Дыхание и водно-солевой обмен гидробионтов.pptx
- Количество слайдов: 19



ДЫХАНИЕ И ВОДНОСОЛЕВОЙ ОБМЕН ГИДРОБИОНТОВ

Особенности дыхания водных животных обусловлены различиями в физических свойствах воды и воздуха как среды дыхания. Плотность воды выше плотности воздуха примерно в 780 раз. Вязкость воды больше вязкости воздуха в 55 раз. Величина коэффициента диффузии кислорода в воде меньше, чем в воздухе, в 10 000 раз! Растворимость кислорода в воде невелика: при 15°С и давлении сухого газа над водной поверхностью в 1 атм (101, 3 к. Па) в 1 л воды растворяется около 34 мл O 2 Практически содержание кислорода в природных водоёмах существенно ниже и редко превышает 10 -11 мл/л. Суммарное воздействие многих и разнообразных факторов на процесс растворения кислорода приводит к тому, что его содержание в естественных водоёмах не только невелико, но и весьма изменчиво, что создаёт сложную экологическую обстановку для реализации процессов газообмену гидробионтов.

Перенос кислорода в воде осуществляется путем конвективной диффузии, но в тонком слое воды на границе раздела твердое тело – жидкость, где скорость жидкости равна нулю, основное значение имеет стационарная диффузия, сильно ограничивающая скорость переноса кислорода из воды в тело животных. Внутри клеток животных, где конвективные движения жидкости зачастую сильно ограничены, перенос кислорода происходит преимущественно за счет стационарной диффузии. Коэффициент диффузии кислорода в клетках и тканях в 1, 5 -3, 0 раза ниже, чем в воде. Размер шарообразного одноклеточного организма, лишенного специальных систем подачи кислорода, теоретически ограничен диаметром около 1 мм.

Переход кислорода из внешней среды во внутреннюю через дыхательную мембрану жабр совершается за счет чисто диффузного процесса, и для того, чтобы этот процесс был достаточно эффективным, пограничная дыхательная поверхность должна быть очень большой. Большая сеть капилляров в жабрах и альвеолах создает большую диффузионную поверхность. Поступивший в кровь кислород связывается гемоглобином и переносится с потоком крови к тканям. Переход кислорода из крови в ткани происходит почти исключительно за счет простой диффузии из множества мелких капилляров. Если в воздушной среде каждый литр попадающего в легкие воздуха приносит 210 мл кислорода, то для того, чтобы принести к жабрам такое же количество кислорода, водному животному необходимо прокачать через жабры около 33 литров воды. В первом случае масса прокачиваемого через легкие кислорода составит около 0, 6 г, а эквивалентный ему по содержанию кислорода объем воды имеет массу 33 000 г. Подавляющее большинство мелких гидробионтов дышат всей поверхностью тела и специальных органов дыхания, как правило, не имеют.

Основные условия для осуществления дыхания Большая удельная дыхательная поверхность Образующий слой не больше 1 мм Мелкие беспозвоночные животные, живущие в толще воды и на дне, усваивают О 2 и выделяют СО 2 через покровы тела, личинки насекомых – посредством мальпигиевых сосудов, как правило, открытых, т. е. соединяющихся с водной средой. У гидробионтов, живущие в условиях дефицита О 2, в гемолимфе содержится пигмент, связывающий кислород воды, и, тем самым, увеличивающий эффективность дыхания,

Многие беспозвоночные гидробионты усваивают кислород из воды плохо, в связи с чем вынуждены подниматься к поверхности воды, чтобы «глотнуть» О 2 из атмосферы.

У эмбрионов рыб на ранних стадиях развития специализированных органов дыхания нет и кислород поглощается всей поверхностью тела зародыша. Личинки рыб имеют органы дыхания, которыми являются сосуд желточного мешка, кювьеровы протоки и др. сосуды

Жаберное дыхание рыб У рыб функцию газообмена выполняют жабры. На каждой жаберной дуге расположено несколько пар жаберных лепестков, внутри которых проходят мельчайшие кровеносные сосуды. Рыбы при помощи мышц способны раздвигать и смыкать жаберные лепестки синхронно с движением жаберных крышек, и даже встряхивать жаберные лепестки, очищая их таким образом от налипших на них частиц.
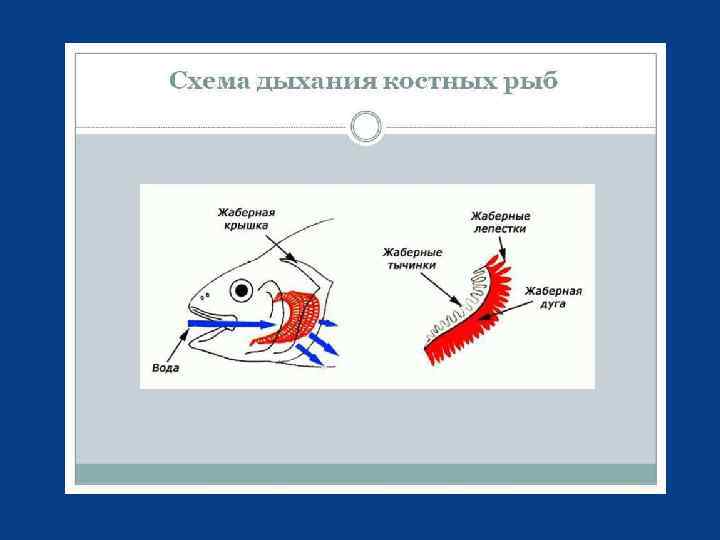
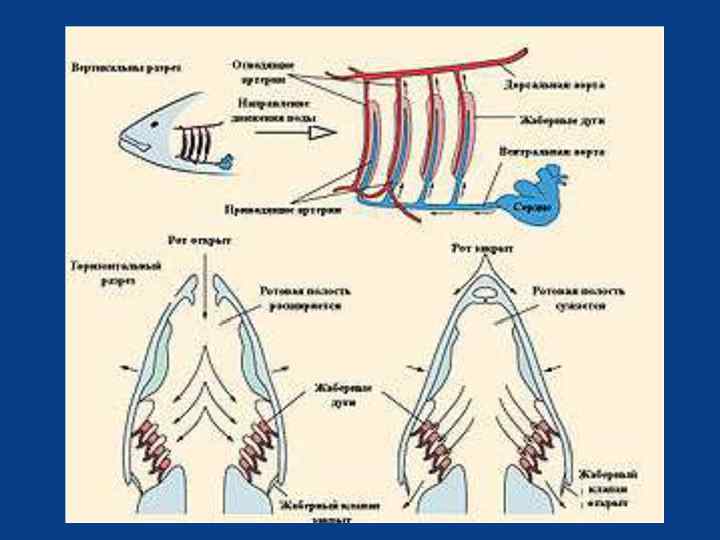

Водно-солевой обмен По характеру водно-солевого обмена гидробионты делятся на пресноводных и морских, хотя некоторые эвригалинные (приспособленные к широкому диапазону солевых концентраций) формы могут обитать и в тех, и в других условиях. Для всех первичноводных организмов характерно наличие проницаемых для воды покровов, поэтому различие осмотической концентрации водной среды и жидкостей организма создаёт осмотический ток воды в сторону большего осмотического давления. Результат осмотических процессов неодинаков для обитателей разных типов водоёмов.

У большинства обитателей морских водоёмов концентрация солей в организме близка к таковой окружающей среды, а благодаря проницаемости покровов любые изменения солёности немедленно уравновешиваются осмотическим током воды. Такие организмы принято называть пдйкилоосмотическими. Таковы практически все цианобактерии и низшие растения, а также большинство морских беспозвоночных животных; последних часто называют осмоконформерами.

Животных, способных к активной регуляции осмотического давления жидкостей тела, поддерживающих относительное постоянство этого параметра внутренней среды независимо от окружающей воды, называют гомойоосмотическими, или авторегуляторами. Существует небольшая группа изотоничных организмов (кишечнополостные, иглокожие). Осмотическое давление жидкостей их тела близко к таковому морской воды и изменяется параллельно изменениям внешней солёности. У большинства других беспозвоночных регистрируется некоторое превышение осмотического давления внутренней среды организма (его гипертоничность), что обеспечивает постоянный приток в организм воды в пределах, легко уравновешивающихся процессами выделения.

Способность изотоничных животных переносить некоторые изменения солёности среды определяются главным образом механизмами клеточной устойчивости к обводнению или дегидратации. Диапазон такой устойчивости обычно не очень велик, поэтому изоосмотические осмоконформеры распространены, как правило, в морских водоёмах с относительно устойчивой солёностью. Беспозвоночным осморегуляторам (высшие раки, моллюски, насекомые и др. ) свойственно переносить более значительные колебания солёности. Это обеспечивается механизмами активной регуляции осмотического давления внутренней среды, которые включают изменения проницаемости мембран, активный перенос ионов (в включая выведение избытка солей), а также изменения внутриклеточной концентрации свободных аминокислот в направлении, уравновешивающем суммарное осмотическое давление в клетке с внешней средой. У пойкилоосмотических организмов имеется возможность осуществления активной ионной регуляции, которая определяет отличия количественных показателей содержания отдельных ионов в среде и в организме. Это основа жизнедеятельности организмов концентраторов, способных избирательно извлекать из среды и накапливать в организме отдельные соли. В отличие от осморегуляции ионная регуляция свойственна большинству живых организмов.

Пресноводные жизненные формы всегда гипертоничны, в силу чего внутрь организма направлен постоянный осмотический поток воды. Они относятся к гомойоосмотическим формам. Наиболее эффективная адаптация позвоночных к обитанию в гипотонической среде - образование клубочковых почек, действие которых основано на принципах избирательной ультрафильтрации и реабсорбции. Продукты азотистого обмена аммиак и мочевина - растворимы в воде и легко выводятся не только через почки, но и через жабры.

Морские формы существуют в условиях гипертоничности внешней среды, обусловливающей постоянные осмотические потери воды (в основном через жабры). Пополнение потерь воды у них происходит путём питья. Избыток получаемых при этом солей выводится через почки и с фекалиями (главным образом двухвалентные ионы), а также активно экскретируются особыми клетками жаберного эпителия (преимущественно Na+ C 1+).

Тоничность – концентрация солей в жидкости клеток и тканей гидробионтов. Тоничность измеряется в градусах температуры замерзания водного раствора. Тоничность морской воды – минус 1, 5 -2, 0 градуса Тоничность …морских рыб – минус 0, 7 -0, 8 градуса Тоничность…пресноводных рыб - минус 0, 5 -0, 6 градуса Тоничность…морских хрящевых рыб – минус 2 и более градуса Хрящевые рыбы, как и живущие в пресных водах, воду пьют Причины – высокое содержание в крови и жидкостяххрящевых рыб МОЧЕВИНЫ.

ИОНИЧНОСТЬ – соотношение ионов солей в организме. Регулируется в организме гидробионтов гораздо строже, чем тоничность. Изотоничность – явление частое, изооничность –редкое. Например, в организме многих морских животных больше, чем в морской воде, калия, но меньше магния, натрия, сульфатов. Все гидробионты (в т. ч. морские беспозвоночные) осуществляют АКТИВНОЕ РЕГУЛИРОВАНИЕ СОЛЕВОГО СОСТАВА ОРГАНИЗМА, но в пределах своих возможностей ( «нормы реакции» генотипа). Хорошо изучен КАЛИЕВО-НАТРИЕВЫЙ НАСОС: натрий внутрь организма, калий (и аммоний – NH 4) - наружу. Известны и другие «НАСОСЫ» .

Активный транспорт ионов солей внутрь и наружу осуществляется только в специализированных клетках кожи, кишечника, жабр, мочевых протоков. В результате того, что ионная регуляция у гидробионтов далека от совершенства, коэффициенты концентрации химических элементов в их организме достигают сотен, тысяч и десятков тысяч единиц. Это касается и водных растений. Отсюда проблемы в связи с загрязнением водоемов!!!
Дыхание и водно-солевой обмен гидробионтов.pptx