов клеток дикого штамма, инкубированного с положительно заряженными частицами Нанозолота, обогащенного HQ серебром для выявления эндоцитоза на ТЭМ. (B) Срезы протопластов клеток мутантного штамма end 3

Ранние стадии эндоцитоза у почкующихся дрожжей
")
Пути эндоцитоза у клеток животных и грибов (Steinberg, 2014)

– Эндоцитоз и апикальный рост: рециклизация и деградация рецепторных белков AP-1/2 адапторные белки везикулярного транспорта Syn. A – синаптобревин Dnf. A и Dnf. B – флиппазы Sag. A и Sla. B факторы эндоцитоза Uap. A - uric acid-xanthine transporter Martzoukou et al. , 2017

Ранняя эндосома и комплекс белков, участвующих в перемещении по микротрубочке у U. maydis.
.")
Схема эндосомо-/лизосомальной системы у клеток животных (Huotari and Helenius, 2011).
 роль ранних эндосом у")
Схема эндоцитозного пути у растительной клетки (Contento and Bassham, 2012) роль ранних эндосом у растений выполняет транс-сеть аппарата Гольджи.

Ранние эндосомы у грибов были описаны у почкующихся дрожжей и Ustilago maydis. Совсем недавно сходные структуры были обнаружены у Aspergillus nidulans и A. oryzae в ходе наблюдения эндоцитозного пути. Эти везикулярные структуры (или ранние эндосомы) перемещаются на длинную дистанцию (50 мкм) вдоль оси гифы со скоростью 5 мкм/с зависимым от микротрубочек образом в отличие от дрожжей. Белки плазматической мембраны и внеклеточные молекулы упаковываются в везикулы путем инвагинации мембраны и последовательно транспортируются к ранним эндосомам.

Мультивезикулярные тела Название «мультивезикулярные тела» было введено еще в 50 -е гг. прошлого века для обозначения мелких внутриклеточных пузырьков, ограниченных мембраной толщиной 50 -60 ангстрем обнаруженных в яйцеклетках крыс (Sotelo & Porter, 1959) Структуры, аналогичные МВТ, найдены в мицелии и конидиях различных грибов (Buckley et al. , 1966; Calonge, 1969), в клетках водоросли Chlаmydomonas reinhardi (Sager, Palade, 1957), в тканях высших растений и животных (Walker & Bisalputra 1967).

Мвт Sclerotinia sclerotiorum

Мультивезикулярное тело Podospora anserina

Мвт Fomes fomentarius

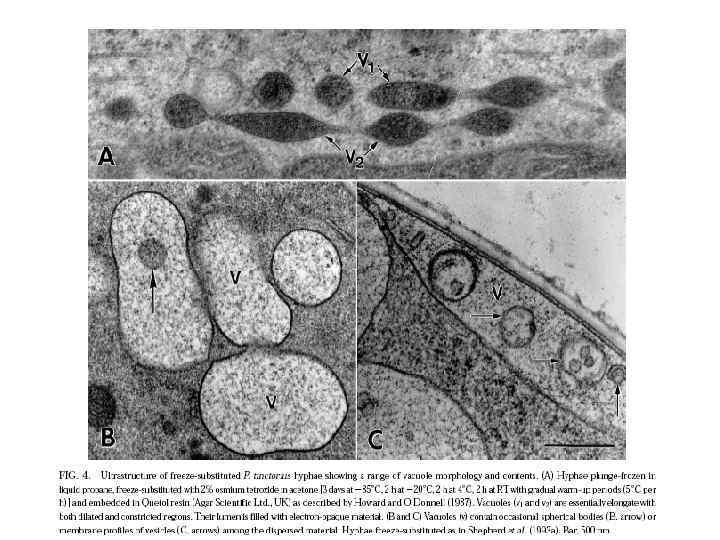
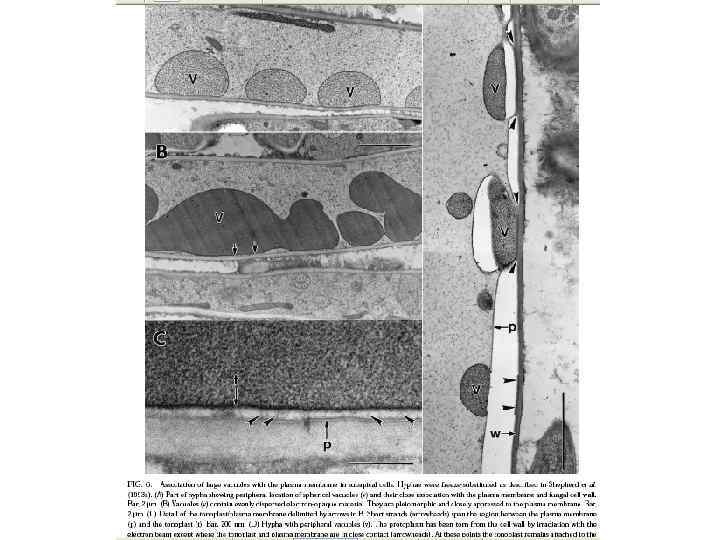
Вакуоль

Вакуоль – окруженная однослойной мембраной или тонопластом органелла клетки эукариот и некоторых прокариот, содержимое которой представляет собой водный раствор неорганических (за исключением комплекса полифосфатов с белками и неорганическими ионами) и органических молекул, включая ферменты, и выполняющая разнообразные функции (запасание, транспорт, автолиз макромолекул и клеточных компонентов и др. ). Вакуоли - это фактически крупная везикула, которая образуется путем слияния множества мелких везикул.

Последние исследования показали, что вакуоли грибов имеют разнообразную морфологию, что связано с адаптацией клеток к требованиям экологических ниш специфических видов. Они участвуют непосредственно в транспорте питательных веществ на длинные дистанции через мицелий, в регуляции роста и ветвления гиф, в сроках клеточного цикла и через автофагию участвуют в индукции морфогенетических процессов, таких как формирование аппрессориев и псевдогифальный рост.

Транспорт мембран, включающий сортировку белка, эндоцитоз, аутофагию и наследование вакуолей у почкующихся дрожжей. Показан поток вакуолярных везикул, которые формируют отдельные подвижные вакуоли благодаря актиновому цитоскелету Myo 2 -actin
 дрожжи: Saccharomyces cerevisiae и Candida")
Типы и распределение вакуолей (Veses et al , 2008) дрожжи: Saccharomyces cerevisiae и Candida albicans гифа C. albicans типичный мицелиальный аскомицет патоген растений Ustilago maydis Basidiobolus ranarum трубчатая вакуолярная система типичная для Phanerochaete, Pisolithus и многих других грибов

а-в Ustilago maydis c-d-e Pisolythus tinctorius f аутофагическая конидия, атакующая аппресорию M. oryzae g-h Candida albicans (Veses et al. , 2008)


Вакуолярная система у Pisolithus tinctorius морфология и подвижность трубчатой вакуолярной системы у Pis. tinctorius зависит от микротрубочек.

Вакуоли у арбускулярно-микоризного гриба Gigaspora margarita контактирующие трубочки


• Было показано, что вакуолярная система у Aspergillus oryzae представлена плейоморфной сетью вакуолей, включающей крупные сферические вакуоли, трубчатые вакуоли и поздние эндосомы. Крупные вакуоли были обнаружены в гифах, контактирующих с субстратом, что указывает на участие их в процессах аутофагии. Гифы не контактирующие с субстратом имели прерывистую систему трубчатых вакуолей, образующую подобную ретикулуму сеть.

Вакуоли в прорастающих конидиях Aspergillus orizae
 и субстратном (f-g) мицелии")
Вакуоли в воздушном (а-е) и субстратном (f-g) мицелии


Phanerochaeta velutina
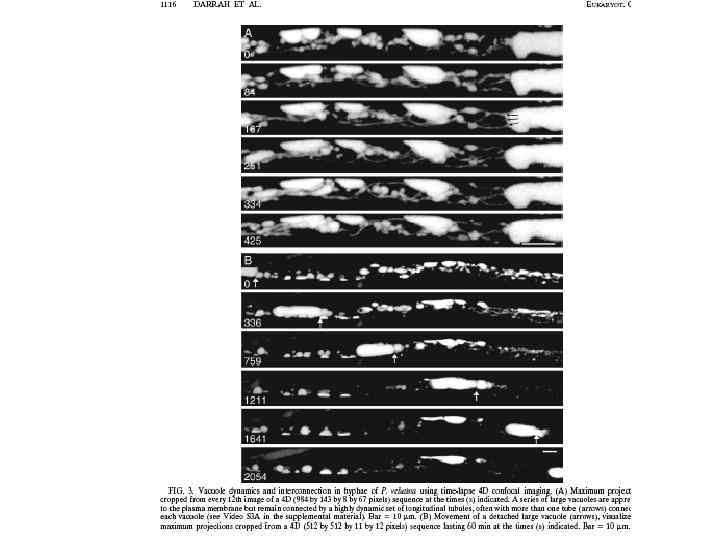
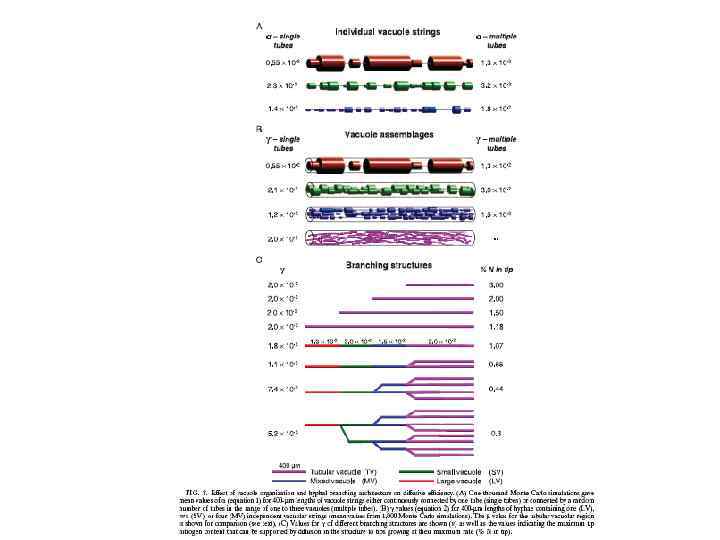

Скорость передвижения фрагментов вакуолей в гифе Neurospora crassa соответствует митохондриям 1, 8± 0, 8 мкм в сек В слиянии и делении мембран вакуолей дрожжей вовлечен динамин (дрожжи) и кинезин (Ustilago maydis) Кинезиновый и динеиновый моторы участвуют в движении вакуолей и локализованы на +концах микротрубочек Морфология и подвижность трубчатой вакуолярной системы у Pisolitus tinctorius зависит от микротрубочек. А у дрожжей Saccharomyces cereviviae движение вакуолей в развивающуюся почку основано на актин-зависимых механизмах.
 у S. cerevisiae")
Содержимое вакуолей и р. Н • полифосфаты • азотсодержащие аминокислоты (аргинин) у S. cerevisiae и N. crassa • белки, включая гидролитические ферменты • р. Н вакуолей ниже (р. Н 6, 28± 0, 10) у клеток S. cerevisiae в стационарной фазе роста по сравнению с клетками в логарифмической стадии роста (р. Н 6, 58± 0, 14) и по сравнению с цитоплазматическим р. Н 7, 19± 0, 05 и 6, 97± 0, 12

В 2016 году Нобелевский комитет присудил премию по физиологии и медицине японскому ученому Ёсинори Осуми за открытие аутофагии и расшифровку ее молекулярного механизма.

Автофагия В природе, организмы возвращают обратно свои внутриклеточные компоненты через вакуоли/лизосомы и процесс автофагии. Термин автофагия на самом деле покрывает множество процессов: (i) Не селективная (или макро-) автофагия, которая вовлекает случайно захваченные порции цитоплазмы (цитозоль и органеллы) в вакуоль/лизосому для повторного использования (ii) селективная деградация (или микро-) старых/лишних органелл, таких как пероксисомы (микропексофагия) и митохондрии (митофагия) (iii) специфический лизис белков в цитоплазме

1 3 аминопептидаза 2 а, б

Типы автофагии у дрожжей

Не селективная автофагия имеет особое значение для клеточных организмов при истощении питательных веществ. В процессе макроаутофагии у Saccharomyces cerevisiae, двойная мембрана окружает порцию цитоплазмы. Это приводит к формированию аутофагососмы. Позднее наружный слой аутофагосомы сливается с мембраной вакуоли. В заключение одномембранная структура в вакуолярном люмене (аутофагическое тело) деградирует вакуолярными гидролазами для пополнения питательных запасов. Селективная автофагия - родственный автофагии процесс вовлекает селективную деградацию органелл (пероксисом и митохондрий).

Специфический лизис белков - основополагающий процесс для сортировки основных резидентных гидролаз к вакуоли. Этот процесс обозначен на схеме как cytoplasm to vacuole targeting (Cvt) путь и не так давно был описан для S. cerevisiae. В процессе Cvt пути формируются ферменты: аминопептидаза I (Ape 1) и a‑маннозидазу (Ams 1), которые встраиваются в двухмембранную структуру Cvt везикулу. Как и в макроаутофагии, двухмембранная везикула сливается с вакуолью и приводит к формированию одномембранного Cvt тела в вакуолярном люмене. После лизиса Cvt гидролазами вакуолей зимогены созревают и активируются.
")
Включение Мвт в вакуоль Podospora anserina (макроавтофагия? )

Аутофагическое тело и лизосома Fomitopsis pinicola

Таким образом, из изложенного выше материала хорошо видно, что эндомембранная система у грибов выполняет чрезвычайно важные функции (эндо-, экзоцитоз), имеет ряд специфических черт, но пока остается во многом слабо изученной системой как на структурном, так и на молекулярном уровнях.
Лекция 6-2017 эндомембраны грибной клетки.ppt