

Дыхание растений.pptx
- Количество слайдов: 39
 ДЫХАНИЕ РАСТЕНИЙ Химизм и энергетика. Взаимосвязь дыхания и фотосинтеза
ДЫХАНИЕ РАСТЕНИЙ Химизм и энергетика. Взаимосвязь дыхания и фотосинтеза
 Дыхание – это физиологический процесс постепенного окисления органических веществ с выделением энергии, которая запасается в молекулах АТФ, являющихся, в свою очередь, донорами энергии для выполнения любой работы в клетке. Органические вещества, разрушающиеся во время дыхания, называют дыхательными субстратами.
Дыхание – это физиологический процесс постепенного окисления органических веществ с выделением энергии, которая запасается в молекулах АТФ, являющихся, в свою очередь, донорами энергии для выполнения любой работы в клетке. Органические вещества, разрушающиеся во время дыхания, называют дыхательными субстратами.
 Субстраты дыхания растений Вопрос о веществах, используемых в процессе дыхания, издавна занимал физиологов. Еще в работах И. П. Бородина (1876) было показано, что интенсивность процесса дыхания прямо пропорциональна содержанию в тканях растений углеводов. Это дало основание предположить, что именно углеводы являются основным веществом, потребляемым при дыхании (субстратом). В выяснении данного вопроса большое значение имеет определение дыхательного коэффициента. Дыхательный коэффициент (ДК) — это объемное или молярное отношение СО 2, выделившегося в процессе дыхания, к поглощенному за этот же промежуток времени О 2. При нормальном доступе кислорода величина ДК зависит от субстрата дыхания. Если в процессе дыхания используются углеводы, то процесс идет согласно уравнению С 6 Н 1206 +602 —> 6 С 02 + 6 Н 20. В этом случае ДК равен единице: 6 С 02/602 = 1
Субстраты дыхания растений Вопрос о веществах, используемых в процессе дыхания, издавна занимал физиологов. Еще в работах И. П. Бородина (1876) было показано, что интенсивность процесса дыхания прямо пропорциональна содержанию в тканях растений углеводов. Это дало основание предположить, что именно углеводы являются основным веществом, потребляемым при дыхании (субстратом). В выяснении данного вопроса большое значение имеет определение дыхательного коэффициента. Дыхательный коэффициент (ДК) — это объемное или молярное отношение СО 2, выделившегося в процессе дыхания, к поглощенному за этот же промежуток времени О 2. При нормальном доступе кислорода величина ДК зависит от субстрата дыхания. Если в процессе дыхания используются углеводы, то процесс идет согласно уравнению С 6 Н 1206 +602 —> 6 С 02 + 6 Н 20. В этом случае ДК равен единице: 6 С 02/602 = 1
 если разложению в процессе дыхания подвергаются более окисленные соединения, например органические кислоты, поглощение кислорода уменьшается, ДК становится больше единицы. Так, если в качестве субстрата дыхания используется яблочная кислота, то ДК = 1, 33. При окислении в процессе дыхания более восстановленных соединений, таких, как жиры или белки, требуется больше кислорода и ДК становится меньше единицы. Так, при использовании жиров ДК = 0, 7.
если разложению в процессе дыхания подвергаются более окисленные соединения, например органические кислоты, поглощение кислорода уменьшается, ДК становится больше единицы. Так, если в качестве субстрата дыхания используется яблочная кислота, то ДК = 1, 33. При окислении в процессе дыхания более восстановленных соединений, таких, как жиры или белки, требуется больше кислорода и ДК становится меньше единицы. Так, при использовании жиров ДК = 0, 7.
 Определение дыхательных коэффициентов разных тканей растений показывает, что в нормальных условиях он близок к единице. Это дает основание считать, что в первую очередь растение использует в качестве дыхательного материала углеводы. При недостатке углеводов могут быть использованы и другие субстраты. Особенно это проявляется на проростках, развивающихся из семян, в которых в качестве запасного питательного вещества содержатся жиры или белки. В этом случае дыхательный коэффициент становится меньше единицы.
Определение дыхательных коэффициентов разных тканей растений показывает, что в нормальных условиях он близок к единице. Это дает основание считать, что в первую очередь растение использует в качестве дыхательного материала углеводы. При недостатке углеводов могут быть использованы и другие субстраты. Особенно это проявляется на проростках, развивающихся из семян, в которых в качестве запасного питательного вещества содержатся жиры или белки. В этом случае дыхательный коэффициент становится меньше единицы.
 При использовании в качестве дыхательного материала жиров происходит их расщепление до глицерина и жирных кислот. Жирные кислоты могут быть превращены в углеводы через глиоксилатный цикл. Использованию белков в качестве субстрата дыхания предшествует их расщепление до аминокислот
При использовании в качестве дыхательного материала жиров происходит их расщепление до глицерина и жирных кислот. Жирные кислоты могут быть превращены в углеводы через глиоксилатный цикл. Использованию белков в качестве субстрата дыхания предшествует их расщепление до аминокислот
 Пути дыхательного обмена Существуют две основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов: 1) гликолиз + цикл Кребса (гликолитический); 2) пентозофосфатный (апотомический). Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от факторов среды. .
Пути дыхательного обмена Существуют две основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов: 1) гликолиз + цикл Кребса (гликолитический); 2) пентозофосфатный (апотомический). Относительная роль этих путей дыхания может меняться в зависимости от типа растений, возраста, фазы развития, а также в зависимости от факторов среды. .
 Процесс дыхания растений осуществляется во всех внешних условиях. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от 50 до +50°С. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растений.
Процесс дыхания растений осуществляется во всех внешних условиях. Растительный организм не имеет приспособлений к регуляции температуры, поэтому процесс дыхания осуществляется при температуре от 50 до +50°С. Нет приспособлений у растений и к поддержанию равномерного распределения кислорода по всем тканям. Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в процессе эволюции разнообразных путей дыхательного обмена и к еще большему разнообразию ферментных систем, осуществляющих отдельные этапы дыхания. При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена приводит к глубоким изменениям во всем метаболизме растений.
 Гликолитический путь дыхательного обмена Данный путь дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая фаза — анаэробная (гликолиз), вторая фаза — аэробная. Эти фазы локализованы в различных компартментах клетки. Анаэробная фаза гликолиз — в цитоплазме, аэробная фаза — в митохондриях.
Гликолитический путь дыхательного обмена Данный путь дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая фаза — анаэробная (гликолиз), вторая фаза — аэробная. Эти фазы локализованы в различных компартментах клетки. Анаэробная фаза гликолиз — в цитоплазме, аэробная фаза — в митохондриях.
 Обычно химизм дыхания начинают рассматривать с глюкозы. Вместе с тем в растительных клетках глюкозы мало, поскольку конечными продуктами фотосинтеза являются сахароза как основная транспортная форма сахара в растении или запасные углеводы (крахмал и др. ). Поэтому, чтобы стать субстратом дыхания сахароза и крахмал должны гидролизоваться с образованием глюкозы.
Обычно химизм дыхания начинают рассматривать с глюкозы. Вместе с тем в растительных клетках глюкозы мало, поскольку конечными продуктами фотосинтеза являются сахароза как основная транспортная форма сахара в растении или запасные углеводы (крахмал и др. ). Поэтому, чтобы стать субстратом дыхания сахароза и крахмал должны гидролизоваться с образованием глюкозы.
 Гликолиз осуществляется во всех живых клетках организмов. В процессе гликолиза") Анаэробная фаза дыхания (гликолиз) Гликолиз осуществляется во всех живых клетках организмов. В процессе гликолиза происходит преобразование молекулы гексозы до двух молекул пировиноградной кислоты: С 6 Н 1206 -> 2 С 3 Н 402 + 2 Н 2. Этот окислительный процесс может протекать в анаэробных условиях и идет через ряд этапов. Прежде всего, для того чтобы подвергнуться дыхательному распаду, глюкоза должна быть активирована.
Анаэробная фаза дыхания (гликолиз) Гликолиз осуществляется во всех живых клетках организмов. В процессе гликолиза происходит преобразование молекулы гексозы до двух молекул пировиноградной кислоты: С 6 Н 1206 -> 2 С 3 Н 402 + 2 Н 2. Этот окислительный процесс может протекать в анаэробных условиях и идет через ряд этапов. Прежде всего, для того чтобы подвергнуться дыхательному распаду, глюкоза должна быть активирована.
 Активация глюкозы происходит путем фосфорилирования шестого углеродного атома за счет взаимодействия с АТФ: глюкоза + АТФ -> глюкозо-6 -фосфат + АДФ Реакция идет в присутствии ионов магния и фермента гексокиназа. Затем глюкозо 6 фосфат изомеризуется до фруктозо 6 фосфата. Процесс катализи руется ферментом фосфоглюкоизомеразой: глюкозо-6 -фосфат —> фруктозо-6 -фосфат Далее происходит еще одно фосфорилирование при участии АТФ. Фосфор ная кислота присоединяется к первому углеродному атому молекулы фруктозы, процесс катализируется ферментом фосфофруктокиназой: фруктозо-6 -фосфат + АТФ -> фруктозо-1, 6 дифосфат + АДФ
Активация глюкозы происходит путем фосфорилирования шестого углеродного атома за счет взаимодействия с АТФ: глюкоза + АТФ -> глюкозо-6 -фосфат + АДФ Реакция идет в присутствии ионов магния и фермента гексокиназа. Затем глюкозо 6 фосфат изомеризуется до фруктозо 6 фосфата. Процесс катализи руется ферментом фосфоглюкоизомеразой: глюкозо-6 -фосфат —> фруктозо-6 -фосфат Далее происходит еще одно фосфорилирование при участии АТФ. Фосфор ная кислота присоединяется к первому углеродному атому молекулы фруктозы, процесс катализируется ферментом фосфофруктокиназой: фруктозо-6 -фосфат + АТФ -> фруктозо-1, 6 дифосфат + АДФ
 Поскольку при распаде одной молекулы глюкозы образуются две молекулы ФГА, то все реакции повторяются дважды. В результате процесса гликолиза образуются четыре молекулы АТФ, однако две из них покрывают расход на первоначальное активирование субстрата. Следовательно, накапливаются две молекулы АТФ. Образование АТФ в процессе следующее:
Поскольку при распаде одной молекулы глюкозы образуются две молекулы ФГА, то все реакции повторяются дважды. В результате процесса гликолиза образуются четыре молекулы АТФ, однако две из них покрывают расход на первоначальное активирование субстрата. Следовательно, накапливаются две молекулы АТФ. Образование АТФ в процессе следующее:
 Реакция гликолиза носит название субстратного фосфорилирования, поскольку макроэргические связи возникают на молекуле окисляемого субстрата. Если считать, что при распаде АТФ из АДФ и Фн выделяется 30, 6 к. Дж, то за период гликолиза накапливается в макроэргических фосфатных связях всего 61, 2 к. Дж. Прямые определения показывают, что распад молекулы глюкозы до пировиноградной кислоты сопровождается выделением 586, 6 к. Дж. Следовательно, энергетическая эффективность гликолиза невелика. Кроме того, образуются 2 молекулы НАДН, которые вступают в дыхательную цепь, что приводит к дополнительному образованию АТФ. Образовавшиеся две молекулы пировиноградной кислоты участвуют в аэробной фазе дыхания
Реакция гликолиза носит название субстратного фосфорилирования, поскольку макроэргические связи возникают на молекуле окисляемого субстрата. Если считать, что при распаде АТФ из АДФ и Фн выделяется 30, 6 к. Дж, то за период гликолиза накапливается в макроэргических фосфатных связях всего 61, 2 к. Дж. Прямые определения показывают, что распад молекулы глюкозы до пировиноградной кислоты сопровождается выделением 586, 6 к. Дж. Следовательно, энергетическая эффективность гликолиза невелика. Кроме того, образуются 2 молекулы НАДН, которые вступают в дыхательную цепь, что приводит к дополнительному образованию АТФ. Образовавшиеся две молекулы пировиноградной кислоты участвуют в аэробной фазе дыхания
 Схема гликолиза
Схема гликолиза
 Субстратное фосфорилировае Первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1, 3 -дифосфоглицерат). Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
Субстратное фосфорилировае Первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1, 3 -дифосфоглицерат). Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
 Второе субстратное фосфолирирование Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно
Второе субстратное фосфолирирование Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно
 Суммарное уравнение гликолиза С 6 Н 12 О 6 + 2 НАД* + 2 АДФ+2 Н 3 РО 4 = глюкоза =2 Сн 3 СОСООН + 2 НАДН + 2 АТФ пируват
Суммарное уравнение гликолиза С 6 Н 12 О 6 + 2 НАД* + 2 АДФ+2 Н 3 РО 4 = глюкоза =2 Сн 3 СОСООН + 2 НАДН + 2 АТФ пируват
 Окончательная судьба пирувата и НАД∙H, образованных в процессе гликолиза зависит от организма и условий внутри клетки, в особенности от наличия или отсутствия кислорода или других акцепторов электронов. У анаэробных организмов пируват и НАД∙H далее подвергаются брожению. При молочнокислом брожении, например, у бактерий пируват под действием фермента лактатдегидрогеназы восстанавливается в молочную кислоту. У дрожжей сходным процессом является спиртовое брожение, где конечными продуктами будут этанол и углекислый газ. Известно также маслянокислое и лимоннокислое брожение.
Окончательная судьба пирувата и НАД∙H, образованных в процессе гликолиза зависит от организма и условий внутри клетки, в особенности от наличия или отсутствия кислорода или других акцепторов электронов. У анаэробных организмов пируват и НАД∙H далее подвергаются брожению. При молочнокислом брожении, например, у бактерий пируват под действием фермента лактатдегидрогеназы восстанавливается в молочную кислоту. У дрожжей сходным процессом является спиртовое брожение, где конечными продуктами будут этанол и углекислый газ. Известно также маслянокислое и лимоннокислое брожение.
 Маслянокислое брожение глюкоза → масляная кислота + 2 CO 2 + 2 H 2 O. Спиртовое брожение: глюкоза → 2 этанол + 2 CO 2. Лимоннокислое брожение: глюкоза → лимонная кислота + 2 H 2 O. Брожение имеет важное значение в пищевой промышленности. У аэробов пируват как правило попадает в цикл трикарбоновых кислот (цикл Кребса), а НАД∙H в итоге окисляется кислородом на дыхательной цепи в митохондриях в процессе окислительного фосфорилирования.
Маслянокислое брожение глюкоза → масляная кислота + 2 CO 2 + 2 H 2 O. Спиртовое брожение: глюкоза → 2 этанол + 2 CO 2. Лимоннокислое брожение: глюкоза → лимонная кислота + 2 H 2 O. Брожение имеет важное значение в пищевой промышленности. У аэробов пируват как правило попадает в цикл трикарбоновых кислот (цикл Кребса), а НАД∙H в итоге окисляется кислородом на дыхательной цепи в митохондриях в процессе окислительного фосфорилирования.
 Несмотря на то, что метаболизм человека преимущественно аэробный, в интенсивно работающих скелетных мышцах наблюдается анаэробное окисление. В условиях ограниченного доступа кислорода пируват превращается в молочную кислоту, как происходит при молочнокислом брожении у многих микроорганизмов: ПВК + НАД∙Н + H+ → лактат + НАД+ Боли в мышцах, возникающие через некоторое время после непривычной интенсивной физической нагрузки, связаны с накоплением в них молочной кислоты. Образование молочной кислоты является тупиковой ветвью метаболизма, но не является конечным продуктом обмена веществ. Под действием лактатдегидрогеназы молочная кислота окисляется снова, образуя пируват, который и участвует в дальнейших превращениях.
Несмотря на то, что метаболизм человека преимущественно аэробный, в интенсивно работающих скелетных мышцах наблюдается анаэробное окисление. В условиях ограниченного доступа кислорода пируват превращается в молочную кислоту, как происходит при молочнокислом брожении у многих микроорганизмов: ПВК + НАД∙Н + H+ → лактат + НАД+ Боли в мышцах, возникающие через некоторое время после непривычной интенсивной физической нагрузки, связаны с накоплением в них молочной кислоты. Образование молочной кислоты является тупиковой ветвью метаболизма, но не является конечным продуктом обмена веществ. Под действием лактатдегидрогеназы молочная кислота окисляется снова, образуя пируват, который и участвует в дальнейших превращениях.
 Вторая фаза дыхания — аэробная локализована в митохондриях и требует присутствия кислорода. В аэробную фазу дыхания вступает пировиноградная ки слота. Общее уравнение этого процесса следующее: 2 ПВК + 502 + 6 Н 20 -> 6 С 02 + 5 Н 20
Вторая фаза дыхания — аэробная локализована в митохондриях и требует присутствия кислорода. В аэробную фазу дыхания вступает пировиноградная ки слота. Общее уравнение этого процесса следующее: 2 ПВК + 502 + 6 Н 20 -> 6 С 02 + 5 Н 20
 окислительное декарбоксилирование пировиноградной кислоты; 2)") Процесс можно разделить на три основные стадии: 1) окислительное декарбоксилирование пировиноградной кислоты; 2) цикл трикарбоновых кислот (цикл Кребса); 3) заключительная стадия окисления — электронтранспортная цепь (ЭТЦ) требует обязательного присутствия 02. Первые две стадии происходят в матриксе митохондрий, электронтранспортная цепь локализована на внутренней мембране митохондрий.
Процесс можно разделить на три основные стадии: 1) окислительное декарбоксилирование пировиноградной кислоты; 2) цикл трикарбоновых кислот (цикл Кребса); 3) заключительная стадия окисления — электронтранспортная цепь (ЭТЦ) требует обязательного присутствия 02. Первые две стадии происходят в матриксе митохондрий, электронтранспортная цепь локализована на внутренней мембране митохондрий.
 Первая стадия — окислительное декарбоксилирование пировиноградной кисло ты. СН 3 СОСООН + НАД + Ко. А - SH -> CH 3 CO-S- Ко. А + НАДН + Н+ + С 02 Процесс этот состоит из ряда реакций и катализируется сложной мультиферментной системой пируватдекарбоксилазой. Пируватдекарбоксилаза включает в себя три фермента и пять коферментов (тиаминпирофосфат, липоевая кисло та, коэнзим А — Ko. A SH, ФАД и НАД). В результате этого процесса образуется активный ацетат — ацетилкоэнзим А (ацетил Ко. А), восстановленный НАД (НАДН + Н+), и выделяет ся углекислый газ (первая молекула). Восстановленный НАД поступает в цепь переноса электронов, а ацетил Ко. А вступает в цикл трикарбоновых кислот. Важ но отметить, что пируватдегидрогеназная система ингибируется АТФ. При накоплении АТФ выше определенного уровня превращение пировиноградной кислоты подавляется. Это один из способов регуляции интенсивности протека ния аэробной фазы.
Первая стадия — окислительное декарбоксилирование пировиноградной кисло ты. СН 3 СОСООН + НАД + Ко. А - SH -> CH 3 CO-S- Ко. А + НАДН + Н+ + С 02 Процесс этот состоит из ряда реакций и катализируется сложной мультиферментной системой пируватдекарбоксилазой. Пируватдекарбоксилаза включает в себя три фермента и пять коферментов (тиаминпирофосфат, липоевая кисло та, коэнзим А — Ko. A SH, ФАД и НАД). В результате этого процесса образуется активный ацетат — ацетилкоэнзим А (ацетил Ко. А), восстановленный НАД (НАДН + Н+), и выделяет ся углекислый газ (первая молекула). Восстановленный НАД поступает в цепь переноса электронов, а ацетил Ко. А вступает в цикл трикарбоновых кислот. Важ но отметить, что пируватдегидрогеназная система ингибируется АТФ. При накоплении АТФ выше определенного уровня превращение пировиноградной кислоты подавляется. Это один из способов регуляции интенсивности протека ния аэробной фазы.
,") В результате этого процесса образуется активный ацетат — ацетилкоэнзим А (ацетил Ко. А), восстановленный НАД (НАДН + Н+), и выделяется углекислый газ (первая молекула). Восстановленный НАД поступает в цепь переноса электронов, а ацетил Ко. А вступает в цикл трикарбоновых кислот. пируватдегидрогеназная система ингибируется АТФ. При накоплении АТФ выше определенного уровня превращение ПВК подавляется. Это один из способов регуляции интенсивности протекания аэробной фазы.
В результате этого процесса образуется активный ацетат — ацетилкоэнзим А (ацетил Ко. А), восстановленный НАД (НАДН + Н+), и выделяется углекислый газ (первая молекула). Восстановленный НАД поступает в цепь переноса электронов, а ацетил Ко. А вступает в цикл трикарбоновых кислот. пируватдегидрогеназная система ингибируется АТФ. При накоплении АТФ выше определенного уровня превращение ПВК подавляется. Это один из способов регуляции интенсивности протекания аэробной фазы.
. В 1935 г. венгерский ученый А.") Вторая стадия — цикл трикарбоновых кислот (цикл Кребса). В 1935 г. венгерский ученый А. Сент Дьердьи установил, что добавление небольших количеств органических кислот (фумаровой, яблочной или янтарной) усиливает поглощение кислорода измельченными тканями. Продолжая эти исследования, Г. Кребс пришел к выводу, что главным путем окисления углеводов являются циклические реакции, в которых происходит постепенное преобразование ряда органических кислот. Эти преобразования и были названы циклом трикарбоновых кислот или циклом Кребса. Сам исследователь за эти работы в 1953 г. был удостоен Нобелевской премии.
Вторая стадия — цикл трикарбоновых кислот (цикл Кребса). В 1935 г. венгерский ученый А. Сент Дьердьи установил, что добавление небольших количеств органических кислот (фумаровой, яблочной или янтарной) усиливает поглощение кислорода измельченными тканями. Продолжая эти исследования, Г. Кребс пришел к выводу, что главным путем окисления углеводов являются циклические реакции, в которых происходит постепенное преобразование ряда органических кислот. Эти преобразования и были названы циклом трикарбоновых кислот или циклом Кребса. Сам исследователь за эти работы в 1953 г. был удостоен Нобелевской премии.
 В цикл вступает активный ацетат, или ацетил. Ко. А. ацетил Ко. А конденсируется с щавелевоуксусной кислотой (ЩУК). Далее превращение идет через ряд ди и трикарбоновых органических кислот. В результате ЩУК регенерирует в прежнем виде. В процессе цикла присоединяются три молекулы Н 20, выделяются две молекулы С 02 и четыре пары водорода, которые восстанавливают соответствующие коферменты (ФАД и НАД). Суммарная реакция цикла выражена уравнением:
В цикл вступает активный ацетат, или ацетил. Ко. А. ацетил Ко. А конденсируется с щавелевоуксусной кислотой (ЩУК). Далее превращение идет через ряд ди и трикарбоновых органических кислот. В результате ЩУК регенерирует в прежнем виде. В процессе цикла присоединяются три молекулы Н 20, выделяются две молекулы С 02 и четыре пары водорода, которые восстанавливают соответствующие коферменты (ФАД и НАД). Суммарная реакция цикла выражена уравнением:
 Суммарная реакция цикла Кребса выражена уравнением: CH 3 CO-S-K 0 A + ЗН 20 + ЗНАД + ФАД + АДФ + Фн -> 2 С 02 + SH-Ko. A + ЗНАДН + ЗН+ + ФАДН 2 + АТФ
Суммарная реакция цикла Кребса выражена уравнением: CH 3 CO-S-K 0 A + ЗН 20 + ЗНАД + ФАД + АДФ + Фн -> 2 С 02 + SH-Ko. A + ЗНАДН + ЗН+ + ФАДН 2 + АТФ
 2.
2.
 образовавшиеся органические кислоты могут служить материалом для построения аминокислот, жиров и углеводов. В этом случае они выводятся из цикла. Однако соединения, входящие в цикл, могут образовываться в ряде других реакций (например, при декарбоксилировании амино кислот) и вступать в цикл. Таким образом, рассмотренные превращения не отделены от других реакций метаболизма, а тесно с ними взаимосвязаны.
образовавшиеся органические кислоты могут служить материалом для построения аминокислот, жиров и углеводов. В этом случае они выводятся из цикла. Однако соединения, входящие в цикл, могут образовываться в ряде других реакций (например, при декарбоксилировании амино кислот) и вступать в цикл. Таким образом, рассмотренные превращения не отделены от других реакций метаболизма, а тесно с ними взаимосвязаны.
 Для реакций цикла Кребса кислород не требуется. Кислород необходим для регенерации или окисления восстановленных коферментов (НАДН + Н+ и ФАДН 2). Количество окисленных форм этих коферментов ограничено. В анаэробных условиях, когда регенерация коферментов невозможна, они быстро оказываются исчерпанными и весь процесс прекращается. Окисление коферментов осуществляется в дыхательной цепи, или цепи переноса водорода и электронов. Конечным акцептором в этой цепи является кислород воздуха. Энергия, высвобождаемая при окислении коферментов, накапливается в макроэргических фосфатных связях АТФ.
Для реакций цикла Кребса кислород не требуется. Кислород необходим для регенерации или окисления восстановленных коферментов (НАДН + Н+ и ФАДН 2). Количество окисленных форм этих коферментов ограничено. В анаэробных условиях, когда регенерация коферментов невозможна, они быстро оказываются исчерпанными и весь процесс прекращается. Окисление коферментов осуществляется в дыхательной цепи, или цепи переноса водорода и электронов. Конечным акцептором в этой цепи является кислород воздуха. Энергия, высвобождаемая при окислении коферментов, накапливается в макроэргических фосфатных связях АТФ.
 в результате распада 1 молекулы ПВК в аэробной фазе (декарбоксилирование ПВК + цикл Кребса) выделяется ЗС 02, 4 молекулы НАДН + Н+ и 1 молекула ФАДН 2. Таким образом, 5 пар Н 2, образующихся из ПВК и воды, поступают в дыхательную цепь.
в результате распада 1 молекулы ПВК в аэробной фазе (декарбоксилирование ПВК + цикл Кребса) выделяется ЗС 02, 4 молекулы НАДН + Н+ и 1 молекула ФАДН 2. Таким образом, 5 пар Н 2, образующихся из ПВК и воды, поступают в дыхательную цепь.
 Пентозофосфатный путь дыхательного обмена Наряду с разложением глюкозы в процессе гликолиза существует другой, не менее распространенный путь — пентозофосфатный, или апотомический. В этом случае глюкоза также подвергается первоначальному фосфорилированию с образованием глюкозо-6 -фосфата, затем пути расходятся. Монофосфорный эфир глюкозы подвергается окислению при участии фермента глюкозо 6 фосфатдегидрогеназы. Коферментом является НАДФ, при этом образуется фосфоглюконовая кислота
Пентозофосфатный путь дыхательного обмена Наряду с разложением глюкозы в процессе гликолиза существует другой, не менее распространенный путь — пентозофосфатный, или апотомический. В этом случае глюкоза также подвергается первоначальному фосфорилированию с образованием глюкозо-6 -фосфата, затем пути расходятся. Монофосфорный эфир глюкозы подвергается окислению при участии фермента глюкозо 6 фосфатдегидрогеназы. Коферментом является НАДФ, при этом образуется фосфоглюконовая кислота
 Суммарное уравнение пентозофосфатного цикла 6 Глюкозо-6 -фосфат + 12 НАДФ 5 Глюкозо-6 фос-фат+6 CO 2 = 12 НАДФН +12 H+ + Н 3 РО 4
Суммарное уравнение пентозофосфатного цикла 6 Глюкозо-6 -фосфат + 12 НАДФ 5 Глюкозо-6 фос-фат+6 CO 2 = 12 НАДФН +12 H+ + Н 3 РО 4
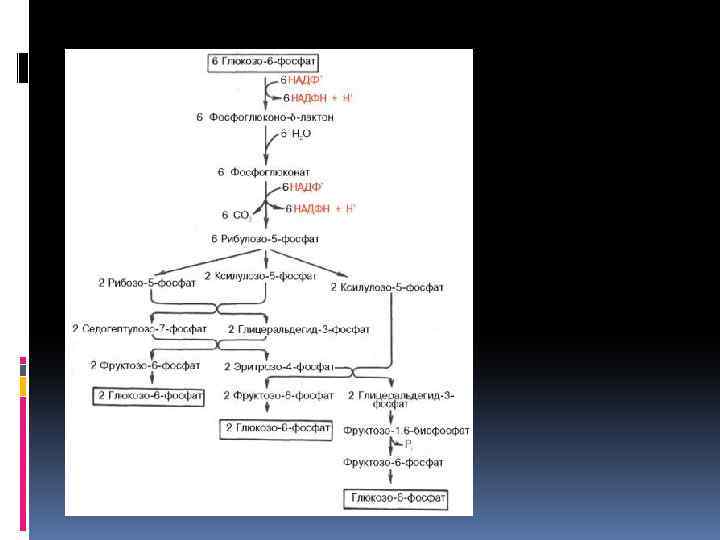
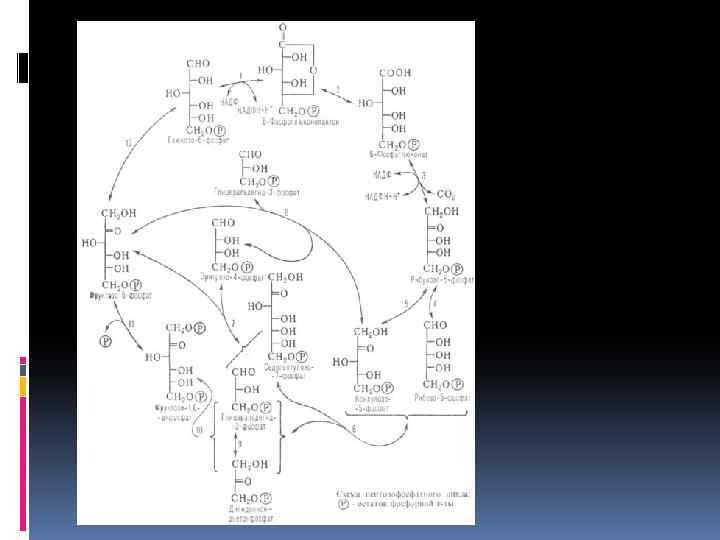
 Реакции окисления глюкозы, в пентозофосфатном пути, показывают его большое сходство с превращением углеводов в фотосинтетическом цикле Кальвина. Возможно, что благодаря наличию одинаковых промежуточных продуктов эти процессы в ряде пунктов взаимосвязаны. Наряду с локализацией пентозофосфатного пути в цитоплазме он может в отсутствие света осуществляться и в хлороплатах. На протяжении онтогенеза растений и в зависимости от условий происходит смена дыхательных путей. В силу образования различных промежуточных продуктов это оказывает значительное влияние на обмен веществ и физиологическую деятельность растительного организма.
Реакции окисления глюкозы, в пентозофосфатном пути, показывают его большое сходство с превращением углеводов в фотосинтетическом цикле Кальвина. Возможно, что благодаря наличию одинаковых промежуточных продуктов эти процессы в ряде пунктов взаимосвязаны. Наряду с локализацией пентозофосфатного пути в цитоплазме он может в отсутствие света осуществляться и в хлороплатах. На протяжении онтогенеза растений и в зависимости от условий происходит смена дыхательных путей. В силу образования различных промежуточных продуктов это оказывает значительное влияние на обмен веществ и физиологическую деятельность растительного организма.

