lct1_community_2012.ppt
- Количество слайдов: 48



ЭКОЛОГИЯ СООБЩЕСТВ COMMUNITY ECOLOGY

низкие Биосфера Биогеоценозы Сообщества Популяции интег ратив связи ные УРОВНИ ОРГАНИЗАЦИИ БИОЛОГИЧЕСКИХ СИСТЕМ Организмы высокие
ЦЕНОЛОГИЯ")
СИНЭКОЛОГИЯ СООБЩЕСТВ= =эволюционная экология Дж. Хатчинсон 1957, 1959 Р. Мак. Артур, 1958 -1972 БИО(ГЕО)ЦЕНОЛОГИЯ Изучение биологического разнообразия и факторов его ограничивающих. Акцент на горизонтальные связи V. Shelford, 1913; Сукачев В. Н. 1928 Изучение потоков вещества и энергии. Акцент на вертикальные связи. Пищевые цепи
 G.")
A. LOTKA, 1925, V. VOLTERRA, 1926 Г. Ф. ГАУЗЕ ( G. GAUSE, 1934) G. Е. HUTCHINSON R. H. MACARTHUR Thomas SHOENER, Ted CASE, Erik PIANKA Martin CODY, Jared DIAMOND, Michael ROSENZWEIG, James H. BROWN D. LACK, 1947

ОПРЕДЕЛЕНИЯ СООБЩЕСТВА

СООБЩЕСТВО = БИОЦЕНОЗ, БИОГЕОЦЕНОЗ Сочетание популяций растений, животных микроорганизмов, взаимодействующих друг с другом и со средой обитания, образующих единую функциональную систему со свойственными ей структурой, разнообразием объектов (видов), конкретными отношениями их со средой, уникальной историей развития и специфическими функциями. R. Whittaker Взаимодействие утверждается

гильдия cовокупность популяций, использующих одну группу ресурсов функционально сходным образом. Взаимодействие предполагается, хотя по определению не необходимо
 – Совокупность обитающих совместно популяций филогенетически близких видов. ВЗАИМОДЕЙСТВИЕ НЕ")
АССАМБЛЕЯ, СООБЩЕСТВО (ASSEMBLAGE, COMMUNITY) – Совокупность обитающих совместно популяций филогенетически близких видов. ВЗАИМОДЕЙСТВИЕ НЕ ОБЯЗАТЕЛЬНО

Сообщество в понимании школы Р. Мак. Артура ВЗАИМОДЕЙСТВИЕ УТВЕРЖДАЕТСЯ

Гипотетико-дедуктивный подход Р. Мак. Артура:

Гипотетико-дедуктивный подход Р. Мак. Артура: 1. Поиск закономерностей = “PATTERNS”. 2. Внимание главным факторам. 3. Формализация результата эмпирических исследований, создание гипотезы (теории) и выражение ее в математической форме. 4. Теория руководит исследователем в его работе. 5. Новые эмпирические данные для подтверждения или опровержения теории 6. Широкое использование математического моделирования и использование наблюдений для проверки и совершенствования моделей.
 Зеленая (D. virens) Еловая (D. fusca) Каштановая (D.")
Dendroica Северовостока США Миртовая (D. coronata) Зеленая (D. virens) Еловая (D. fusca) Каштановая (D. castanea) Тигровая (D. tigrina)

Зеленая Еловая Миртовая Каштановая Тигровая % % времени наблюдений Зависима от пиков численности насек.
 равновесие со средой б) недостаток ресурсов в) конкуренция действует постоянно г) изменение")
Предполагается: а) равновесие со средой б) недостаток ресурсов в) конкуренция действует постоянно г) изменение условий (интенсивности конкуренции) сразу отражаются на структурной организации сообщества д) отношения между видами в природе суть результат действия естественного отбора

КОНКУРЕНЦИЯ – ГЛАВНЫЙ ФАКТОР

Вопросы в формулировках шк. Мак. Артура. 1. Структурированы ли сообщества (предвзятость? методические ошибки? статистические артефакты? ) 2. Насколько часто мы имеем дело с системами в состоянии равновесия? 3. Насколько общими являются известные примеры "конкурентных сообществ"? 4. Какие факторы кроме конкуренции аналогичным образом детерминируют структуру сообщества? 5. На экологических шкалах какого масштаба проявляется действие конкуренции? 6. Какова вообще роль теории в исследовании многовидовых ассоциаций животных?

НЕ ЯВЛЯЮТСЯ ЛИ СООБЩЕСТВА, СЛУЧАЙНЫМИ СКОПЛЕНИЯМИ НЕЗАВИСИМЫХ ПОПУЛЯЦИЙ ?

D A PSEUDOMYS SIU RI D S ПРАВИЛО ОРГАНИЗАЦИИ “АССАМБЛЕЙ” БЛИЗКИХ ВИДОВ (B. FOX, 1989). 0 1 2 RATTUS На вересковых пустошах в Австралии живут сумчатые крысы из сем. Dasyuridae, aборигенные грызуны Pseudomys и крысы Rattus

D A PSEUDOMYS SIU RI D S Биотопические группировки грызунов формируются таким образом, что включение другого вида того же рода (или той же функциональной группы) возможно лишь после того как произошло включение вида другого рода (другой функциональной группы) 0 1 Разрешенные сочетания Запрещенные сочетания 2 RATTUS

ГНЕЗДОВОЙ ТИП УПАКОВКИ СООБЩЕСТВ (NESTED SUBSET STRUCTURE; May, 1976; Patterson, Atmar, 1986; Kodric-Brown, 1993; Kelt et al. , 1995). Каждое менее богатое видами сообщество входит в состав более богатого видами сообщества (ассамблеи). Сообщества птиц и пресмыкающихся на островах, рыб в изолированных водоемах Центральной Австралии (пескарь, сом, твёрдоголовка, окунь), грызунов в пустынях.

РЕГУЛЯРНОЕ РАСПРЕДЕЛЕНИЕ ПО РАЗМЕРАМ ГРЫЗУНОВ В СООБЩЕСТВАХ ПУСТЫНЬ СЕВЕРНОЙ АМЕРИКИ (из J. H. Brown, 1975)

РЕГУЛЯРНОЕ РАСПРЕДЕЛЕНИЕ ПО РАЗМЕРАМ ГРЫЗУНОВ В СООБЩЕСТВАХ ПУСТЫНЬ СЕВЕРНОЙ АМЕРИКИ (из J. H. Brown, 1975)
 ХОЛИСТИЧЕСКИЙ = Изучение общих закономерностей")
ПОДХОДЫ ИНДИВИДУАЛИСТИЧЕСКИЙ = Внутри- и межпопуляционные взаимодействия (Пары видов) ХОЛИСТИЧЕСКИЙ = Изучение общих закономерностей организации многовидовых сообществ

ЧИСЛО ВИДОВ ГРАНИЦЫ СООБЩЕСТВА 1 2 3 ЧИСЛО ПРОБНЫХ ПЛОЩАДОК

ВОПРОСЫ ЭКОЛОГИИ СООБЩЕСТВ 1. ФАКТОРЫ, ОПРЕДЕЛЯЮЩИЕ ЧИСЛО ОБИТАЮЩИХ СОВМЕСТНО ВИДОВ 2. ПРЕДЕЛЫ ЭКОЛОГИЧЕСКОГО СХОДСТВА И ПУТИ РАЗДЕЛЕНИЯ РЕСУРСОВ СОВМЕСТНО ОБИТАЮЩИМИ ВИДАМИ 3. ПРИНЦИПЫ СТРУКТУРНОЙ ОРГАНИЗАЦИИ СООБЩЕСТВ 4. ПРИНЦИПЫ ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ СООБЩЕСТВ = ИЗУЧЕНИЕ МЕЖВИДОВОЙ КОНКУРЕНЦИИ 5. СООТНОШЕНИЕ МЕЖДУ СТРУКТУРНОЙ И ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИЕЙ СООБЩЕСТВА

ЭКОЛОГИЧЕСКАЯ НИША КАК ХАРАКТЕРИСТИКА ИСПОЛЬЗОВАНИЯ ПОПУЛЯЦИЕЙ РЕСУРСОВ СРЕДЫ

Определения экологической ниши Дж. Гриннел J. Grinell 1917, 24, 28. Положение вида в сообществе. Акцент на пространственное распределение и поведение (выбор условий обитания). Ч. Элтон (C. Elton, 1927). Место в биотической среде = функциональная роль в сообществе. Акцент на биотические отношения (пища, враги). Л. Дайс (L. Dice, 1952). Адаптация с условиям местообитания. Акцент на пространственное распределение. Дж. Кларк (G. Clark, 1954). Подразделение на “функциональную нишу” и “нишу места”. Дж. Хатчинсон (G. Hatchinson, 1957). Весь диапазон условий, в котором живет и воспроизводит себя особь, популяция. Акцент на пространственное распределение. Ю. Одум (E. Odum, 1959). Положение, статус вида в структуре сообщества. Акцент на функциональной роли в сообществе. Местообитаниеадрес, а ниша – “профессия вида”. Э. Пьянка (E. Pianka, 1978). Вся совокупность адаптаций индивида к окружающей среде в широком смысле. В применении к особи, популяции, виду. Т. Шёнер (T. Shoener, 1977). Место вида (популяции) в структуре сообщества, определяемое его способностью осваивать среду.

Экологи об экологической нише Автор Дж. Гриннел J. Grinell, 1917, 24, 28 Ч. Элтон (Ch. Elton, 1927) Л. Дайс (L. Dice, 1952) Размерность определяемого пространста Положение по отношению к объекту Местообитание, физичес- Первоначально внешнее, в последствии кие условия среды внутреннее (как свойство организма, вида) Трофо-энергетические характеристики (пища, размеры тела) Местообитание, условия среды Внутреннее (свойство организма, вида) В основном внешнее
 Дж. Хатчинсон (G. Hatchinson, 1957). Ю. Одум (E.")
Автор Дж. Кларк (G. Clark, 1954) Дж. Хатчинсон (G. Hatchinson, 1957). Ю. Одум (E. Odum, 1959) Э. Пьянка (E. Pianka, 1978) Т. Шёнер (T. Shoener, 1977) Размерность определяемого пространста Положение по отношению к объекту Местообитание, условия Внешнее (разные виды в среды, трофо-энергетичес- разных регионах могут кие характеристики занимать сходные ниши) (“ниша места”и”функциональная ниша”) Вся совокупность условий и ресурсов среды Внешнее (не является функцией поведения) Вся совокупность адаптаций Внутреннее (свойство организма, популяции, вида)
 унимодальны")
ПРИСПОСОБЛЕННОСТЬ ГРАДИЕНТ РЕСУРСА Кривые приспособленности (толерантности) унимодальны
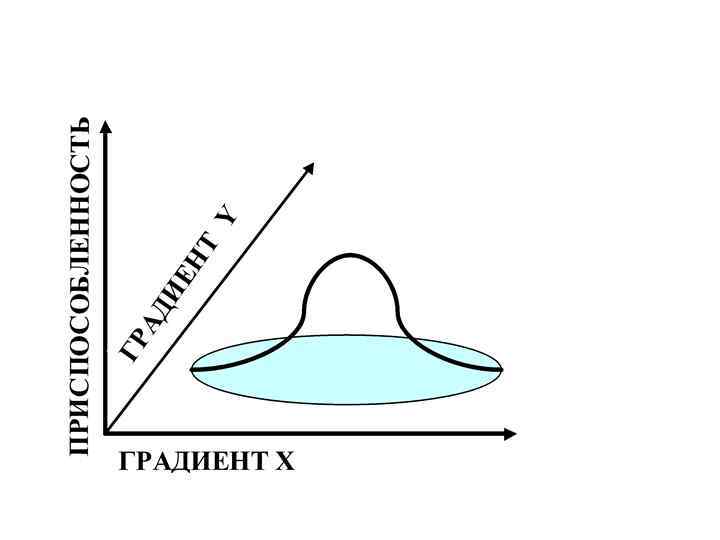
 ниша – вся совокупность оптимальных условий для существования вида Реализованная (постконкурентная) ниша")
Фундаментальная (преконкурентная) ниша – вся совокупность оптимальных условий для существования вида Реализованная (постконкурентная) ниша – фактический диапазон условий

УТИЛИЗАЦИЯ РЕСУРСА ВЫСОКИЙ ВНУТРИФЕНОТИЧЕСКИЙ КОМПОНЕНТ ВЫСОКИЙ МЕЖФЕНОТИЧЕСКИЙ КОМПОНЕНТ ГРАДИЕНТ РЕСУРСА

приспособленность градиент ресурса

обилие ШИРИНА НИШИ ПО ГРАДИЕНТУ РЕСУРСА РАСПРЕДЕЛЕНИЕ РЕСУРСА обилие градиент ресурса ВИД В градиент ресурса Без учета распределения ресурса B A< B B С учетом распределения ресурса BA>BB
 можно перемножать.")
РЕСУРС 2 РЕСУРС 1 Измерения независимы (высота и вероятность поимки жертвы) можно перемножать.
 НИШИ ЭНТРОПИЙНАЯ МЕРА РАЗНООБРАЗИЯ ШЕННОНА-УИВЕРА H=- Pilog. Pi ПОКАЗАТЕЛЬ РАЗНООБРАЗИЯ СИМПСОНА D=")
ШИРИНА (ОБЪЕМ) НИШИ ЭНТРОПИЙНАЯ МЕРА РАЗНООБРАЗИЯ ШЕННОНА-УИВЕРА H=- Pilog. Pi ПОКАЗАТЕЛЬ РАЗНООБРАЗИЯ СИМПСОНА D= Pi 2 ИНДЕКС ХИЛЛА D=1/ Pi 2 , где Pi - доля ресурса (градации i в общем спектре используемых ресурсов )

ИНДЕКСЫ ПЕРЕКРЫВАНИЯ НИШ 1. Мера пересечения множеств Шорыгина 2. Вероятностные меры 2. 1. АССИМЕТРИЧНЫЕ ИНДЕКСЫ ЛЕВИНСА 2. 2. ИНДЕКС МОРИСИТЫ 2. 3. ИНДЕКС ПЬЯНКИ

ПРОБЫ (ПЛОЩАД- ПЕРЕМЕННЫЕ=ХАРАКТЕРИСТИКИ КИ ОПИСАНИЙ И СРЕДЫ В ПЕРВИЧНЫХ ОПИСАНИЯХ V 1 V 2 V 3 V 4 V 5 V 6 УЧЕТОВ) 1 (+) 2 (-) 3 (-) 4 5 6 7 8 9 10 0, 1 0, 4 0, 5. . 0, 5 0, 7 0, 8. . . . При Анализе главных компонент мы строим таблицу, описывая все площадки (где есть и где нет зверей), затем по корреляционной матрице строим диаграмму. В ней можем оценить положение ниш.

ВИД B ВИД А ПРОБЫ (ПЛОЩАД- ПЕРЕМЕННЫЕ=ХАРАКТЕРИСТИКИ СРЕДЫ В ПЕРВИЧНЫХ ОПИСАНИЯХ КИ ОПИСАНИЙ И V 1 V 2 V 3 V 4 V 5 V 6 УЧЕТОВ) 1 (+) 2 (+) 3 (+) 4 5 6 (+) 7 (+) 8 (+) 9 10 0, 1 0, 4 0, 5. . 0, 1 0, 5 0, 4 0, 5 0, 7 0, 8. . 0, 5 0, 8 0, 7 . . . При дискриминантном анализе описываются только места поимок столько раз сколько попался зверь, затем вычисляется отношение дисперсии к общей дисперсии и строится диаграмма по дискриминантным осям.

АНАЛИЗ ГЛАВНЫХ ДИСКРИМИНАНТНЫЙ КОМПОНЕНТ АНАЛИЗ A 2 PC DF A PC 1 D FA 1

СТРУКТУРА СООБЩЕСТВ ПУСТЫННЫХ ГРЫЗУНОВ. ОРДИНАЦИЯ ПРОСТРАНСТВЕННЫХ НИШ 12 ВИДОВ В СИСТЕМЕ КООРДИНАТ, ОПРЕДЕЛЕННОЙ ДВУМЯ ОСНОВНЫМИ ДИСКРИМИНАНТНЫМИ ФУНКЦИЯМИ
 (МЕКСИКА) Из Shenbrot et al. , 2000")
СООБЩЕСТВО ГРЫЗУНОВ ЦЕНТРАЛЬНОЙ ЧИУАУА ПУСТЫНИ ТАР (ИНДИЯ) (МЕКСИКА) Из Shenbrot et al. , 2000

СТРУКТУРА СООБЩЕСТВ ПУСТЫННЫХ ГРЫЗУНОВ ПО РЕЗУЛЬТАТАМ КЛАСТЕРНОГО АНАЛИЗА ПЕРЕКРЫВАНИЙ НИШ


ГРАДИЕНТ РЕСУРСА 2 ЦИКЛЫ РАЗВИТИЯ ТАКСОНОВ И СПЕЦИАЛИЗАЦИЯ с ГРАДИЕНТ РЕСУРСА 1

ДИФФУЗНАЯ КОНКУРЕНЦИЯ С В D I А G E F
 с a")
вид А вид В местообитание Гипотеза компрессии (Mac. Arthr & Wilson, 1967) с a d e f g i h b j j пища Под давлением возросшей межвидовой конкуренции вид снижает число используемых местообитаний, но разнообразие потребляемой пиши остается тем же или увеличивается. Экологическое высвобождение как альтернатива.
lct1_community_2012.ppt