ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ.ppt
- Количество слайдов: 74





 Метод электрораздражения и регистрации электрических токов, токи покоя и действия (1848)")
Э. Дюбуа-Реймон (Германия) Метод электрораздражения и регистрации электрических токов, токи покоя и действия (1848)
 особенности действия постоянного тока на нерв и мышцу (1859)")
Э. Пфлюгер (Германия) особенности действия постоянного тока на нерв и мышцу (1859)
. Нобелевская премия 1903 г")
С. А. Аррениус Теория электролитической диссоциации (1887). Нобелевская премия 1903 г
 Определение равновесного потенциала Диффузия ионов через полупроницаемую мембрану (1888)")
В. Нернст (Германия) Определение равновесного потенциала Диффузия ионов через полупроницаемую мембрану (1888)
 первая ионная теория биотоков, основанная на теории электролитической диссоциации С.")
В. Ю. Чаговец (Россия) первая ионная теория биотоков, основанная на теории электролитической диссоциации С. Аррениуса (1896)

Алан Ходжкин Сэр Джон Экклс Нобелевская премия 1963 года в области физиологии и медицины за открытие ионных механизмов потенциала действия и передачи возбуждения Эндрю Хаксли
: Генез МП , причины поляризации -")
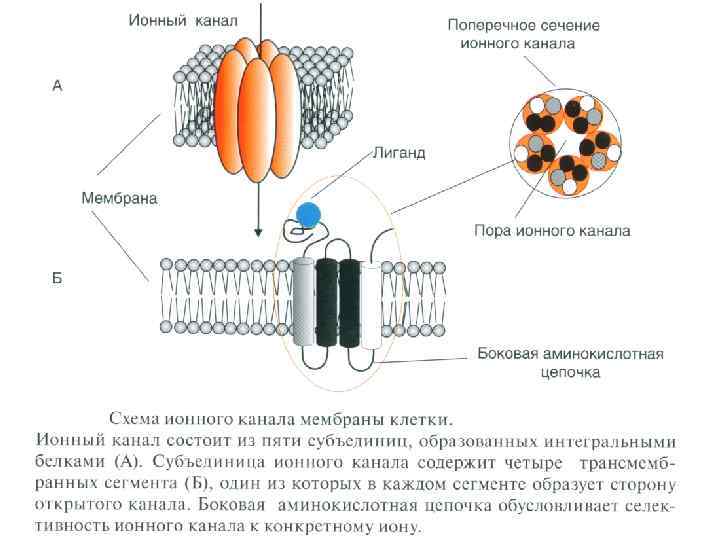
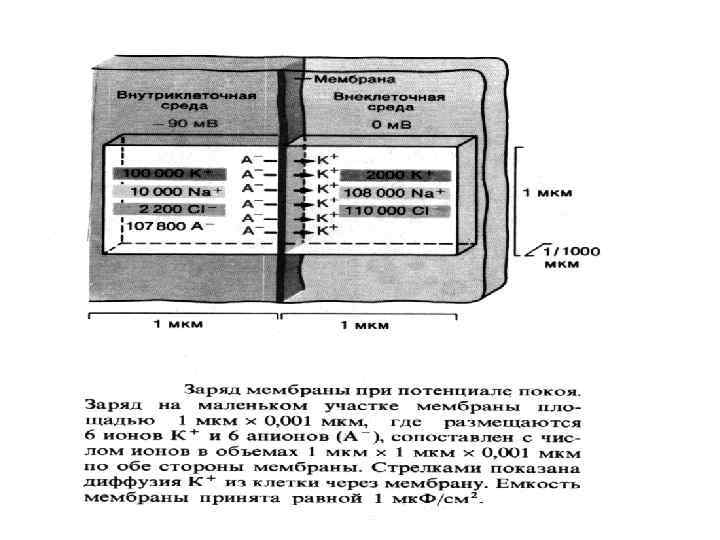
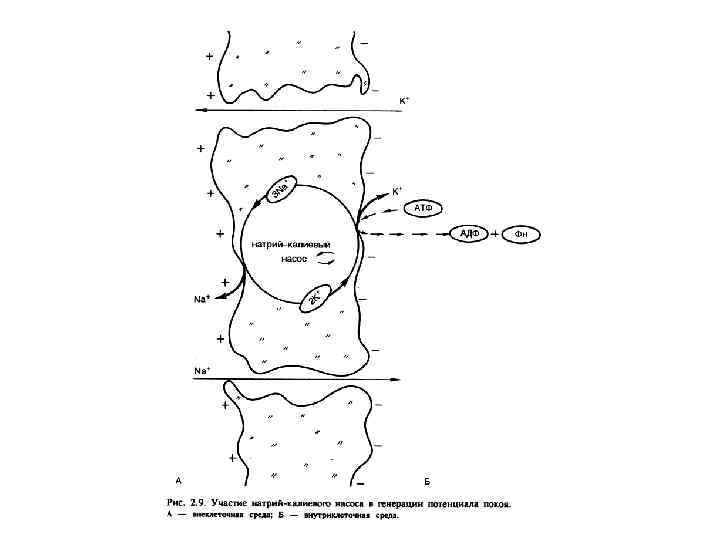
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ Мембранный отенциал (П. П. ): Генез МП , причины поляризации - природа МП - калиевая; - градиент концентраций ионов и в первую очередь К; - избирательная проницаемость для ионов К в покое; (выход К из клетки обуславливает положительный заряд на наружной поверхности мембраны; органические анионы, оставаясь в клетке в силу не проницаемости для них мембраны, обусловливают отрицательный заряд на внутренней поверхности).

Величина П. П. колеблется от -70 до -95 м. В. Потенциал покоя (М. П. ) зависит от: - селективной проницаемости клеточной мембраны для различных ионов. ; - различной концентрации ионов цитоплазмы клетки и ионов окружающей среды; - работы механизма К-Nа насоса.
 местное изменение ионной проницаемости мембраны. (первые признаки появляются при действии раздражителя")
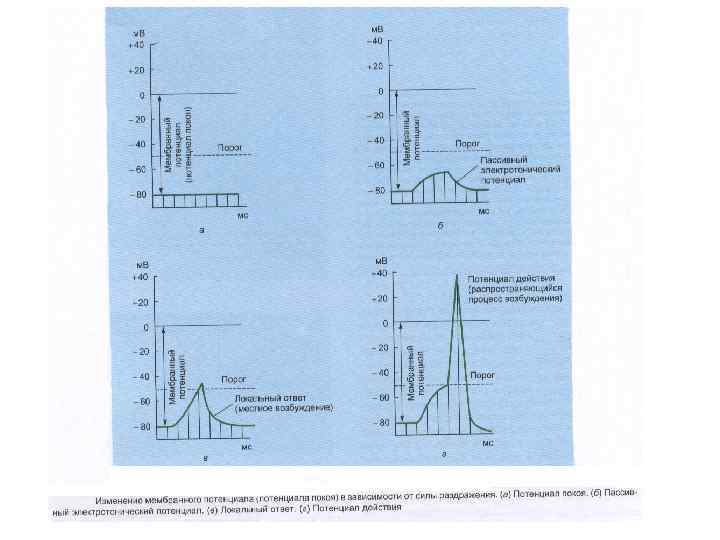
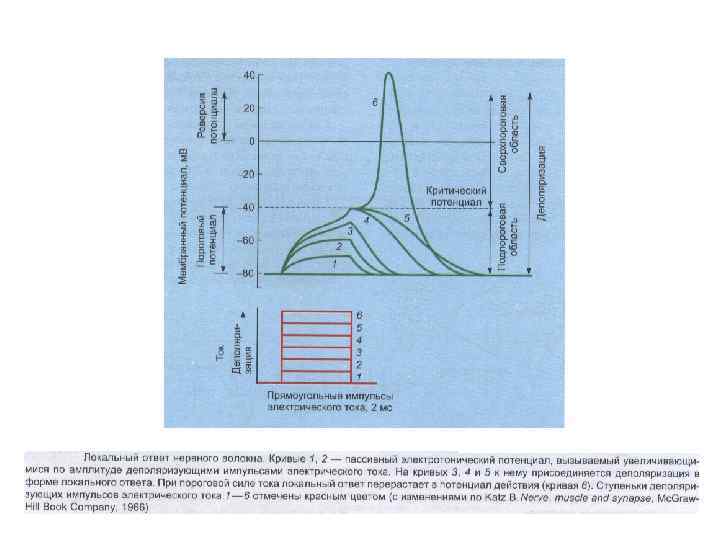
Локальный ответ (ЛО) местное изменение ионной проницаемости мембраны. (первые признаки появляются при действии раздражителя 50 - 70 % пороговой величины). ЛО отличается от ПД: - не имеет четкого порога возникновения; - сопровождается не абсолютной рефрактерностью, а наоборот повышенной возбудимостью; - - градуально зависит от силы деполяризующего тока (не подчиняется закону "все или ничего"); - не способен к бездекрементному распространению по нервному волокну.

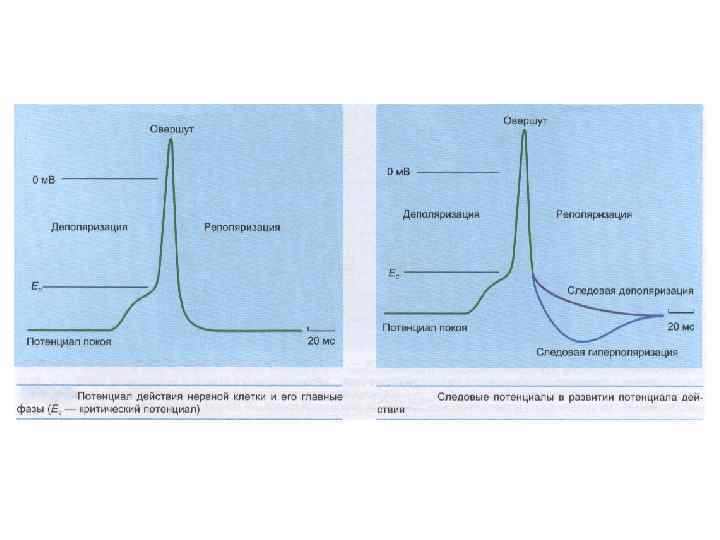
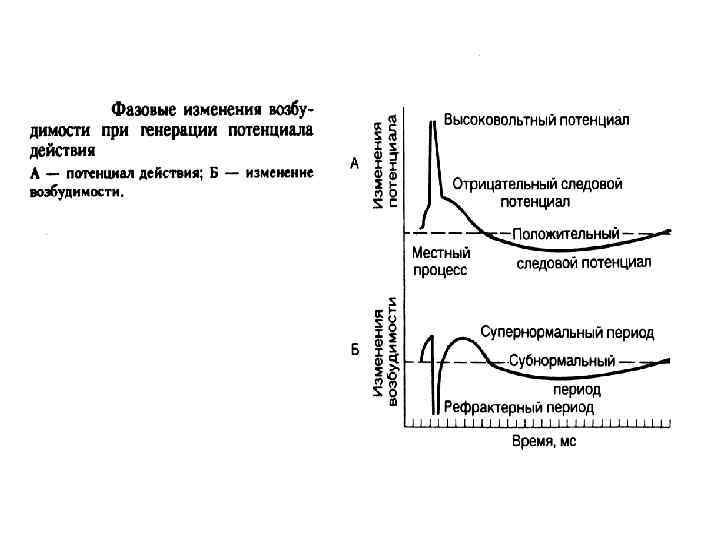
Потенциал действия - быстрое колебание мембранного потенциала, возникающее при возбуждении Фазы П. Д. : - фаза деполяризации (быстрой и медленной); - овершут - фаза реполяризации; - следовые потенциалы (деполяризационный и гиперполяризационный или отрицательный и положительный);
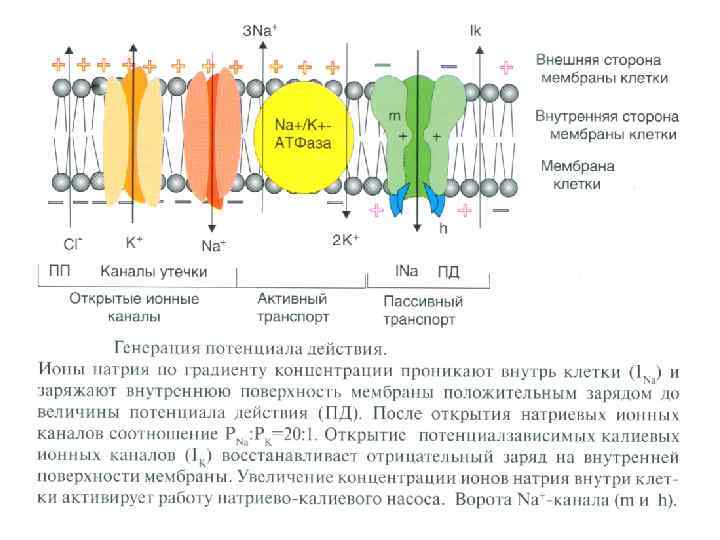
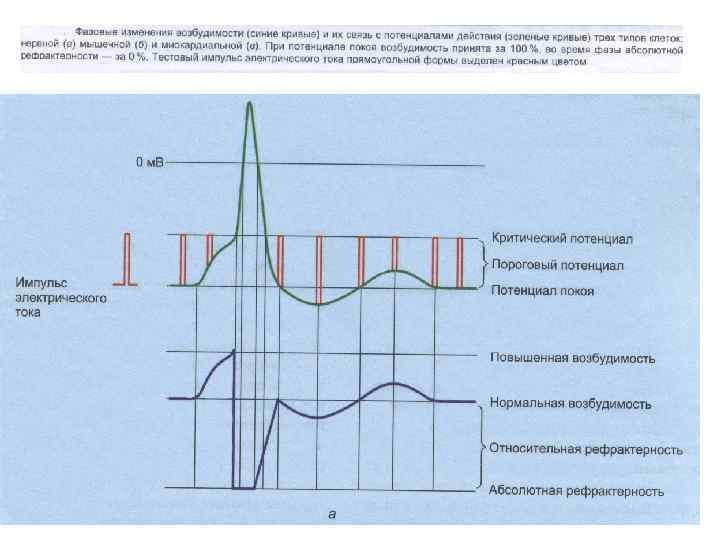
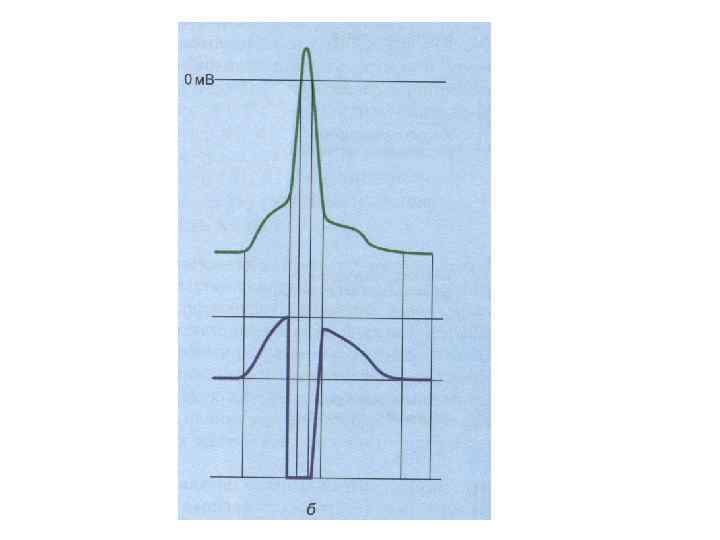
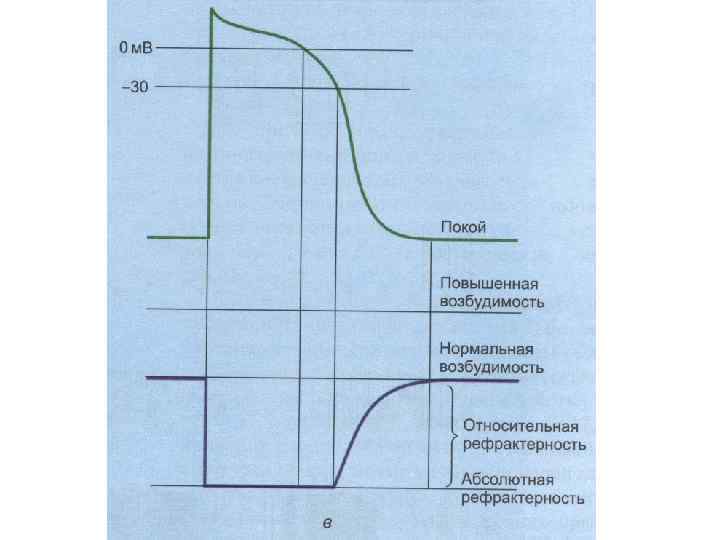
 кратковременное снижение возбудимости нервной и мышечной ткани во время и")
РЕФРАКТЕРНОСТЬ (refractaire - невосприимчивый) кратковременное снижение возбудимости нервной и мышечной ткани во время и вслед за ПД. Фазы (периоды) рефрактерности: - абсолютный рефрактерный период; - относительный рефрактерный период (восстановление возбудимости происходит прежде всего в наиболее возбудимых волокнах) - супернормальный период (фаза экзальтации); - субнормальный период.
 русский физиолог, ученик И. М. Сеченова основоположник учения")
Введенский Николай Евгеньевич ( (1852 -1922) русский физиолог, ученик И. М. Сеченова основоположник учения об общих закономерностях реагирования возбудимых систем организма (ввел понятия функциональной подвижности, оптимума и пессимума раздражения, парабиоз и др).
 максимальное число потенциалов действия, которое возбудимая ткань способна генерировать в 1")
Лабильность (функциональная подвижность) максимальное число потенциалов действия, которое возбудимая ткань способна генерировать в 1 секунду в соответствии с ритмом раздражения. Лабильность возбудимой ткани в первую очередь определяется продолжительностью периода рефрактерности и является показателем функционального состояния тканей;

ЗАКОНЫ РАЗДРАЖЕНИЯ возбудимых тканей: ЗАКОН СИЛЫ РАЗДРАЖЕНИЯ чем сильнее раздражение, тем сильнее ответная реакция возбудимой ткани (до известного предела); Закон силы для клетки: при повышении силы для подпороговых раздражителей растет амплитуда локального ответа. Закон силы для тканей и органов: чем сильнее раздражение, тем сильнее (до определенных пределов) функциональный ответ).
 и ответная реакция")
ЗАКОН ДЛИТЕЛЬНОСТИ РАЗДРАЖЕНИЯ чем длиннее раздражение, тем сильнее (до известного предела) и ответная реакция ткани (для проявления минимальной реакции ткани требуется определенная критическая длительность раздражения; при увеличении длительности увеличивается и ответная реакция; увеличение длительности за пределы максимальной величины не ведет к реакции ткани);
 нарастания раздражителя во времени, тем сильнее выражено раздражающее")
ЗАКОН ГРАДИЕНТА чем быстрее крутизна (скорость) нарастания раздражителя во времени, тем сильнее выражено раздражающее действие тока. Уменьшение скорости (крутизны) нарастания силы тока повышает порог раздражения При действии медленно нарастающей силы раздражителя генерации ПД не происходит так как возбудимая клетка «приспосабливается» к действию этого раздражителя (аккомодация тканей). Аккомодация клетки – исчезновение возбуждения при определенной минимальной крутизне нарастания силы раздражителя.
 при любой силе")
ЗАКОН "ВСЕ ИЛИ НИЧЕГО" Возбудимая ткань, дает максимальную ответную реакцию ("все") при любой силе порогового и надпорогового раздражения и не дает ("ничего") при подпороговых стимулах (установлен эмпирически Боудич, Готи, Вунд). (Установлено, что возбуждение возникает лишь при достижении стимула пороговой величины; в то же время понятие "порог" относительно т. к. изменяется реагирующая структура, меняющая скорость реакции и ее градуальность. Поэтому установлена лишь относительная не зависимость эффекта от силы действующего раздражителя: оказалось, что "все" или "ничего" тоже носят градуальный характер);

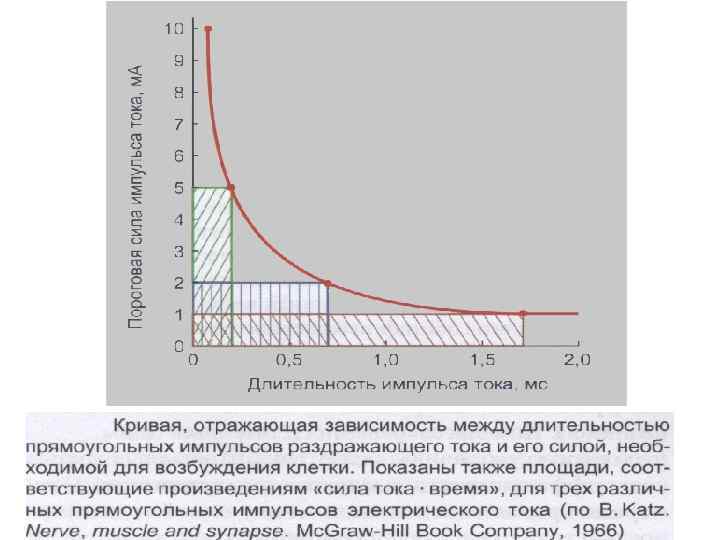
ЗАКОН о СООТНОШЕНИИ между СИЛОЙ и ДЛИТЕЛЬНОСТЬЮ РАЗДРАЖЕНИЯ (кривая "СИЛА-ВРЕМЯ; закон Гооверга-Вейса-Лапика; реобаза - хронаксия) пороговая сила любого стимула в известных пределах находится в обратной зависимости от его длительности (кривизны гиперболы). Ток ниже некоторой минимальной силы не вызывает возбуждения, как бы длительно он не действовал (горизонтальная часть гиперболы). При очень коротких стимулах, как бы не велика была сила раздражителя, возбуждения не возникает (вертикальная часть гиперболы).
; 2")
Кривая «силы–времени» . 1 — реобаза (минимальная сила раздражителя, способная вызвать ПД); 2 — удвоенная реобаза; 3 — кривая «силы-времени» ; а — полезное время (минимальное время, в течение которого ток, равный одной реобазе, вызывает генерацию ПД); б — хронаксия (минимальное время, в течение которого ток, равный двум реобазам, вызывает ПД).

ЗАКОНЫ проведения возбуждения по нервным волокнам - анатомической и физиологической целостности; - одностороннего проведения возбуждения; - изолированного проведения возбуждения; - проведение возбуждения без декремента.
 - возбуждение по нерву распространяется")
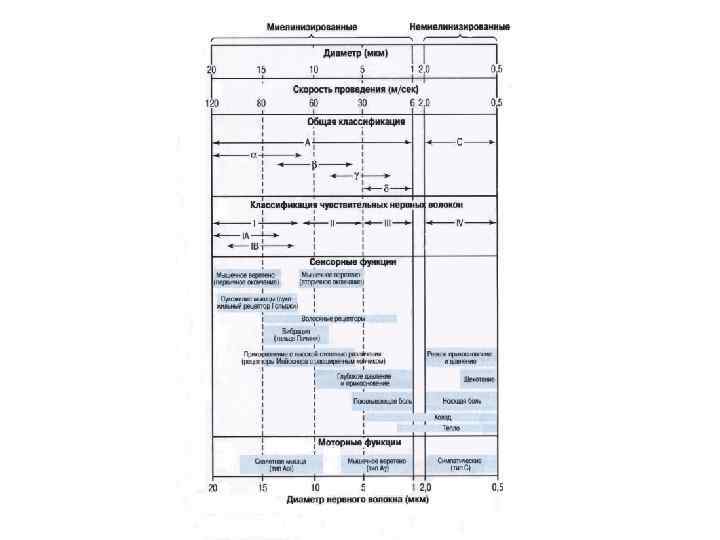
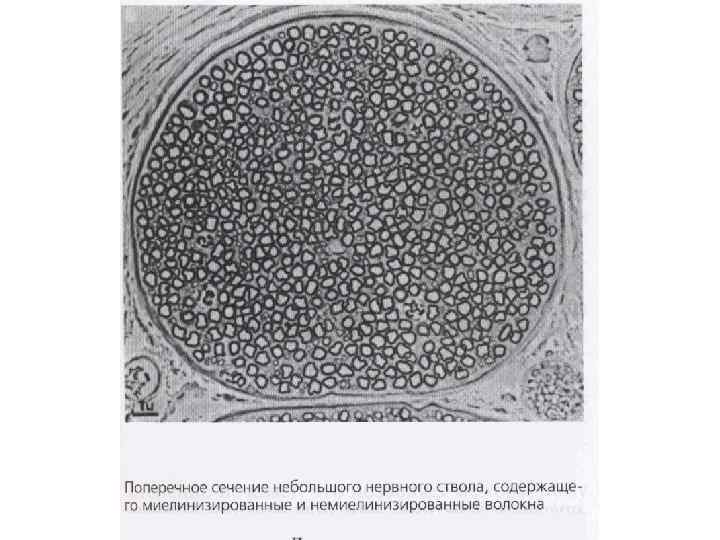
Законы проведения возбуждения по нервным волокнам (по Камкин, каменский) - возбуждение по нерву распространяется в обе стороны; - распространение возбуждения по нерву в обе стороны происходит с одинаковой скоростью; - возбуждение по нерву распространяется без затухания (без декремента); - необходимы анатомическая и физиологическая целостность нерва; - возбуждение распространяется по нервным волокнам нервного ствола изолировано; - нерв не утомляем; - в различных волокнах возбуждение распространяется с разной скоростью (зависит от сопротивлений среды окружающей волокно, аксоплазмы на единицу длины, мембраны аксона и диаметра волокна).
, Немецкий физиолог, ученик Дюбуа. Реймона, иностранный членкорреспондент Петербургской АН")
Пфлюгер, Эдуард Фридрих (1829 -1910), Немецкий физиолог, ученик Дюбуа. Реймона, иностранный членкорреспондент Петербургской АН (1894)
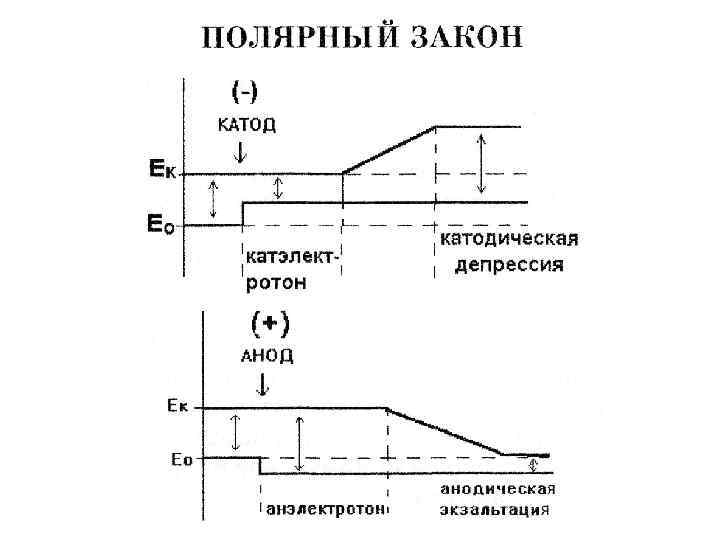
 1. В момент замыкания раздражающего тока (или возрастания его")
Закон полярности раздражения (Пфлюгер, 1859) 1. В момент замыкания раздражающего тока (или возрастания его силы) возбуждение возникает в области отрицательного полюса (катода) и отсюда распространяется на остальную часть мышцы или нерва; 2. В момент же размыкания тока (или его ослабления) возбуждение создается в области положительного полюса (анода); 3. При одной и той же силе тока раздражающее действие замыкания выражено сильнее, чем действие размыкания.
 Катэлектротон - повышение возбудимости и проводимости в области катода. Анэлектротон -")
Физиологический электротон (Пфлюгер) Катэлектротон - повышение возбудимости и проводимости в области катода. Анэлектротон - понижение возбудимости и проводимости в области анода.

При прохождении по нерву сильного постоянного тока повышенная возбудимость под катодом сменяется ее падением, до полного подавления возбудимости и проводимости – катодическая депрессия (Б. Ф. Вериго, 1883). Первоначальное снижение возбудимости под анодом сменяется ее повышением — анодная экзальтация.
 – область слияния внешних листков мембран")
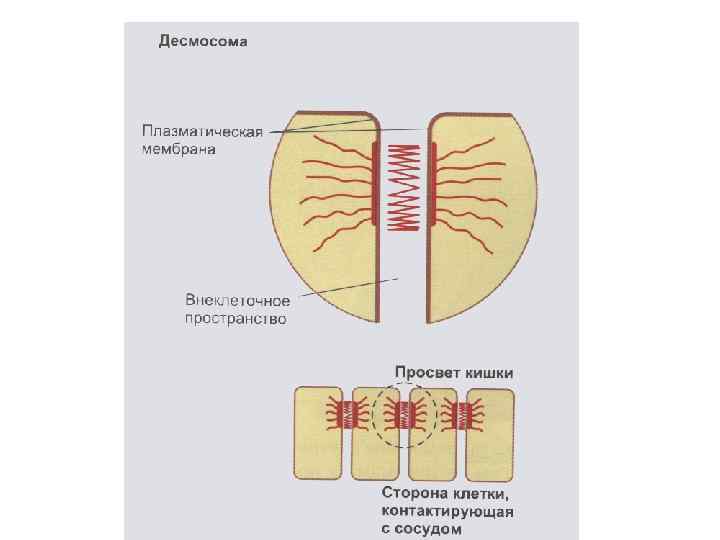
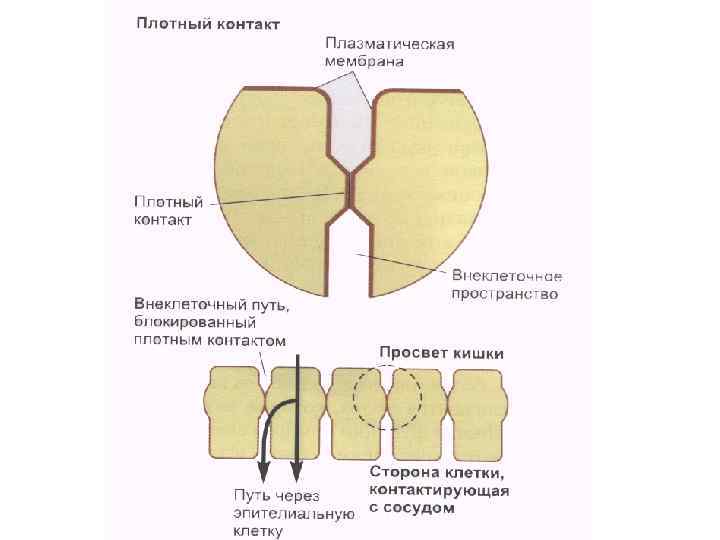
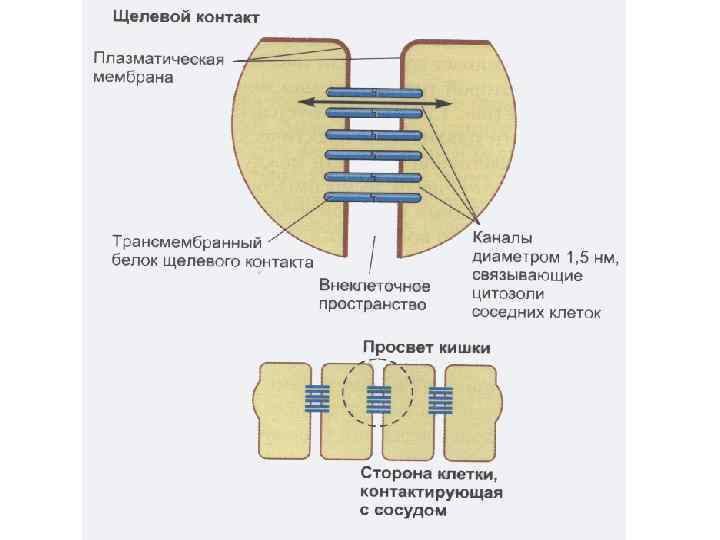
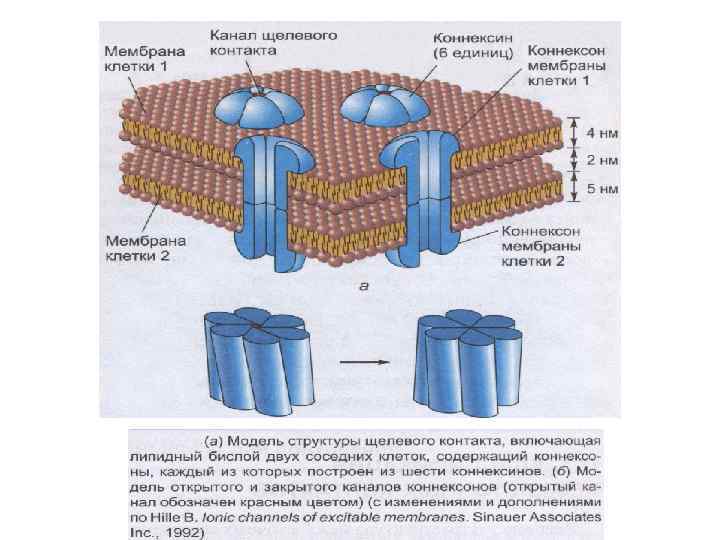
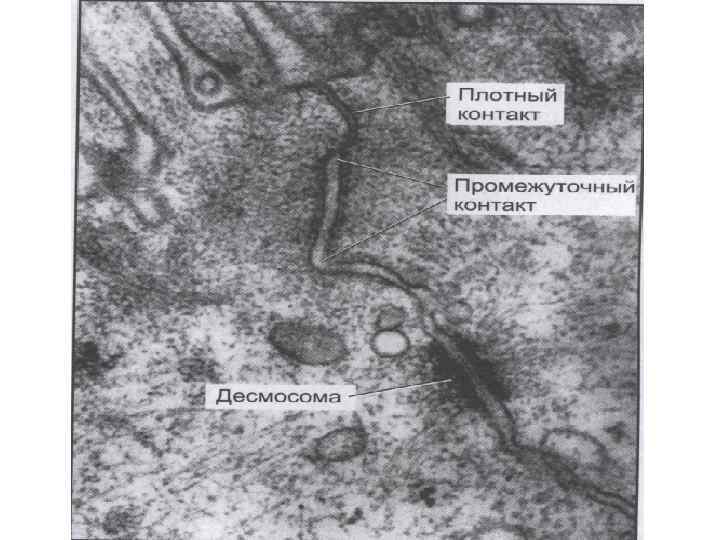
Межклеточные взаимодействия 1. Плотный контакт (tight junction) – область слияния внешних листков мембран соседних клеток с образованием одиночной электронно-плотной полосы вдоль контакта и представляет собой сеть ветвящихся тонких гребней. 2. Промежуточный контакт (intermediate junction) – зона, характеризующаяся наличием межклеточной щели шириной 150 – 200 А, заполненной гомогенным материалом низкой электронной плотности. 3. Десмосома (desmosome) –локальное дискообразное электронноплотное образование, располагающееся параллельно внутреннему листку каждой клеточной мембраны. 4. Септированный контакт (septate junction) – межклеточное пространство шириной 150 -170 А, пересекается септами (перегородками) шириной 40 -50 А, связывающими внешние поверхности соседних клеток. 5. Щелевой контакт (gap junction) – имеет в области тесного сближения мембран двух соседних клеток межклеточную щель шириной 20 -40 А.
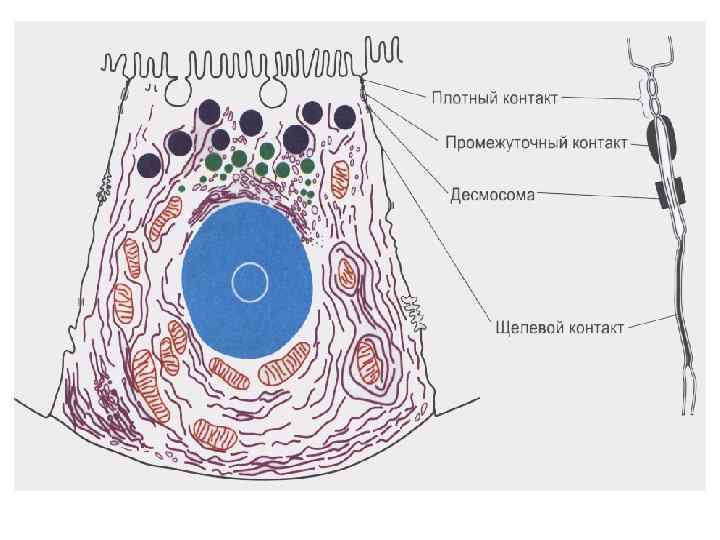

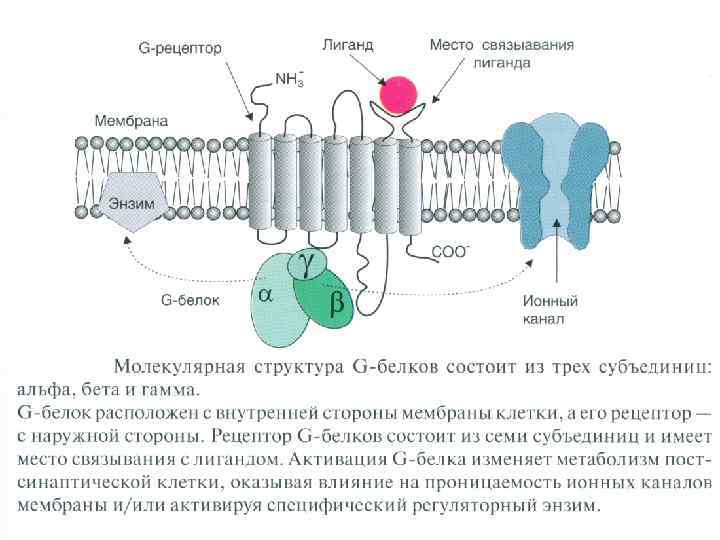
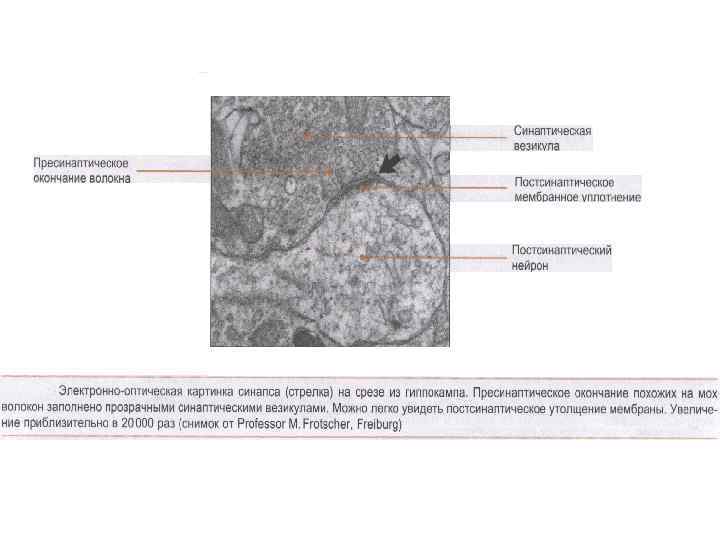
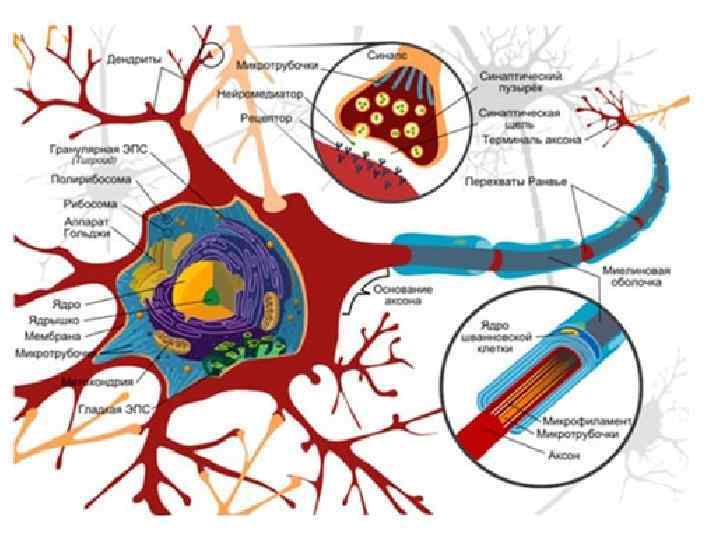
Синапс соединение между окончанием аксона и нервной клеткой, мышечной или железистой клеткой Ч. Шерингтон (1857 -1952) английский нейрофизиолог, описал принципы реципрокности и общего конечного пути Нобелевская премия (совместно с Э. Эдрианом) за открытие передачи информации в виде электрических импульсов

Классификация по механизму передачи информации: химические, электрические и смешанные по типу контактов: аксо - соматические и аксо - дендритические (до 10% всех синапсов мозга); кроме этого есть А-А, Д-Д, Д-С, С-Д, С-С. от вида выделяемого медиатора адренергические, холинергические, серотонинергические, дофаминергические, ГАМК-ергические и т. д. возбуждающие и тормозные центральные и периферические
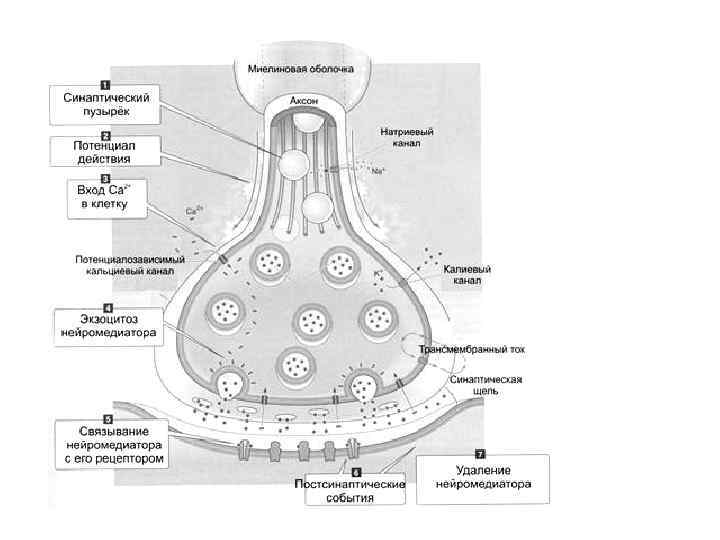
 - проведение нервного импульса по аксону; - секреция")
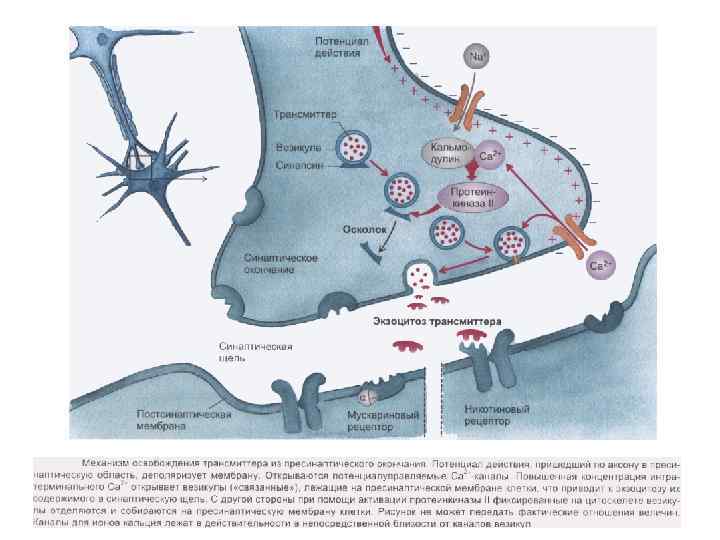
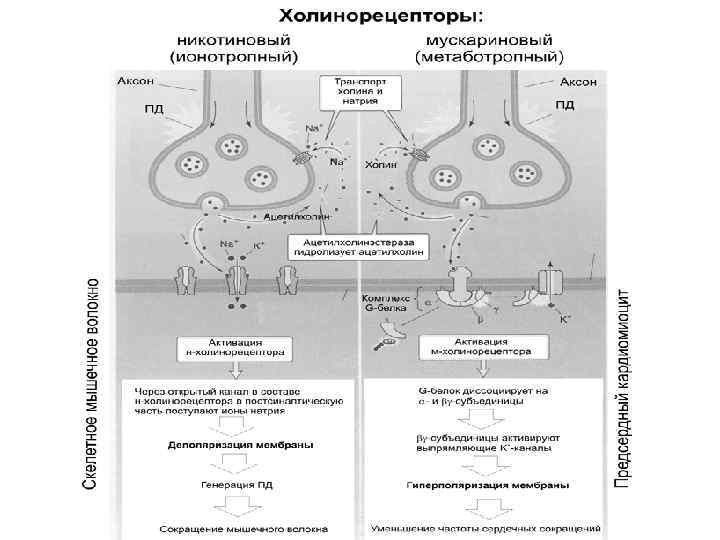
Этапы синаптической передачи возбуждения (химический синапс) - проведение нервного импульса по аксону; - секреция медиаторов нервным окончанием (происходит постоянно порциями, «квантами» ; - диффузия медиатора через синаптическую щель к постсинаптической мембране и взаимодействие с рецепторами; - инактивация отработанного медиатора в синаптической щели; - восстановительная работа в нервных окончаниях.
; - синаптическая задержка")
Особенности функциональных свойств химических синапсов - наличие медиатора (передача сигнала медиатором); - синаптическая задержка (0, 2 -0, 5 мс); - относительная медиаторная специфичность синапса; - одностороннее проведение возбуждения (в электрических возможна в обе стороны); - химические синапсы обеспечивают возбуждение и торможение, (электрические только возбуждение); - химические синапсы более термочувствительны и лучше сохраняют следы предшествующей активности; - наличие хемочувствительных рецептирующих каналов постсинаптической мембраны; - зависимость увеличения эффективности синаптической передачи от частоты использования синапса (эффект тренировки).
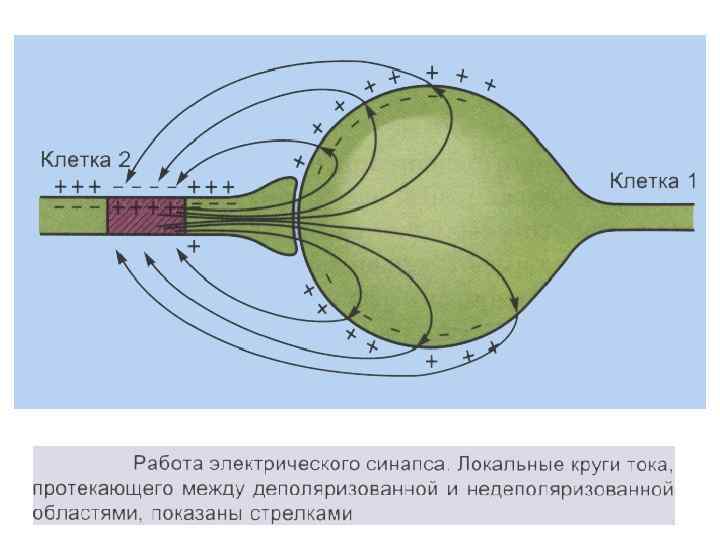
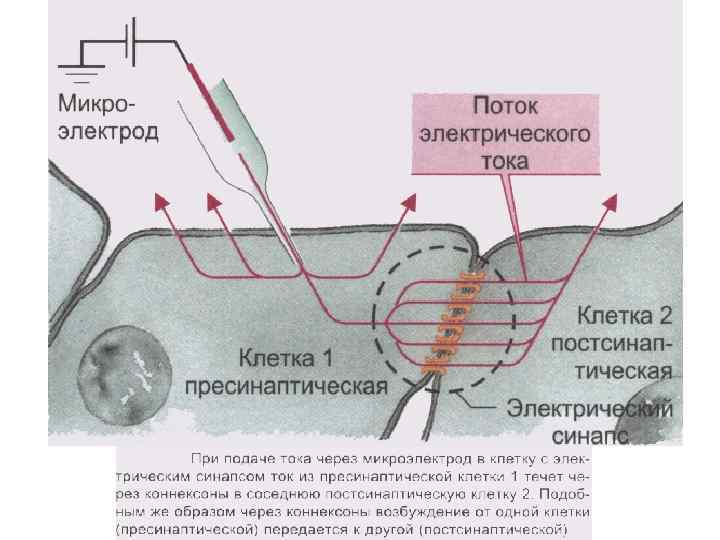
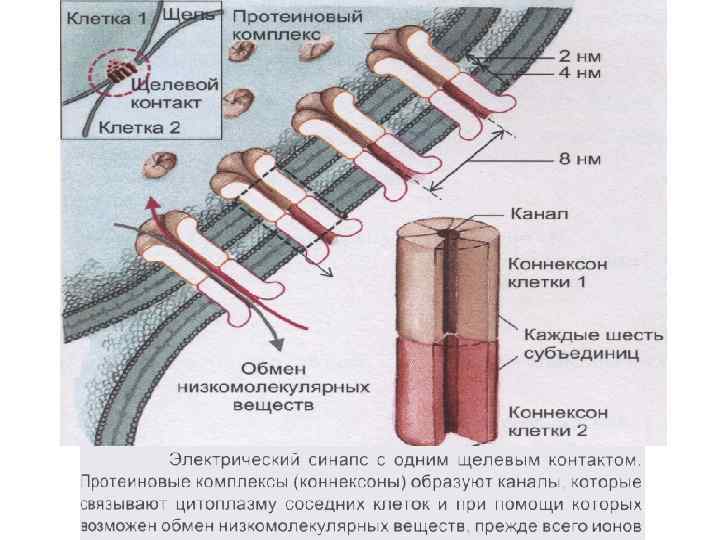

Электрические синапсы - малая величина синаптической щели; - микрофиламенты; - нет медиатора; - малая синаптическая задержка; - сравнительно мало утомляем; - устойчив к изменениям внешней и внутренней среды, что обеспечивает его высокую надежность; - деполяризация возникает как в пре- так и в постсинапсе.

Свойства синапсов - квантование медиатора; - трансформация ритма; - синаптическое облегчение; - синаптическая депрессия; - временная суммация; - пространственная суммацмя; - утомляемость.

Функциональное значение синапсов - одностороннее проведение возбуждения упорядочивает направление распространения возбуждения в ЦНС; - эффективность синаптической передачи может снижаться или увеличиваться, что происходит при увеличении или уменьшении тока ионов Са++ в пресинапс, что в свою очередь приводит к повышению или снижению количества выделяемого медиатора; - синапс представляет собой область действия многих фармакологических веществ; одни конкурируют с медиаторами, другие мешают медиаторам разрушаться и захватываться пресинапсом, третьи – усиливают или ослабляют действие тормозных медиаторов.
 биологически активное вещество, находящееся в нейроне в связанной, депонированной форме в")
Медиатор (нейропередатчик, нейротрансмиттер) биологически активное вещество, находящееся в нейроне в связанной, депонированной форме в синаптических пузырьках. Общие свойства медиаторов: - проявляют физиологические свойства в очень малых концентрациях (10 -7 - 10 -5 М) и только в свободной форме; - плохо поступают в мозг из кровотока; - реакционноспособны и лабильны; - неравномерно распределены в разных отделах мозга и периферической нервной системе; - большая скорость обмена; - действие кратковременно и в ограниченном пространстве; - действуют на незначительном расстоянии.
 с образованием возбуждающего постсинаптического потенциала")
Медиаторы вызывают два типа эффектов: - либо возбуждение (деполяризация) с образованием возбуждающего постсинаптического потенциала (ВПСП), сопровождаемый снижением величины потенциала покоя с -70 м. В до -10 м. В, с одновременным увеличением проницаемости к ионам Na и К; (АХ, НА, дофамин, серотонин, глутаминовая кислота и т. д); - либо торможение (гиперполяризация), с образованием тормозного постсинаптического потенциала (ТПСП), сопровождаемый увеличением потенциала покоя с -70 м. В до -80 м. В и увеличением ионной проницаемости для К и (или) СL (ГАМК, глицин).



Нобелевская премия за 2013 г в области физиологии и медицины Рис. 1. Слева: везикула представляет из себя пузырек, окруженный билипидным слоем — такой же мембраной, как та, что ограничивает клетку. Справа: электронная микрофотография синапса в месте контакта двух нейронов; хорошо видны синаптические пузырьки-везикулы, содержащие молекулы нейромедиатора. Изображения с сайта en. wikipedia. org и с сайта Стэнфордского университета
 и при температуре")
Рис. 2. Электронные микрофотографии дрожжевых клеток, растущих при нормальной температуре (B) и при температуре активации мутаций (D). Заметно значительное накопление везикул, содержащих выделяемые ферменты, внутри клетки. Рисунок из статьи: P. Novick & R. Schekman. Secretion and cell-surface growth are blocked in a temperature-sensitive mutant of Saccharomyces cerevisiae // Proc. Natl. Acad. Sci. USA. 1979. V. 76(4). P. 1858– 1862
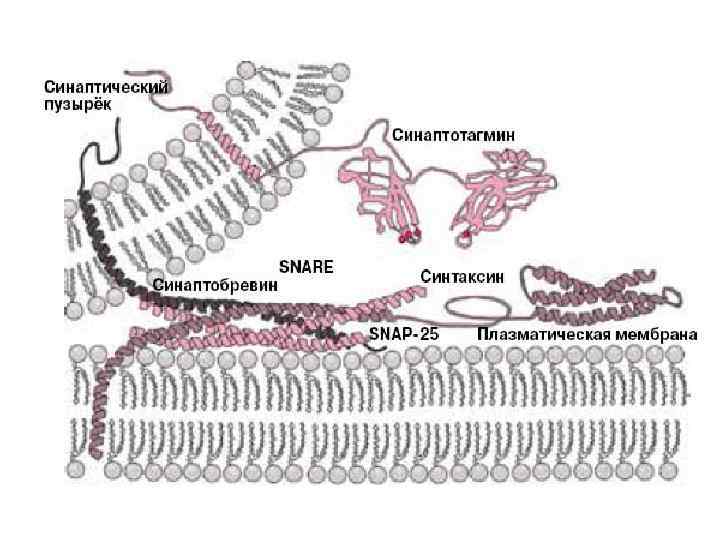
 между разными отделами клетки. Пузырьки (везикулы)")
Рис. 3. Схема, поясняющая принцип везикулярного транспорта (SNARE-гипотезу) между разными отделами клетки. Пузырьки (везикулы) отпочковываются от мембраны одной из органелл (например, эндоплазматического ретикулума) и получают «ключ» — один из белков семейства v-SNARE. Точная доставка осуществляется благодаря тому, что на целевой органелле есть специфический белок семейства t-SNARE, выступающий в роли «замка» . Изображение с сайта www. zoology. ubc. ca
ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ.ppt