6be6e4d5ee1c64c33f15dcc5054149f9.ppt
- Количество слайдов: 42



Эмбриональное кроветворение

Линии мышей, несущих мутации, при которых наблюдается снижение численности популяции и нарушение миграции кроветворных клеток : W – мутации в локусе W Sl (Steel) – мутации в локусе Sl # локус W несет ген рецептора тирозинкиназы (c-Kit) # локус Sl кодирует его лиганд (Kit-ligand или Steel factor – SLF). Мыши из коллекций Jackson’s Laboratories (США), крупнейшего центра, хранящего генетические линии лабораторных животных: Рис. 1: у мыши окрашены только радужка глаз и уши. Рис. 2: меланоциты отсутствуют на некоторых участках тела мышей. Высокая активность тирозинкиназы необходима для нормального процесса размножения и миграции популяций, клеток, мигрирующих в эмбриональный период развития (ППК, клетки нервного гребня, кроветворные клетки).

Экспрессия различных типов гемоглобинов в онтогенезе человека

Газообмен между гемоглобинами матери и плода

Эмбрион мыши, 15 сут. кроветворение

Происхождение кроветворных клеток: схема на конец 20 в.
:")
Эмбрион мыши, 10, 5 сут. Эмбрион мыши, 11, 5 сут. Аorta-gonad-mesonephros region (AGM):

Diagram of the ontogeny of hematopoietic stem cells. Different letters show distinct hypotheses. Dotted arrows represent alternative migration pathways Int J Dev Biol. 2009; 53(8 -10): 1529 -40.

Diagram of the ontogeny of mesenchymal stem cells. Different arrows colors and letters show distinct hypotheses
: • subaortic patches (SAPs), located below the aortic")
Происхождение эмбриональных HSC ? Аorta–gonads–mesonephros (AGM): • subaortic patches (SAPs), located below the aortic floor • hematopoietic intraaortic clusters (HIACs) SAP появляются раньше, чем HIAC (8. 5– 9 vs 10 dpc) 10 dpc: 80 -100 HSC (C) Модель появления и миграции предшественников кроветворных клеток: HSCs (оранжевые) выявляются в области SAP (1), мигрируют по направлению к аорте (2), затем HSC проникают через дно аорты и участвуют в формировании кластеров HIAC (3), а затем выходят в кровоток. Toward a model for emergence and migration of embryonic HSCs. (A) Localization of hemogenicrelated structures, SAPs (purple), and HIAC (pink) within a 10. 5 -dpc embryo. (B) Relative distribution of multipotent precursors (orange), SAPs (purple), and HIACs (pink) in 8. 5 - to 12 -dpc embryos. Whereas GATA-3 SAPs appear below the aortic floor as soon as the first multipotent precursor is generated, HIACs are present only at the AGM stage, when the number of precursors generated reaches a peak. Bertrand YJ et al. , Proc Natl Acad Sci U S A. 2005 102(1): 134 -9

Происхождение эмбриональных HSC ? Проточная цитометрия и клеточный сортинг The combinatorial approach undertaken here allowed us to distinguish between ECs and hematopoietic cells, by GATA-3, CD 41, and Flk 1 differential expression. The 3 D examination of SAP anatomy also distinguishes SAP cells from ECs because the CD 31 cell clusters located below the aortic floor do not harbor a lumen and do not connect with the vascular network Bertrand YJ et al. , Proc Natl Acad Sci U S A. 2005 102(1): 134 -9
 region Ангиобласт")
Гемангиобласт: • стенка желточного мешка • область P-Sp/AGM Рara-aortic splanchnopleura/aorta-genital ridges-mesonephros (P-Sp/AGM) region Ангиобласт HSC Гемангиобласт?

Дифференцировка гемангиобластов в стенке желточного мешка

Метод клонального анализа для исследования происхождения гемангиобластов Адгезивные клетки Неадгезивные клетки Analysis of embryos carrying complementary DNA of the green fluorescent protein targeted to the brachyury locus demonstrates that the haemangioblast is a subpopulation of mesoderm that co-expresses brachyury (also known as T). GFP–Bry embryos at E 7. 5 were dissected into the yolk sac (YS), anterior (Ant), lateral (L), posterior primitive streak (PPS) and distal primitive streak (DPS) and cultured in haemangioblast conditions (numbers of haemangioblast-derived colonies from fragments) CD 31 (эндотелиальный маркер) –зеленый Гладкомышечный актин - красный A model of haemangioblast development in the mouse embryo. The haemangioblast is a brachyury+ cell that arises in the primitive streak and migrates onto the yolk sac where it differentiates into haematopoietic (H), endothelial (E) and vascular smooth muscle (VSM) progenitor cells. NATURE |VOL 432 | 2 DECEMBER 2004 |www. nature. com/nature

Методы in vitro дифференцировки ЭСК для исследования ранней спецификации клеточных линий Notch signaling respecifies the hemangioblast to a cardiac fate To efficiently generate cardiomyocytes from embryonic stem (ES) cells in culture it is essential to identify key regulators of the cardiac lineage and to develop methods to control them. Using a tet-inducible mouse ES cell line to enforce expression of a constitutively activated form of the Notch 4 receptor, we show that signaling through the Notch pathway can efficiently respecify hemangioblasts to a cardiac fate, resulting in the generation of populations consisting of more than 60% cardiomyocytes. Notch signaling can initiate cardiaclineage specification in part through the coordinated activation of BMP signaling and the inhibition of the bcatenin dependent Wnt pathway. The cardiogenic effects of Notch signaling were demonstrated in the normal progression of cardiac mesoderm (Bry-GFP+/Flk-1–) to cardiomyocytes and in the dramatic respecification of hemangioblasts to cardiovascular progenitors. These progenitors appear to be similar to the cardiovascular progenitors (cardiovascular colony-forming cell; CV-CFC) Chen et al VOLUME 26 NUMBER 10 OCTOBER 2008 NATURE BIOTECHNOLOGY

Производные мезодермы: формирование урогенитальнной системы

Нефротом: смена 3 поколений почек в эмбриогенезе

Формирование метанефроса: взаимодействие выроста протока мезонефроса и нефрогенной бластемы
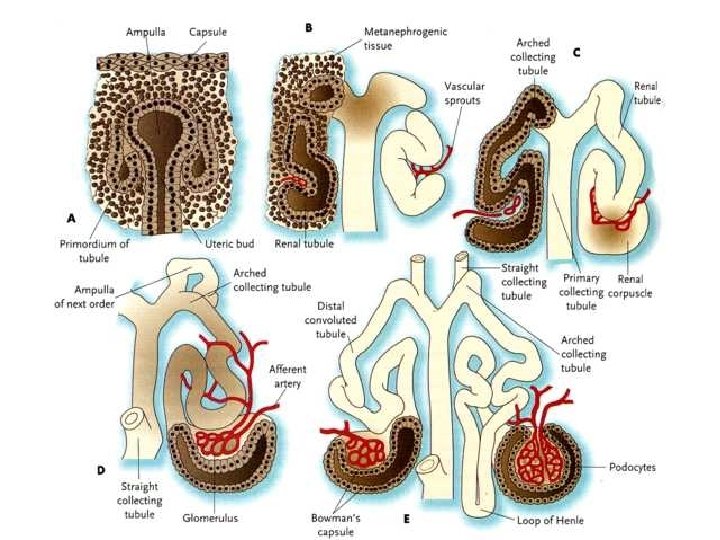
 и ламинин (формирование базальной мембраны). Начало")
Эпителизация почечных Канальцев: Клетки экспрессируют кадгерины (адгезия клеток) и ламинин (формирование базальной мембраны). Начало апикально-базальной поляризации клеток Формирование плотных контактов

Dors Vent мезонефрос метанефрос Эмбрион свиньи, 1 мес.
")
Положение метанефроса плода человека (поперечный срез)

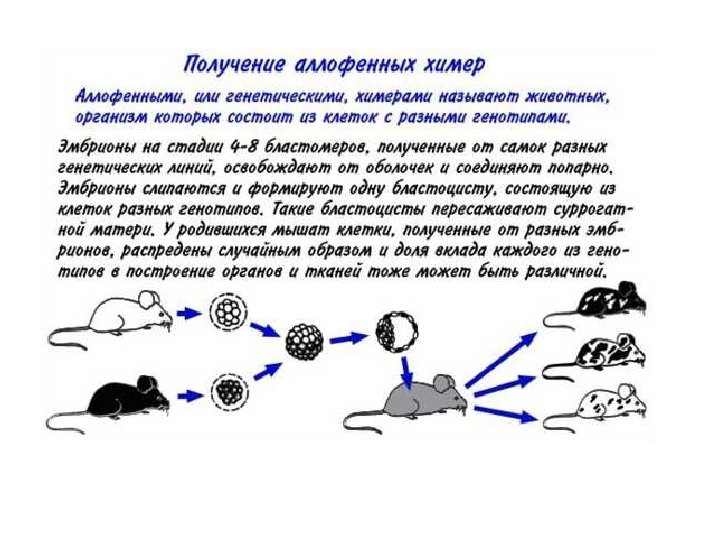
1 4 5 2 3 самец самка 6
 мыши, окраска на щелочную фосфатазу")
Первичные половые клетки (ППК) мыши, окраска на щелочную фосфатазу
 Красным заштрихованы участки, с")
Четыре гипотезы происхождения ППК (по Дыбану А. П. , 1988) Красным заштрихованы участки, с которыми связывают происхождение ППК. 1. Гипотеза половой плазмы: Цитоплазматические детерминанты, ответственные за формирование ППК, образуются во время оогенеза. 2. Сегрегационная гипотеза: Половые детерминанты образуются в цитоплазме зиготы после оплодотворения. Как и в первом случае, эти цитоплазматические факторы попадают при дроблении только в некоторые бластомеры, которые и являются предшественниками ППК. 3. Гипотеза стволовых клеток: ППК обособляются во внутренней клеточной массе бластоцисты или в эпибласте до начала гаструляции и формирования зародышевых листков. 4. Гипотеза зародышевых листков: ППК происходят от одного из трех зародышевых листков (эктодермы, энтодермы или мезодермы)

Выделение линии половых клеток у птиц: CVG – маркер клеток половаго ряда, гомолог гена Vasa N. Tsunekawa and others, 2000

Выделение линии половых клеток у мыши 6, 0 dpc 7, 0 dpc Diagram from Mc. Laren (1999). At 6. 0 dpc, a signal (solid arrows) coming from the extraembryonic ectoderm (blue) predisposes cells in the proximal layer of the epiblast (brown) towards a germ-line fate. This whole layer of PGC precursors moves (dashed arrow) towards the primitive streak and up into the extraembryonic region. At 7. 0 dpc, the newly formed extraembryonic mesoderm (gold) is moving across to form the exocoelomic cavity (white). Some of the PGC precursors stop migrating and constitute the cluster of cells representing the origin of the germ cell lineage. This may involve a second (unidentified) signal or signals (solid arrows). Yellow, visceral endoderm Diagram reproduced from Saitou et al. (2002), illustrating the fragilis expression pattern during gastrulation. (a) Lateral view of an early bud stage embryo with expression of fragilis. Anterior (A) is to the left and posterior (P) to the right. Strong expression was observed specifically at the base of the incipient allantois, the location of nascent PGCs. (b) fragilis expression from mid bud (farthest left) to early head fold (farthest right) stages, as viewed from the lateral side. fragilis expression is strong in the centre (arrowheads)—the site of the founder PGC cluster. fragilis expression fades at the early head fold stage (extreme right) when PGCs commence migration. Arrows indicate developing allantois where fragilis expression is weak or absent. (c) Lateral views of pre-streak-stage embryos (6. 25 dpc) with expression of fragilis. Intense signal was observed in proximal epiblast cells adjacent to the extraembryonic ectoderm (arrowheads). (d) fragilis expression from pre-streak (6. 0 dpc) (farthest left) to early bud (farthest right) stages. The initial domain of fragilis expression in the most proximal epiblast followed by its movement to the posterior region during gastrulation is shown. This expression pattern matches with the observations from clonal analysis for the origin, migration and segregation of the germ cell lineage. A. Mc. Laren / Developmental Biology 262 (2003) 1– 15
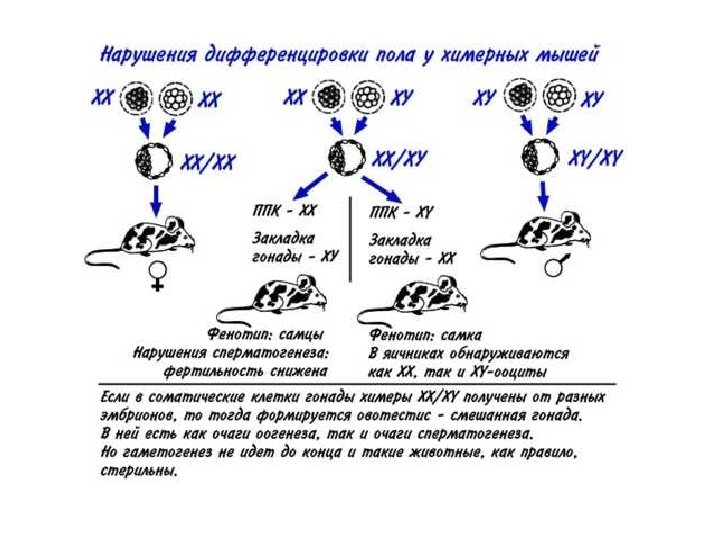

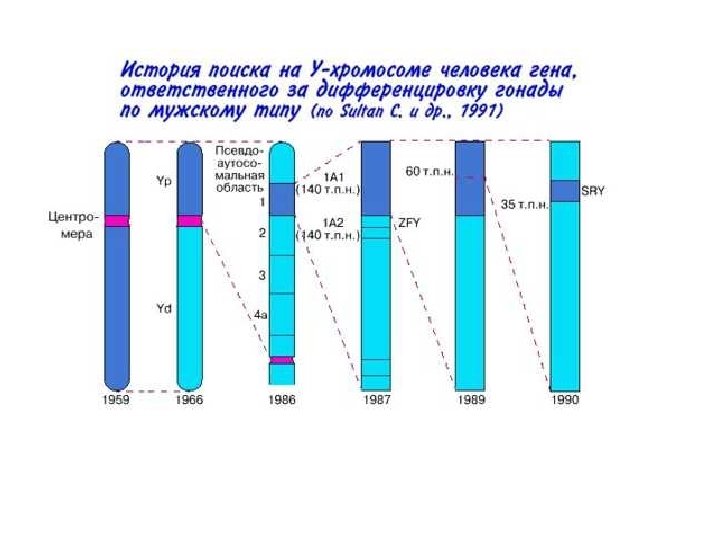
Расположение генов ответственных за дифференцировку на половых хромосомах Х-хромосома: Dax 1 AR ATRX Y-хромосома: SRY PAR: рseudo-autosomal regions; ATRX: Alpha-thalassemia/mental retardation syndrome X-linked; AZF: azoospermia factor; CSF 2 RA: Colony-stimulating factor 2 receptor alpha; DAX 1: DSS-AHC critical region X chromosome gene 1; DAZ: Deleted in azoospermia; FRA-X: Fragile X syndrome; DMD: Duchenne muscular dystrophy; GK: Glycerol kinase; HY: Histocompatibility antigen Y; IL 3 RA: Interleukin 3 receptor alpha; IL 9 R: Interleukin 9 receptor; Kal 1: Kallmann syndrome 1; POLA: DNA polymerase alpha; RBMY: RNA-binding motif protein Y chromosome; SHOX: Short stature homeo box; SRY: Sex-determining region Y chromosome; USP 9 Y: Ubiquitin-specific protease 9 Y chromosome; XIST: X inactivation-specific transcript; ZFX: Zinc finger protein X-linked; FY: Zinc finger protein Y-linked http: //www. endotext. org/pediatrics 7/pediatrics 7. html

Структура SRY Structure of mouse and human SRY protein. The HMG DNA-binding domain is shown in red and the large glutamine-rich domain of the mouse SRY COOH terminus is in dark yellow. Nuclear translocation is mediated by one NLS (pink) at either end of the HMG domain. The NH 2 -terminal NLS is recognized and bound by calmodulin (Ca. M), whereas the COOH-terminal acts via importin. For both mouse and human SRY, a putative transactivation domain (TAD, light yellow) has been described. The hinge or bridge region (green) interacts with mouse SRY-interacting protein 1 (SIP 1/NHERF 2) and the KRAB-only protein, whereas human SRY interacts with SIP-1/NHERF 2 via its COOH terminus. Sex-reversing mutations in human SRY (marked by asterisks) leading to gonadal dysgenesis or hermaphroditism are mainly found in the HMG domain.

Гонады: мышь, 12, 5 суток развития

• General transcription factors, like LHX 1, EMX 2 and PAX 2, are necessary for intermediate mesoderm development. • The gonadal ridge differentiates from the intermediate mesoderm following the action of SF 1, which is regulated by LHX 9 and WT 1. • SRY expression, activated by WT 1 and GATA 4, induces testis differentiation, characterized by an increase in SOX 9 and a decrease in DAX 1 expression. The effect of SRY overcomes the action of genes that have been proposed to repress testicular differentiation, like WNT 4, RSPO 1, βcatenin and FOXL 2. http: //www. endotext. org/pediatrics 7/pediatrics 7. html

Дифференцировка клеток Сертоли Differentiation of pre-Sertoli cells into Sertoli cells. Nonpolarized, dispersed somatic cells visualized by SOX 9 immunofluorescence (green) represent pre-Sertoli cells at the 24 -tail somite (ts) stage. A few hours later, by 28 ts, these cells become polarized, forming epithelial aggregates that assemble into testis cords; at this stage they are referred to as Sertoli cells. PECAM-1 counterstaining (red) marks PGCs and endothelial cells. Model for cell-autonomous and prostaglandin-mediated upregulation of Sox 9 in pre-Sertoli cells. Sry induces Sox 9 cell-autonomously either via a direct or indirect regulatory mechanism (1). Subsequently, Sox 9 maintains its own expression in an autoregulatory loop (2). In addition, Sry and/or Sox 9 serve to upregulate Pgds (3), which leads to prostaglandin D 2 (PGD 2) synthesis (4) and secretion. PGD 2 can act by binding to its receptor DP (5), to upregulate Sox 9 expression in a paracrine, and possibly also an autocrine manner (6). Thus cells that do not express Sry or fail to reach a threshold of Sry expression can be induced to upregulate Sox 9 and differentiate as Sertoli cells. [Adapted from Smith et al.

Клетки Сертоли и молекулярные каскады дифференцировки клеток гонады по мужскому типу Double-headed arrows, binding to a receptor; colored arrows (blue, red, green), differentiation of precursor cells into testis-specific cell types; black, bold arrow, gene important for cellular process. Physiol Rev • VOL 87 • JANUARY 2007

Proposed signaling pathways for AMH. Binding of AMH to its type 2 receptor, AMHR 2, probably triggers the formation of a complex between AMHR 2 and one or several candidate type 1 receptors, ALK 2, 3 and 6. Activated type 1 receptor(s) phosphorylate Smad molecules 1, 5 or 8, which then bind to Smad 4 and enter the nucleus to activate BMP-specific reporter genes XVent 2 and Tlx 2. Smad 6 inhibits this pathway. Putative accessory signaling pathways include nuclear translocation of ß-catenin and its binding to LEF-1 and induction of NFκB nuclear binding activity. In the absence of AMH, NF-κB subunits p 65/Rel. A and p 50/NF-κB are retained in the cytoplasm by the inhibitory protein IκBa. AMH releases the subunits from this inhibition and allows them to translocate into the nucleus. Abbreviations: ALK, activin-like kinase; AMH, anti-Müllerian hormone; AMHR 2, AMH receptor type 2; BMP, bone morphogenetic protein; LEF-1 lymphoid enhancer factor 1; NFκB, nuclear factor κB; Smad, mothers against decapentaplegic related gene product, TGF-β, transforming growth factor.
")
Мезонефрос и закладка гонады (эмбрион человека, 1, 5 мес. , сагиттальный срез)
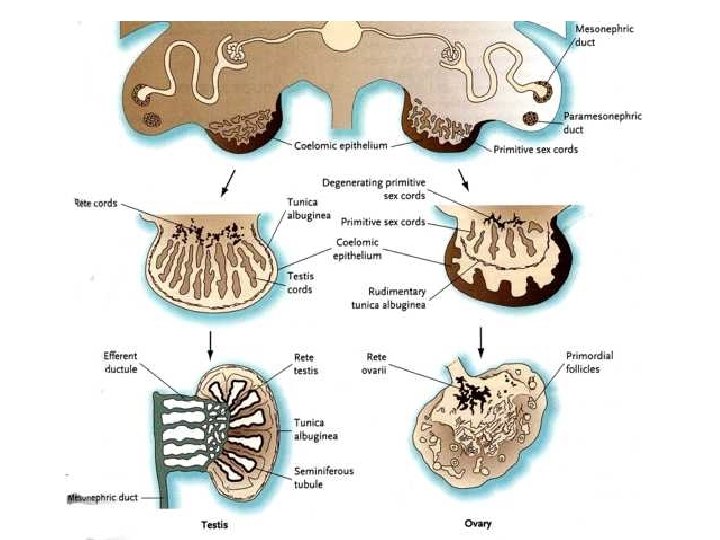

• Like in the male, general transcription factors, as LHX 1, EMX 2 and PAX 2, are necessary for intermediate mesoderm development. • The gonadal ridge differentiates from the intermediate mesoderm following the action of SF 1, which is regulated by LHX 9 and WT 1. Gene(s) in the DSS region of the X chromosome (DAX 1, MAGEB, other? ), as well as WNT 4 and FST should be expressed to antagonize testis differentiation. RSPO 1 and β-Catenin are both anti-testis and pro-ovary genes, essential for early ovarian differentiation. Germ cell colonization (dependent on BMP family members, SCF and its receptor c-kit, WNT 4 and FST) and meiosis (dependent on the existence of two X chromosomes as well as several factors like DAZLA, MSH 5, STRA 8 and DMC 1) are essential for fetal ovary stabilization. A number of other factors are involved in early folliculogenesis (FOXL 2, neurotrophins and neurotrophin tyrosine kinase receptors) http: //www. endotext. org/pediatrics 7/pediatrics 7. html

Сайты по теме лекции: Эмбриональное кроветворение у человека: http: //hematologiya. ru/teoriya/stroenie-kletki. -gemopoehz. /gemopoehz-u-ehmbriona-i-ploda. htm Дифференцировка пола по мужскому и по женскаму типу: http: //www. endotext. org/pediatrics 7/pediatrics 7. html
6be6e4d5ee1c64c33f15dcc5054149f9.ppt