DNK_lektsia.ppt
- Количество слайдов: 54
 ДНК Строение и организация наследственного материала в клетке
ДНК Строение и организация наследственного материала в клетке
 Одна молекула Днк – соответствует одной хромосоме
Одна молекула Днк – соответствует одной хромосоме
 ДНК l l l Комплекс ДНК со специальными белками называется хроматином. Компактизированный хроматин – гетерохроматин Декомпактизированный - эухроматин
ДНК l l l Комплекс ДНК со специальными белками называется хроматином. Компактизированный хроматин – гетерохроматин Декомпактизированный - эухроматин
 Нуклеосомный уровень организации наследственного материала Нуклеосомы
Нуклеосомный уровень организации наследственного материала Нуклеосомы
 Нуклеосомный уровень организации наследственного материала l l Основа нуклеосомы – глобула из 8 белков 4 –х типов (Н 2 А, Н 2 В, Н 3, Н 4) 2 обороты = 140 н. п.
Нуклеосомный уровень организации наследственного материала l l Основа нуклеосомы – глобула из 8 белков 4 –х типов (Н 2 А, Н 2 В, Н 3, Н 4) 2 обороты = 140 н. п.
 Нуклеосомы закручиваются в спираль – нить укорачивается в 40 раз
Нуклеосомы закручиваются в спираль – нить укорачивается в 40 раз
 Сверхспирализация В гетерохроматине к нуклеосомному уровню добавляются следующие уровни компактизации: 1. Нуклеосомная нить укладывается в спираль 2. Хроматиновые нити образуют петли, которые укладываются в розетки
Сверхспирализация В гетерохроматине к нуклеосомному уровню добавляются следующие уровни компактизации: 1. Нуклеосомная нить укладывается в спираль 2. Хроматиновые нити образуют петли, которые укладываются в розетки
 репликативная вилка Распространение репликативных вилок на ДНК
репликативная вилка Распространение репликативных вилок на ДНК
 Современное представление о гене l l l Различают 3 уровня организации наследственного материала: 1. Хромосомный 2. Генный 3. Геномный (уровень генотипа) 4. Элементарная структура генного уровня –ген (материальный носитель генетической информации)
Современное представление о гене l l l Различают 3 уровня организации наследственного материала: 1. Хромосомный 2. Генный 3. Геномный (уровень генотипа) 4. Элементарная структура генного уровня –ген (материальный носитель генетической информации)
 Современное представление о гене l l Гены находятся в определенных хромосомах. Место расположения гена в хромосоме называется –локусом Совокупность всех генов в организме, проявляющихся фенотипически, называется генотипом Геном- вся ДНК, содержащейся в гаплоидном наборе хромосом
Современное представление о гене l l Гены находятся в определенных хромосомах. Место расположения гена в хромосоме называется –локусом Совокупность всех генов в организме, проявляющихся фенотипически, называется генотипом Геном- вся ДНК, содержащейся в гаплоидном наборе хромосом
 у вирусов от") Организация генома у прокариот l l l Небольшой размер генома А) у вирусов от неск. тыс. до неск. сотен нуклеотидных пар (н. п. ) Б)у вирусов кодирующие белок гены расположены один за другим и занимают до 100% длины соответствующей ДНК
Организация генома у прокариот l l l Небольшой размер генома А) у вирусов от неск. тыс. до неск. сотен нуклеотидных пар (н. п. ) Б)у вирусов кодирующие белок гены расположены один за другим и занимают до 100% длины соответствующей ДНК
 размер генома значительно больше") Организация генома у прокариот l l l У бактерий: А) размер генома значительно больше (у кишечной палочки 4, 2*10*6 н. п. ) Б)структурные гены (кодирующие структуру белков) занимают более 50% бактериальной хромосомы В)остальную часть занимают некодирующие последовательности Г)подавляющее большинство бактериальных генов уникально т. е.
Организация генома у прокариот l l l У бактерий: А) размер генома значительно больше (у кишечной палочки 4, 2*10*6 н. п. ) Б)структурные гены (кодирующие структуру белков) занимают более 50% бактериальной хромосомы В)остальную часть занимают некодирующие последовательности Г)подавляющее большинство бактериальных генов уникально т. е.
 исключение – гены транспортных") l l l представлены в геноме только 1 раз. Д) исключение – гены транспортных и рибосомальных РНК, которые повторяются 10 раз. Е) для организации генетического материала прокариот характерны опероны, состоящие из нескольких генов. Они являются единицами транскрипции. Каждый транскрипт (и – РНК) содержит одну нуклеотидную последовательность
l l l представлены в геноме только 1 раз. Д) исключение – гены транспортных и рибосомальных РНК, которые повторяются 10 раз. Е) для организации генетического материала прокариот характерны опероны, состоящие из нескольких генов. Они являются единицами транскрипции. Каждый транскрипт (и – РНК) содержит одну нуклеотидную последовательность
 структурный ген прокариот непрерывен – все триплеты несут информацию о") одного структурного гена Ж) структурный ген прокариот непрерывен – все триплеты несут информацию о структуре белка.
одного структурного гена Ж) структурный ген прокариот непрерывен – все триплеты несут информацию о структуре белка.
 Организация генома у эукариот l l Очень большой по размерам геном ( У человека 5*10*9 н. п. ) Количество структурных генов при этом возрастает не очень сильно т. о. геном эукариот избыточен: а) некоторые гены и нуклеотидные последовательности многократно повторяются б)много генетических элемнтов имеющих регуляторную функцию
Организация генома у эукариот l l Очень большой по размерам геном ( У человека 5*10*9 н. п. ) Количество структурных генов при этом возрастает не очень сильно т. о. геном эукариот избыточен: а) некоторые гены и нуклеотидные последовательности многократно повторяются б)много генетических элемнтов имеющих регуляторную функцию
 часть ДНК не содержит генов – «молчащая ДНК» – у") l l l в) часть ДНК не содержит генов – «молчащая ДНК» – у некоторых видов до 99% Г) гены эукариот « прерывисты» – включают кодирующие участки – экзоны и некодирующие участки интроны д) Характерно присутствие Т – кластеров (сложных генов), кодирующих длинные полипептидные цепи с несколькими ферментными активностями
l l l в) часть ДНК не содержит генов – «молчащая ДНК» – у некоторых видов до 99% Г) гены эукариот « прерывисты» – включают кодирующие участки – экзоны и некодирующие участки интроны д) Характерно присутствие Т – кластеров (сложных генов), кодирующих длинные полипептидные цепи с несколькими ферментными активностями
 Общая черта Перекрывание генов – тенденция максимального использования информационной емкости
Общая черта Перекрывание генов – тенденция максимального использования информационной емкости
 ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА : общие принципы: Функциональные отделы генома. Гены и их структура. Ген – это участок ДНК, кодирующий белок Цистрон – участок ДНК, кодирующий одну полипептидную цепь Такое подразделение ГЕН БЕЛКА ДНК Цистрон А Белок А Цистрон В В относится в основном к бактериям, где цистроны следуют один за другим У эукариот цистроны нередко располагаются в разных хромосомах Кроме генов белков в хромосомах имеются гены РНК
ОРГАНИЗАЦИЯ ГЕНЕТИЧЕСКОГО МАТЕРИАЛА : общие принципы: Функциональные отделы генома. Гены и их структура. Ген – это участок ДНК, кодирующий белок Цистрон – участок ДНК, кодирующий одну полипептидную цепь Такое подразделение ГЕН БЕЛКА ДНК Цистрон А Белок А Цистрон В В относится в основном к бактериям, где цистроны следуют один за другим У эукариот цистроны нередко располагаются в разных хромосомах Кроме генов белков в хромосомах имеются гены РНК
 Общая совокупность генов, определяющих наследственную информацию организма – геном. • Всего в геноме бактерий 2500 цистронов • В хромосомах человека число генов – 30 000 • Гены эукариот содержат не только кодирующие участки экзоны , но и некодирующие - интроны (прерывистое строение генов) интрон экзон интрон цистрон экзон цистрон интрон эукариоты ПРОКАРИОТЫ
Общая совокупность генов, определяющих наследственную информацию организма – геном. • Всего в геноме бактерий 2500 цистронов • В хромосомах человека число генов – 30 000 • Гены эукариот содержат не только кодирующие участки экзоны , но и некодирующие - интроны (прерывистое строение генов) интрон экзон интрон цистрон экзон цистрон интрон эукариоты ПРОКАРИОТЫ
 Ген – единица наследственности, определяющая развитие какого- либо признака организма. Гены находятся в определенных локусах хромосом. Концепция Дж. Бидла и Т. Тейтема (40 -е годы 20 в. ). «один ген – один фермент» ГЕН- и. РНК – синтез белка – биохимическая реакция – признак внешняя среда
Ген – единица наследственности, определяющая развитие какого- либо признака организма. Гены находятся в определенных локусах хромосом. Концепция Дж. Бидла и Т. Тейтема (40 -е годы 20 в. ). «один ген – один фермент» ГЕН- и. РНК – синтез белка – биохимическая реакция – признак внешняя среда
 ПРОЧИЕ ОТДЕЛЫ ДНК НЕКОДИРУЮЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ – СПЕЙСЕРЫ: ВЫПОЛНЯЮТ СТРУКТУРНУЮ РОЛЬ: §УЧАСТВУЮТ В ПРАВИЛЬНОЙ УКЛАДКЕ НУКЛЕОСОМНОЙ ЦЕПИ §В ПРИКРЕПЛЕНИИ ХРОМОСОМ К АППАРАТУ ЦЕНТРИОЛЕЙ СЛУЖАТ СПЕЦИФИЧЕСКИМИ ЛОКУСАМИ СВЯЗЫВАНИЯ ОПРЕДЕЛЕННЫХ БЕЛКОВ: §ФУНКЦИОНИРУЮЩИХ НА ДНК ПОЛИМЕРАЗНОГО КОМПЛЕКСА) ФЕРМЕНТОВ §БЕЛКОВ. ВЫПОЛНЯЮЩИХ РЕГУЛЯТОРНУЮ ФУНКЦИЮ (ДНК-
ПРОЧИЕ ОТДЕЛЫ ДНК НЕКОДИРУЮЩИЕ ПОСЛЕДОВАТЕЛЬНОСТИ – СПЕЙСЕРЫ: ВЫПОЛНЯЮТ СТРУКТУРНУЮ РОЛЬ: §УЧАСТВУЮТ В ПРАВИЛЬНОЙ УКЛАДКЕ НУКЛЕОСОМНОЙ ЦЕПИ §В ПРИКРЕПЛЕНИИ ХРОМОСОМ К АППАРАТУ ЦЕНТРИОЛЕЙ СЛУЖАТ СПЕЦИФИЧЕСКИМИ ЛОКУСАМИ СВЯЗЫВАНИЯ ОПРЕДЕЛЕННЫХ БЕЛКОВ: §ФУНКЦИОНИРУЮЩИХ НА ДНК ПОЛИМЕРАЗНОГО КОМПЛЕКСА) ФЕРМЕНТОВ §БЕЛКОВ. ВЫПОЛНЯЮЩИХ РЕГУЛЯТОРНУЮ ФУНКЦИЮ (ДНК-
 • Участок связывания РНК полимеразы- промотор Бокс Прибнова ТАТА-бокс –область инициации ПРОМОТОР ПРОКАРИОТ ПРОМОТОР ЭУКАРИОТ • УЧАСТОК СВЯЗЫВАНИЯ РЕГУЛЯТОРНЫХ БЕЛКОВ – ОПЕРАТОР. С ОПЕРАТОРОМ СВЯЗЫВАЕТСЯ БЕЛОК – РЕПРЕССОР. Это блокирует прочтение РНК -ПОЛИМЕРАЗОЙ СООТВЕТСТВУЮЩЕЙ ГРУППЫ ГЕНОВ.
• Участок связывания РНК полимеразы- промотор Бокс Прибнова ТАТА-бокс –область инициации ПРОМОТОР ПРОКАРИОТ ПРОМОТОР ЭУКАРИОТ • УЧАСТОК СВЯЗЫВАНИЯ РЕГУЛЯТОРНЫХ БЕЛКОВ – ОПЕРАТОР. С ОПЕРАТОРОМ СВЯЗЫВАЕТСЯ БЕЛОК – РЕПРЕССОР. Это блокирует прочтение РНК -ПОЛИМЕРАЗОЙ СООТВЕТСТВУЮЩЕЙ ГРУППЫ ГЕНОВ.
, расположенные в позиции -30 / -20 относительно точки инициации синтеза") ТАТА-боксы - последовательности ТАТАА(А), расположенные в позиции -30 / -20 относительно точки инициации синтеза РНК. Они обнаружены в большинстве эукариотических генов; считается, что ТАТА-боксы определяют, что потенциально активная точка начала синтеза РНК оказывается реальным стартом транскрипции. СААТ-боксы — последовательности С(О)СААТ, находящиеся в положении -50 / -150. В отсутствие ТАТА-бокса могут «активировать» потенциальный старт транскрипции GС-мотивы — участки длиной 5— 15 нуклеотидных пар, состоящие из О и С; не имеют строгого консенсуса, но обычно содержат последовательность (О)ООСОО(О). В отсутствие и ТАТА-, и САА'Г боксов ОС-последовательность определяет правильную инициацию транскрипции.
ТАТА-боксы - последовательности ТАТАА(А), расположенные в позиции -30 / -20 относительно точки инициации синтеза РНК. Они обнаружены в большинстве эукариотических генов; считается, что ТАТА-боксы определяют, что потенциально активная точка начала синтеза РНК оказывается реальным стартом транскрипции. СААТ-боксы — последовательности С(О)СААТ, находящиеся в положении -50 / -150. В отсутствие ТАТА-бокса могут «активировать» потенциальный старт транскрипции GС-мотивы — участки длиной 5— 15 нуклеотидных пар, состоящие из О и С; не имеют строгого консенсуса, но обычно содержат последовательность (О)ООСОО(О). В отсутствие и ТАТА-, и САА'Г боксов ОС-последовательность определяет правильную инициацию транскрипции.
 Последовательности, непосредственно прилегающие к точкам инициации синтеза РНК, не объединены строгим консенсусом. б)") а) Последовательности, непосредственно прилегающие к точкам инициации синтеза РНК, не объединены строгим консенсусом. б) Как правило, каждая промоторная область содержит и ТАТА-бокс, и СААТ или ОС, но каждая из приведенных последовательностей может находиться в промоторе и без двух других и самостоятельно регулировать транскрипцию
а) Последовательности, непосредственно прилегающие к точкам инициации синтеза РНК, не объединены строгим консенсусом. б) Как правило, каждая промоторная область содержит и ТАТА-бокс, и СААТ или ОС, но каждая из приведенных последовательностей может находиться в промоторе и без двух других и самостоятельно регулировать транскрипцию
 Специфические регуляторные последовательности Цис-элементы являются участками связывания специфических белковых транскрипционных факторов; они расположены, как правило, на расстоянии до 300— 400 нуклеотидных пар перед точкой начала транскрипции. Их наборы индивидуальны для каждого гена, что обеспечивает специфичность регуляции транскрипции. Связывание транскрипционных факторов может приводить как к усилению, так и к подавлению транскрипции.
Специфические регуляторные последовательности Цис-элементы являются участками связывания специфических белковых транскрипционных факторов; они расположены, как правило, на расстоянии до 300— 400 нуклеотидных пар перед точкой начала транскрипции. Их наборы индивидуальны для каждого гена, что обеспечивает специфичность регуляции транскрипции. Связывание транскрипционных факторов может приводить как к усилению, так и к подавлению транскрипции.
 • могут быть расположены на расстоянии до нескольких тысяч") Энхансеры. : (Всегда активизируют транскрипцию) • могут быть расположены на расстоянии до нескольких тысяч нуклеотидных пар от кодирующей области гена; • сохраняют свою активность при изменении ориентации; • располагаются как перед, так и после кодирующей области. • Последовательности, обладающие аналогичными свойствами, но ослабляющие транскрипцию, получили название «сайленсеры» . Действие энхансеров и сайлейсеров осуществляется, как и в предыдущем случае, благодаря связыванию специфических транскрипционных факторов
Энхансеры. : (Всегда активизируют транскрипцию) • могут быть расположены на расстоянии до нескольких тысяч нуклеотидных пар от кодирующей области гена; • сохраняют свою активность при изменении ориентации; • располагаются как перед, так и после кодирующей области. • Последовательности, обладающие аналогичными свойствами, но ослабляющие транскрипцию, получили название «сайленсеры» . Действие энхансеров и сайлейсеров осуществляется, как и в предыдущем случае, благодаря связыванию специфических транскрипционных факторов
 Функциональные отделы в ДНК у эукариот
Функциональные отделы в ДНК у эукариот
 Типичный эукариотический транскрипционный фактор имеет два функциональных") Основные элементы структуры белковых транскрипционных факторов (транс-фактор) Типичный эукариотический транскрипционный фактор имеет два функциональных домена - ДНКсвязывающий и трансактивирующий (или активирующий транскрипцию). Стартовая точка транскрипции — нуклеотид, с которого начинается синтез РНК. Терминатор — последовательность ДНК, узнаваемая РНК-полимеразой и являющаяся сигналом окончания транскрипции
Основные элементы структуры белковых транскрипционных факторов (транс-фактор) Типичный эукариотический транскрипционный фактор имеет два функциональных домена - ДНКсвязывающий и трансактивирующий (или активирующий транскрипцию). Стартовая точка транскрипции — нуклеотид, с которого начинается синтез РНК. Терминатор — последовательность ДНК, узнаваемая РНК-полимеразой и являющаяся сигналом окончания транскрипции
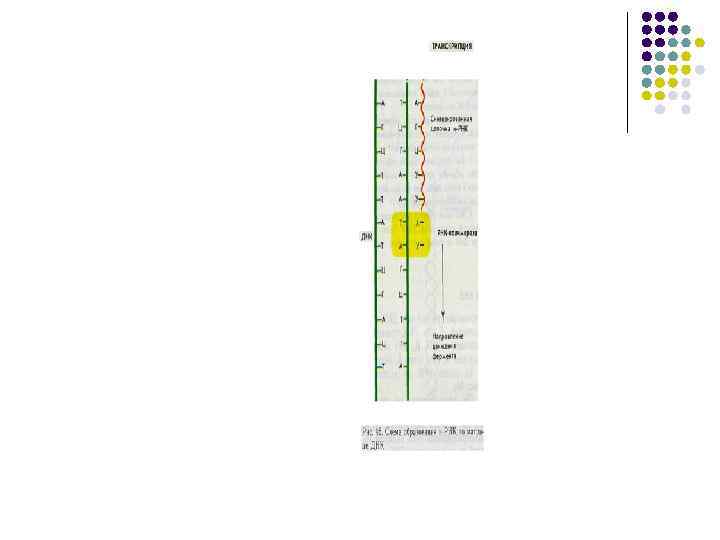
-процесс синтеза молекулы и-РНК на молекуле ДНК, выступающей в роли матрицы Транскрипция проходит 4") ТРАНСКРИПЦИЯ(переписывание)-процесс синтеза молекулы и-РНК на молекуле ДНК, выступающей в роли матрицы Транскрипция проходит 4 стадии 1. связывания ДНК-матрицы, 2. инициации, 3. элонгации 4. терминации; в рамках каждой из этих стадий осуществляются определенные функции мультимолекулярного ферментного комплекса — РНКполимеразы :
ТРАНСКРИПЦИЯ(переписывание)-процесс синтеза молекулы и-РНК на молекуле ДНК, выступающей в роли матрицы Транскрипция проходит 4 стадии 1. связывания ДНК-матрицы, 2. инициации, 3. элонгации 4. терминации; в рамках каждой из этих стадий осуществляются определенные функции мультимолекулярного ферментного комплекса — РНКполимеразы :
 Собственно РНК-полимеразная активность, т. е. образование фосфодиэфирных связей с нуклеотидами, комплементарными матрице, свойственна так называемому минимальному ферменту; таким образом, холофермент РНКполимеразы может быть разделен на минимальный фермент α 2ββ’ и σ -фактор. Инициацию синтеза РНК осуществляет только холофермент, затем σ -фактор диссоциирует и минимальный фермент ведет элонгацию. Основная функция σ -фактора — узнавание промоторов и обеспечение прочного связывания РНК-полимеразы с промоторами. Существуют минорные разновидности σ фактора, узнающие определенные группы промоторов.
Собственно РНК-полимеразная активность, т. е. образование фосфодиэфирных связей с нуклеотидами, комплементарными матрице, свойственна так называемому минимальному ферменту; таким образом, холофермент РНКполимеразы может быть разделен на минимальный фермент α 2ββ’ и σ -фактор. Инициацию синтеза РНК осуществляет только холофермент, затем σ -фактор диссоциирует и минимальный фермент ведет элонгацию. Основная функция σ -фактора — узнавание промоторов и обеспечение прочного связывания РНК-полимеразы с промоторами. Существуют минорные разновидности σ фактора, узнающие определенные группы промоторов.
 ИНИЦИАЦИЯ Узнавание промотора Сигма фактор ЭЛОНГАЦИЯ Минимальный фермент Прочное связывание РНК –полимеразы с промотором ХОЛОФЕРМЕНТ РНК – ПОЛИМЕРАЗЫ
ИНИЦИАЦИЯ Узнавание промотора Сигма фактор ЭЛОНГАЦИЯ Минимальный фермент Прочное связывание РНК –полимеразы с промотором ХОЛОФЕРМЕНТ РНК – ПОЛИМЕРАЗЫ
 ЕРНК полим ера за Генрегуля тор промо Опера тор Аттенюитор Структурны й ген Rбелок репрес сор Оперон у прокариот Тер ми на тор
ЕРНК полим ера за Генрегуля тор промо Опера тор Аттенюитор Структурны й ген Rбелок репрес сор Оперон у прокариот Тер ми на тор
 ХОЛОФЕРМЕНТ РНК – ПОЛИМЕРАЗЫ Сигма фактор Минимальный фермент Бокс Прибнова Промотор прокариот
ХОЛОФЕРМЕНТ РНК – ПОЛИМЕРАЗЫ Сигма фактор Минимальный фермент Бокс Прибнова Промотор прокариот
 Схема функционирования оперона
Схема функционирования оперона
 • В области ТАТА бокса с промотором сначала связывается ТВР – белок. • Это инициирует присоединение еще 8 ( И БОЛЕЕ) белков ТАF – белков. • Комплекс этих белков обозначается – TFIID. твр TAF TATA -бокс ПРОМОТОР
• В области ТАТА бокса с промотором сначала связывается ТВР – белок. • Это инициирует присоединение еще 8 ( И БОЛЕЕ) белков ТАF – белков. • Комплекс этих белков обозначается – TFIID. твр TAF TATA -бокс ПРОМОТОР
 Матричной для каждой молекулы РНК является одна из двух нитей ДНК; в областях перекрывания участков синтеза различных молекул РНК (транскриптонов) матричными могут быть обе цепи В КЛЕТКАХ ПРОКАРИОТ СИНТЕЗ ВСЕХ ВИДОВ РНК ОСУЩЕСТВЛЯЕТСЯ ОДНИМ ТИПОМ РНК - ПОЛИМЕРАЗЫ. ИНИЦИАЦИЯ ОБЕСПЕЧИВАЕТСЯ КОМПЛЕКСОМ ПОЛИМЕРАЗА + БЕЛКИ (6 БЕЛКОВЫХ КОМПЛЕКСОВ) –
Матричной для каждой молекулы РНК является одна из двух нитей ДНК; в областях перекрывания участков синтеза различных молекул РНК (транскриптонов) матричными могут быть обе цепи В КЛЕТКАХ ПРОКАРИОТ СИНТЕЗ ВСЕХ ВИДОВ РНК ОСУЩЕСТВЛЯЕТСЯ ОДНИМ ТИПОМ РНК - ПОЛИМЕРАЗЫ. ИНИЦИАЦИЯ ОБЕСПЕЧИВАЕТСЯ КОМПЛЕКСОМ ПОЛИМЕРАЗА + БЕЛКИ (6 БЕЛКОВЫХ КОМПЛЕКСОВ) –
 В КЛЕТКАХ ЭУКАРИОТ СУЩЕСТВУЕТ 3 РНК – ПОЛИМЕРАЗЫ: РНК – ПОЛИМЕРАЗА I – СИНТЕЗИРУЕТ РИБОСОМАЛЬНУЮ РНК – ПОЛИМЕРАЗА II – СИНТЕЗИРУЕТ ВСЕ М-РНК И И ЧАСТЬ МАЛЫХ Р- РНК – ПОЛИМЕРАЗА – III – СИНТЕЗИРУЕТ Т – РНК И ЧАСТЬ БОЛЬШИХ Р – РНК – ПОЛИМЕРАЗА ЭУКАРИОТ УСТРОЕНА БОЛЕЕ СЛОЖНО - ВКЛЮЧАЕТ В СВОЙ СОСТАВ БОЛЬШЕЕ ЧИСЛО РАЗЛИЧНЫХ ПОЛИПЕПТИДОВ (¥ 12), ЧАСТЬ ИЗ КОТОРЫХ ГОМОЛОГИЧНА ПОЛИПЕПТИДАМ РНК – ПОЛИМЕРАЗ ПРОКАРИОТ
В КЛЕТКАХ ЭУКАРИОТ СУЩЕСТВУЕТ 3 РНК – ПОЛИМЕРАЗЫ: РНК – ПОЛИМЕРАЗА I – СИНТЕЗИРУЕТ РИБОСОМАЛЬНУЮ РНК – ПОЛИМЕРАЗА II – СИНТЕЗИРУЕТ ВСЕ М-РНК И И ЧАСТЬ МАЛЫХ Р- РНК – ПОЛИМЕРАЗА – III – СИНТЕЗИРУЕТ Т – РНК И ЧАСТЬ БОЛЬШИХ Р – РНК – ПОЛИМЕРАЗА ЭУКАРИОТ УСТРОЕНА БОЛЕЕ СЛОЖНО - ВКЛЮЧАЕТ В СВОЙ СОСТАВ БОЛЬШЕЕ ЧИСЛО РАЗЛИЧНЫХ ПОЛИПЕПТИДОВ (¥ 12), ЧАСТЬ ИЗ КОТОРЫХ ГОМОЛОГИЧНА ПОЛИПЕПТИДАМ РНК – ПОЛИМЕРАЗ ПРОКАРИОТ
 ТРАНСКРИПТОН Структурный ген оператор промотор а 1 R 1 Е а 2 а 3 а 4 а 5 R 2 R 3 R 4 R 5 Э И ТЕРМИНАТОР
ТРАНСКРИПТОН Структурный ген оператор промотор а 1 R 1 Е а 2 а 3 а 4 а 5 R 2 R 3 R 4 R 5 Э И ТЕРМИНАТОР
 Механизмы репрессии и активации транскрипции l Белки, подавляющие транскрипцию, называются l l репрессорами. Если белок усиливает транскрипцию путем уменьшения активности репрессора, он называется индуктором; если транскрипция активируется непосредственно, без участия репрессора, белок называется активатором. Участки связывания репрессора или активатора на ДНК называются операторами единица транскрипции, находящаяся под контролем данного оператора, — опероном (у прокариот это, как правило, несколько генов, регулируемых параллельно транскрипции
Механизмы репрессии и активации транскрипции l Белки, подавляющие транскрипцию, называются l l репрессорами. Если белок усиливает транскрипцию путем уменьшения активности репрессора, он называется индуктором; если транскрипция активируется непосредственно, без участия репрессора, белок называется активатором. Участки связывания репрессора или активатора на ДНК называются операторами единица транскрипции, находящаяся под контролем данного оператора, — опероном (у прокариот это, как правило, несколько генов, регулируемых параллельно транскрипции
 l l Механизмы репрессии и активации транскрипции Простейший механизм репрессии — связывание репрессора с промоторной областью, т. е. перекрывание промотора и оператора. В этом случае инициации транскрипции не происходит, поскольку участок связывания РНКполимеразы уже занят репрессором. Простейший механизм активации — образование комплекса активатора и РНКполимеразы, что облегчает формирование открытого комплекса на матрице ДНК.
l l Механизмы репрессии и активации транскрипции Простейший механизм репрессии — связывание репрессора с промоторной областью, т. е. перекрывание промотора и оператора. В этом случае инициации транскрипции не происходит, поскольку участок связывания РНКполимеразы уже занят репрессором. Простейший механизм активации — образование комплекса активатора и РНКполимеразы, что облегчает формирование открытого комплекса на матрице ДНК.
 Генетический код - система записи информации в молекулах РНК, которая отражена в последовательности нуклеотидов, определяющих последовательность аминокислот в белке. Свойства генетического кода l l l Триплетность – одну аминокислоту кодирует последовательность из 3 нуклеотидов (кодон = триплет) Вырожденность – каждая аминокислота закодирована более, чем одним кодоном. Исключение – метионин ( стартовый) и триптофан. Триплеты терминации (УАА. УАГ, УГА). Универсальность. Одни и те же триплеты кодируют одинаковые аминокислоты. Однозначность – каждый триплет кодирует только одну аминокислоту. Колинеарность – совпадение последовательности аминокислот в синтезируемой молекуле белка с последовательностью триплетов в м - РНК
Генетический код - система записи информации в молекулах РНК, которая отражена в последовательности нуклеотидов, определяющих последовательность аминокислот в белке. Свойства генетического кода l l l Триплетность – одну аминокислоту кодирует последовательность из 3 нуклеотидов (кодон = триплет) Вырожденность – каждая аминокислота закодирована более, чем одним кодоном. Исключение – метионин ( стартовый) и триптофан. Триплеты терминации (УАА. УАГ, УГА). Универсальность. Одни и те же триплеты кодируют одинаковые аминокислоты. Однозначность – каждый триплет кодирует только одну аминокислоту. Колинеарность – совпадение последовательности аминокислот в синтезируемой молекуле белка с последовательностью триплетов в м - РНК
 Второе положение кодона У Ц А") Первое по ложение кодона (с 5 ' конца) Второе положение кодона У Ц А Г У УУУ Фен УУЦ УЦУ Сер УЦЦ УАУ Тир УАЦ УГУ Цис УГЦ Фен УУА Лей Сер УЦА Сер Тир УАА Стоп Цис УГА УУГ Лей УЦГ Сер УАГ Стоп УГГ Три ц ЦУУ Лей ЦУЦ ЦЦУ Про ЦЦЦ ЦАУ Гис ЦАЦ ЦГУ Арг ЦГЦ - Лей ЦУА Лей Про ЦЦА Про Гис ПДА Глн Арг ЦГА Арг ЦУГ Лей ЦЦГ Про ЦГГ Арг ЦАТ Глн Третье по ложение кодона (с З' конца) У ЦАГ У Ц АГ А АУУ Иле АУЦ АЦУ Тре АЦЦ ААУ Асн ААЦ АГУ Сер АЩ Иле АУА Иле Тре АЦА Тре Асн ААА — Сер АГА Арг АУГ Мет АЦГ Тре Лиз ААГ Лиз АГГ Арг УЦАГ Г ГУУ Вал ГУЦ ГЦУ Ала ГЦЦ ГАУ Асп ГАЦ ГГУ Гли ГГЦ Вал ГУА Вал Ала ГЦА Ала Асп ГАА Глу Гли ГГА Гли ГУГ Вал ГЦГ Ала ГГГ Гли ГАГ ГЛУ УЦАГ Таблица 2. 1. Кодоны м. РНК Сокращенные обозначения аминокислот: Ала — аланин, Арг аргинин, Асн — аспарагин, Асп — аспарагиновая кислота, Вал — ва лин, Гис — гистидин, Гли — глицин, Глн — глутамин, Глу — глутами новая кислота, Иле — изолейцин, Лей — лейцин, Лиз — лизин, Мет метионин, Про — пролин, Сер — серин. Тир — тирозин, Тре — трео нин, Три— триптофан, Фен — фенилаланин, Цис — цистеин. Стоп — «бессмысленный» кодон, прерывающий трансляцию м. РШ.
Первое по ложение кодона (с 5 ' конца) Второе положение кодона У Ц А Г У УУУ Фен УУЦ УЦУ Сер УЦЦ УАУ Тир УАЦ УГУ Цис УГЦ Фен УУА Лей Сер УЦА Сер Тир УАА Стоп Цис УГА УУГ Лей УЦГ Сер УАГ Стоп УГГ Три ц ЦУУ Лей ЦУЦ ЦЦУ Про ЦЦЦ ЦАУ Гис ЦАЦ ЦГУ Арг ЦГЦ - Лей ЦУА Лей Про ЦЦА Про Гис ПДА Глн Арг ЦГА Арг ЦУГ Лей ЦЦГ Про ЦГГ Арг ЦАТ Глн Третье по ложение кодона (с З' конца) У ЦАГ У Ц АГ А АУУ Иле АУЦ АЦУ Тре АЦЦ ААУ Асн ААЦ АГУ Сер АЩ Иле АУА Иле Тре АЦА Тре Асн ААА — Сер АГА Арг АУГ Мет АЦГ Тре Лиз ААГ Лиз АГГ Арг УЦАГ Г ГУУ Вал ГУЦ ГЦУ Ала ГЦЦ ГАУ Асп ГАЦ ГГУ Гли ГГЦ Вал ГУА Вал Ала ГЦА Ала Асп ГАА Глу Гли ГГА Гли ГУГ Вал ГЦГ Ала ГГГ Гли ГАГ ГЛУ УЦАГ Таблица 2. 1. Кодоны м. РНК Сокращенные обозначения аминокислот: Ала — аланин, Арг аргинин, Асн — аспарагин, Асп — аспарагиновая кислота, Вал — ва лин, Гис — гистидин, Гли — глицин, Глн — глутамин, Глу — глутами новая кислота, Иле — изолейцин, Лей — лейцин, Лиз — лизин, Мет метионин, Про — пролин, Сер — серин. Тир — тирозин, Тре — трео нин, Три— триптофан, Фен — фенилаланин, Цис — цистеин. Стоп — «бессмысленный» кодон, прерывающий трансляцию м. РШ.
 l Процессинг – созревание м- РНК
l Процессинг – созревание м- РНК
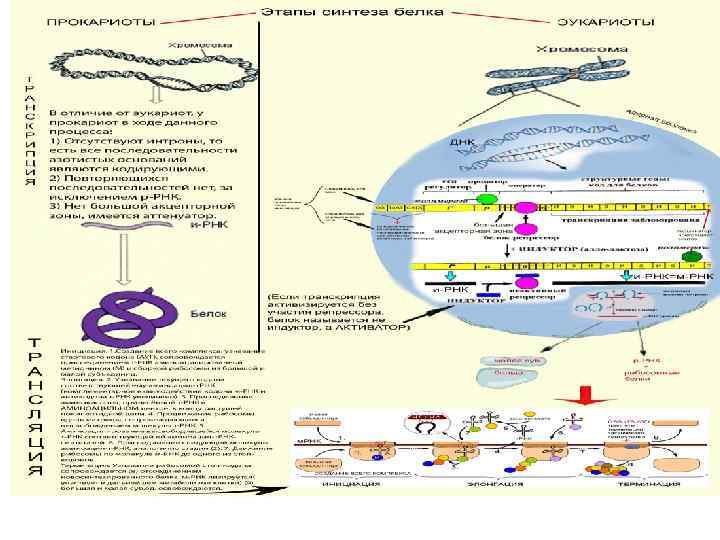
 l l l Этапы синтеза белка Транскрипция Процессинг Трансляция
l l l Этапы синтеза белка Транскрипция Процессинг Трансляция
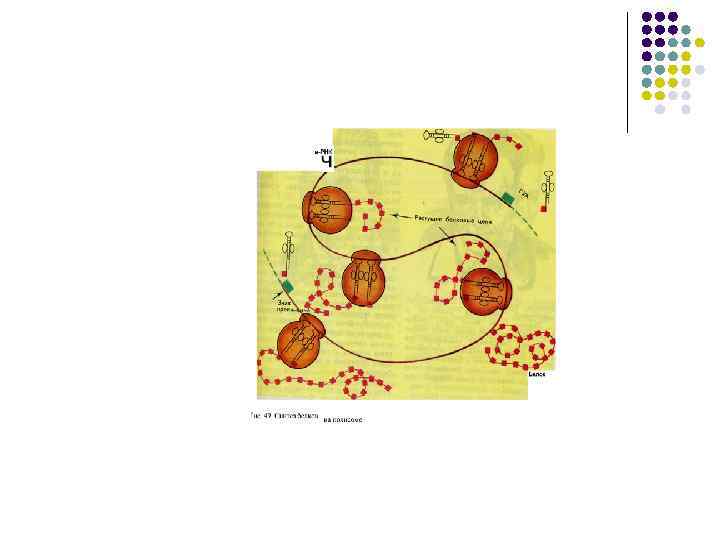
 Трансляция состоит из трех последовательных фаз: 1. Инициация – сборка всего комплекса, участвующего в синтезе молекулы белка: l Рибосомальная РНК объединяется с рибосомальными белками для формирования больших и малых субъединиц l При наличии цитоплазматического фактора инициации и зрелой м РНК субъединицы объединяются в рибосомы l т. РНК выбирает аминокислоту из «пула» аминокислот, находящихся в цитоплазме
Трансляция состоит из трех последовательных фаз: 1. Инициация – сборка всего комплекса, участвующего в синтезе молекулы белка: l Рибосомальная РНК объединяется с рибосомальными белками для формирования больших и малых субъединиц l При наличии цитоплазматического фактора инициации и зрелой м РНК субъединицы объединяются в рибосомы l т. РНК выбирает аминокислоту из «пула» аминокислот, находящихся в цитоплазме
 l 1. 2. 3. Одновременно с этим Аминокислоты в цитоплазме комбинируются с АТФ и специфическим ферментом аминоацил – т. РНК полимеразой -образутся комплекс: АМИНОКИСЛОТА+ АМФ + фермент Этот комплекс взаимодействует с т. РНК и образуется аминоацил т. РНК Декодирование м. РНК на рибосоме
l 1. 2. 3. Одновременно с этим Аминокислоты в цитоплазме комбинируются с АТФ и специфическим ферментом аминоацил – т. РНК полимеразой -образутся комплекс: АМИНОКИСЛОТА+ АМФ + фермент Этот комплекс взаимодействует с т. РНК и образуется аминоацил т. РНК Декодирование м. РНК на рибосоме
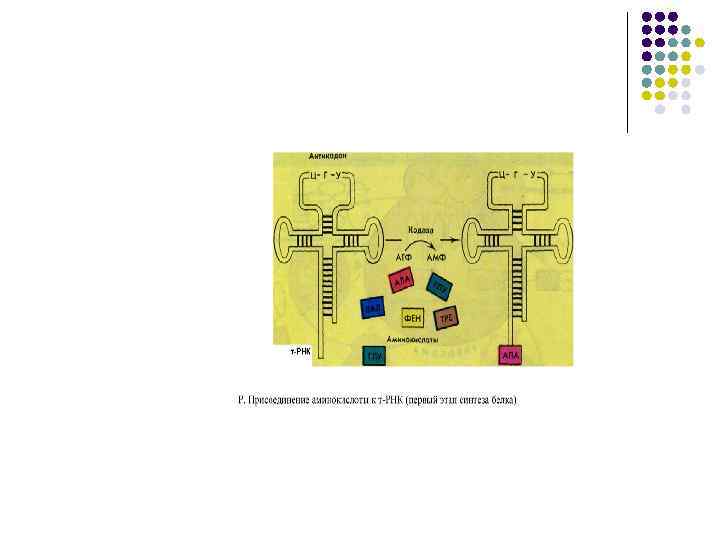
 l l l Элонгация Центр связывания матричной рнк –М находится в малой субъединице м. РНК на 5 -конце имеет кэп (шапочку=колпачок) за счет присоединения к первому нуклеотиду трифосфонуклеозида) Кэпы обеспечивают узнавание м. РНК малыми субъединицами Рядом с центром связывания находится инициирующий стартовый кодон АУГ (метионин) к которому должна подойти формилметионин -т. РНК
l l l Элонгация Центр связывания матричной рнк –М находится в малой субъединице м. РНК на 5 -конце имеет кэп (шапочку=колпачок) за счет присоединения к первому нуклеотиду трифосфонуклеозида) Кэпы обеспечивают узнавание м. РНК малыми субъединицами Рядом с центром связывания находится инициирующий стартовый кодон АУГ (метионин) к которому должна подойти формилметионин -т. РНК
 Пептидный сайт (Р)") Рибосомосвязывающий сайт м. РНК - последовательность Шайно. Дальгарно Аминоацильный сайт (А) Пептидный сайт (Р) Р А ПТЦ Пептидилтрансферазный центр. Обепечивает формирование связей между аминокислотами
Рибосомосвязывающий сайт м. РНК - последовательность Шайно. Дальгарно Аминоацильный сайт (А) Пептидный сайт (Р) Р А ПТЦ Пептидилтрансферазный центр. Обепечивает формирование связей между аминокислотами
 терминация l l Терминация – завершение синтеза белковой молекулы. В клетке нет антикодонов соответствующих кодонам терминации К рибосоме присоединяется фактор терминации (специфическая нуклеотидная последовательность). Он способствует разъединению субъединиц и освобождению синтезированного белка. Для увеличения производства белков возникают полирибосомы
терминация l l Терминация – завершение синтеза белковой молекулы. В клетке нет антикодонов соответствующих кодонам терминации К рибосоме присоединяется фактор терминации (специфическая нуклеотидная последовательность). Он способствует разъединению субъединиц и освобождению синтезированного белка. Для увеличения производства белков возникают полирибосомы