

lecture 3.ppt
- Количество слайдов: 80
 • DNA полимераза может синтезировать ДНК только в направлении 5’ to 3’ и не может сама инициировать синтез ДНК • В силу этих причит концы хромосом не могут быть полностью реплицированы обычным путем
• DNA полимераза может синтезировать ДНК только в направлении 5’ to 3’ и не может сама инициировать синтез ДНК • В силу этих причит концы хромосом не могут быть полностью реплицированы обычным путем
 Проблема репликации концов ДНК 5' 3' 3' 5' 5' 3' 5' Ori ДНК реплицируется в двух направлениях После каждого Полимеразы синтезируют ДНК в раунда репликации ДНК направлении 5' to 3' 50 -200 bp DNA на 3' конце Для начала синтеза ДНК необходим остается недореплицированными праймер
Проблема репликации концов ДНК 5' 3' 3' 5' 5' 3' 5' Ori ДНК реплицируется в двух направлениях После каждого Полимеразы синтезируют ДНК в раунда репликации ДНК направлении 5' to 3' 50 -200 bp DNA на 3' конце Для начала синтеза ДНК необходим остается недореплицированными праймер
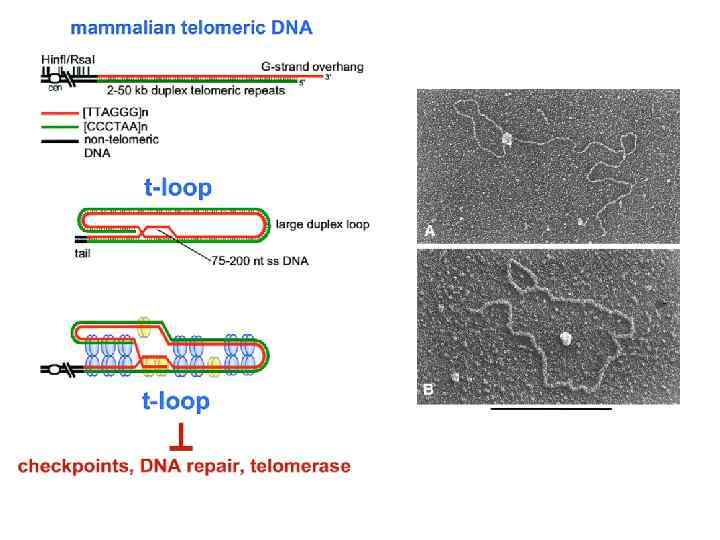
n Протяженность") Теломеры В теломерных участках хромосом расположены блоки коротких повторяющихся последовательностей у человека: (TTAGGG)n Протяженность таких блоков у человека составляет 5 -15 kilobases Особые белки связываются с теломерными повторами TRF 1 & TRF 2 Это предотвращает слияние хромосом (посредством рекомбинации)
Теломеры В теломерных участках хромосом расположены блоки коротких повторяющихся последовательностей у человека: (TTAGGG)n Протяженность таких блоков у человека составляет 5 -15 kilobases Особые белки связываются с теломерными повторами TRF 1 & TRF 2 Это предотвращает слияние хромосом (посредством рекомбинации)
 Теломераза представляет собой особую ДНК-полимеразу, которая содержит в своем составе матрицу для синтеза теломерных повторов
Теломераза представляет собой особую ДНК-полимеразу, которая содержит в своем составе матрицу для синтеза теломерных повторов
 Теломераза удлиняет выступающий 3’-конец ДНК, используя в качестве матрицы присутствующую в ее составе РНК У разных организмов теломерные повторы различаются. Соответственно, различаются и РНКматрицы в составе теломераз разных организмов
Теломераза удлиняет выступающий 3’-конец ДНК, используя в качестве матрицы присутствующую в ее составе РНК У разных организмов теломерные повторы различаются. Соответственно, различаются и РНКматрицы в составе теломераз разных организмов
 Удлиненный 3’-конец ДНК обеспечивает возможность присоединения дополнительного праймера для синтеза отстающей цепи. При этом выступающий 3’-конец все равно сохраняется RNA
Удлиненный 3’-конец ДНК обеспечивает возможность присоединения дополнительного праймера для синтеза отстающей цепи. При этом выступающий 3’-конец все равно сохраняется RNA
 2009 Nobel prize for medicine. Elizabeth Blackburn Carol Greider Jack Szostak
2009 Nobel prize for medicine. Elizabeth Blackburn Carol Greider Jack Szostak
 ИНИЦИАЦИЯ РЕПЛИКАЦИИ ДНК У ЭУКАРИОТ
ИНИЦИАЦИЯ РЕПЛИКАЦИИ ДНК У ЭУКАРИОТ
 9 У млекопитающих размер генома составляет ~2 х10 п. н. средняя скорость движения репликативных вилок у эукариот составляет ~ 2 т. п. н. /мин (для сравнения, у E. Coli ~ 60 т. п. н. /мин Таким образом, для репликации всего генома человека одной парой вилок потребовалось бы 1000000 мин = 16600 часов В клетках эукариот одновременно активируется много участков начала репликации Сегмент хромосомы, реплицирующийся с одного участка начала репликации, называется репликоном
9 У млекопитающих размер генома составляет ~2 х10 п. н. средняя скорость движения репликативных вилок у эукариот составляет ~ 2 т. п. н. /мин (для сравнения, у E. Coli ~ 60 т. п. н. /мин Таким образом, для репликации всего генома человека одной парой вилок потребовалось бы 1000000 мин = 16600 часов В клетках эукариот одновременно активируется много участков начала репликации Сегмент хромосомы, реплицирующийся с одного участка начала репликации, называется репликоном
 Bidrectional DNA synthesis Replication forks will merge
Bidrectional DNA synthesis Replication forks will merge
 Репликоны впервые были обнаружены при изучении реплицирующейся ДНК высших эукариот с использованием метода “fiber autoradiography”
Репликоны впервые были обнаружены при изучении реплицирующейся ДНК высших эукариот с использованием метода “fiber autoradiography”
 20 мин пульс 40 т. п. н. 1, 4 мкм =
20 мин пульс 40 т. п. н. 1, 4 мкм =
 Начинается ли репликация в строго определенных участках генома? Основные вопросы Существуют ли специфические последовательбности ДНК, служащие местами инициации репликации? Какие белковые факторы участвуют в инициации репликации? Как именно происходит инициация репликации? (последовательность событий) Функциональные изучение модельных систем, таких как вирусы и плазмиды, автономно реплицирующиеся в эукариотических клетках Экспериментальные подходы Создание бесклеточных систем, позволяющих воспроизвести процесс инициации репликации Структурные Картирование участков репликации в клонированных областях генома и изучение их организации
Начинается ли репликация в строго определенных участках генома? Основные вопросы Существуют ли специфические последовательбности ДНК, служащие местами инициации репликации? Какие белковые факторы участвуют в инициации репликации? Как именно происходит инициация репликации? (последовательность событий) Функциональные изучение модельных систем, таких как вирусы и плазмиды, автономно реплицирующиеся в эукариотических клетках Экспериментальные подходы Создание бесклеточных систем, позволяющих воспроизвести процесс инициации репликации Структурные Картирование участков репликации в клонированных областях генома и изучение их организации
 ARS элементы Saccharomyces cerevisiae Cloning site Selective marker cлучайные фрагменты геномной ДНК стабильный рост если плазмида реплицируется Selective marker трансформация дрожжевых клеток и высев на селективную среду нестабильный рост если плазмида не реплицируется
ARS элементы Saccharomyces cerevisiae Cloning site Selective marker cлучайные фрагменты геномной ДНК стабильный рост если плазмида реплицируется Selective marker трансформация дрожжевых клеток и высев на селективную среду нестабильный рост если плазмида не реплицируется
 Функциональные домены дрожжевых ARS элементов ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein. Subunits are named according to their size, with ORC 1 being the largest and ORC 6 being the smallest subunit. Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.
Функциональные домены дрожжевых ARS элементов ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein. Subunits are named according to their size, with ORC 1 being the largest and ORC 6 being the smallest subunit. Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.
 ARS консенсус ORC, Origin Recognition Complex состоит из 6 белков с молекулярными весами от 50 до 120 КД ORC белки чрезвычайно консервативны Они присутствуют в клетках как низших, так и высших эукариот
ARS консенсус ORC, Origin Recognition Complex состоит из 6 белков с молекулярными весами от 50 до 120 КД ORC белки чрезвычайно консервативны Они присутствуют в клетках как низших, так и высших эукариот
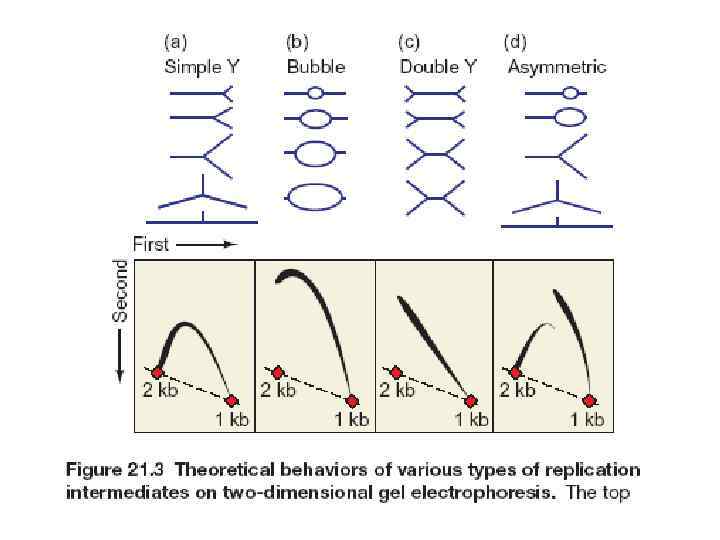
 как проверить, работает ли ARS в хромосоме? ARS 1 2 3 выбирают рестриктный фрагмент, содержащий ARS. Синтезируют несколько радиоактивных проб, узнающих различные участки этого фрагммента Геномную ДНК из делящихся клеток разделяю сначала в одном направлении в нейтральных условиях, а потом – в другом направлении в денатурирующих условиях, приводящих к освобождению коротких цепей новосинтезированной ДНК
как проверить, работает ли ARS в хромосоме? ARS 1 2 3 выбирают рестриктный фрагмент, содержащий ARS. Синтезируют несколько радиоактивных проб, узнающих различные участки этого фрагммента Геномную ДНК из делящихся клеток разделяю сначала в одном направлении в нейтральных условиях, а потом – в другом направлении в денатурирующих условиях, приводящих к освобождению коротких цепей новосинтезированной ДНК
 Нейтральный электрофорез 1 2 3 2 2 3 1 1 1 2 2 3 1 1 2 3 3 Денатурирующий форез 1
Нейтральный электрофорез 1 2 3 2 2 3 1 1 1 2 2 3 1 1 2 3 3 Денатурирующий форез 1
 2 1 Нейтральный электрофорез 1 2 3 3 2 3 1") 2 (ori) 2 1 Нейтральный электрофорез 1 2 3 3 2 3 1 1 2 2 2 3 1 1 2 3 Денатурирующий форез 1
2 (ori) 2 1 Нейтральный электрофорез 1 2 3 3 2 3 1 1 2 2 2 3 1 1 2 3 Денатурирующий форез 1
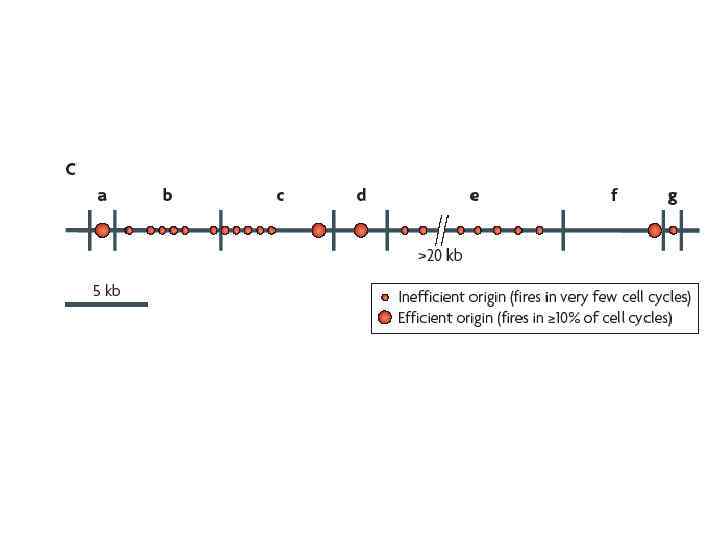
 Все реально работающие участки начала репликации в хромосомах Saccharomyces cerevisiae колокализуются c ARS элементами Существует много ARS элементов, которые не работают в качестве участков начала репликации в дрожжевых хромосомах
Все реально работающие участки начала репликации в хромосомах Saccharomyces cerevisiae колокализуются c ARS элементами Существует много ARS элементов, которые не работают в качестве участков начала репликации в дрожжевых хромосомах
 У высших эукариот поиски автономно реплицирующихся последовательностей привели к очень невыразительным и противоречивым результатам С весьма низкой эффективностью в эукариотических клетках реплицируется любая плазмидная ДНК. Включение в эту ДНК эукариотических инсерций повышает эффективнось репликации вне зависимости от последовательности ДНК. В то же время существует позитивная корреляция между длиной инсерции и эффективностью автономной репликации
У высших эукариот поиски автономно реплицирующихся последовательностей привели к очень невыразительным и противоречивым результатам С весьма низкой эффективностью в эукариотических клетках реплицируется любая плазмидная ДНК. Включение в эту ДНК эукариотических инсерций повышает эффективнось репликации вне зависимости от последовательности ДНК. В то же время существует позитивная корреляция между длиной инсерции и эффективностью автономной репликации
 В экспериментах по картированию позиций участков начала репликации в индивидуальных областях генома выявилось явное противоречие меду результатами, полученными с помощью биохимических методов (интегральная картина, показывающая наиболее вероятные события) электрофоретических методов (возможность регистрации как частых, так и редких событий)
В экспериментах по картированию позиций участков начала репликации в индивидуальных областях генома выявилось явное противоречие меду результатами, полученными с помощью биохимических методов (интегральная картина, показывающая наиболее вероятные события) электрофоретических методов (возможность регистрации как частых, так и редких событий)
 ori Новосинтезированные цепи ДНК пробы 3 2 Интенсивность ПЦР сигнала 1 4 6 4 2 0 1 2 3 4 пробы
ori Новосинтезированные цепи ДНК пробы 3 2 Интенсивность ПЦР сигнала 1 4 6 4 2 0 1 2 3 4 пробы
 Являются ли ARS участками начала репликации, работающими в геноме?
Являются ли ARS участками начала репликации, работающими в геноме?
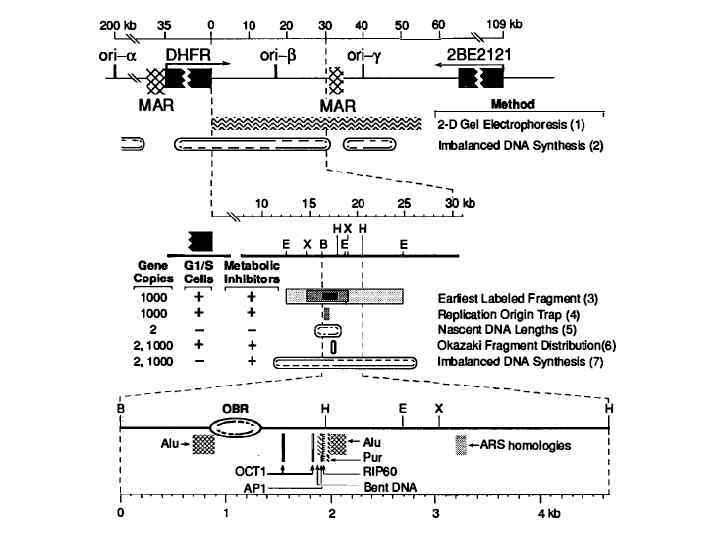
 зона, в любом участке") В 3’ концевой области гена DHFR имеется протяженная (50 Кb) зона, в любом участке которой может происходить инициация репликации
В 3’ концевой области гена DHFR имеется протяженная (50 Кb) зона, в любом участке которой может происходить инициация репликации
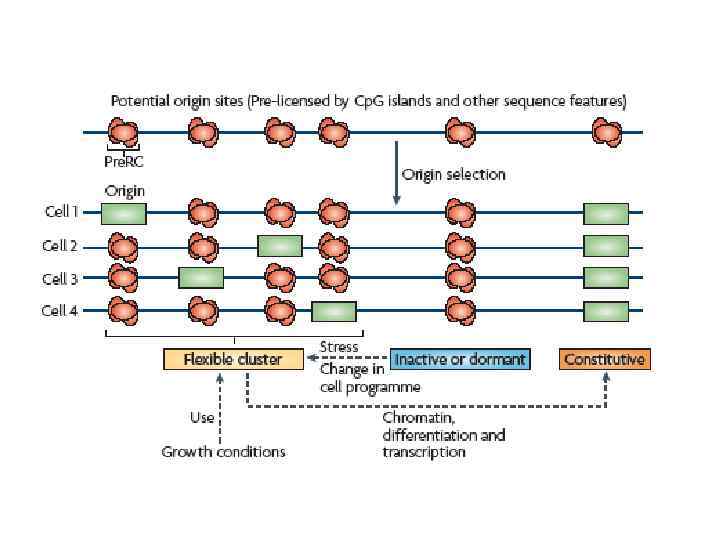
 – это миф Для того, чтобы разобраться в") типичная клетка в популяции (среднестатистическая клетка) – это миф Для того, чтобы разобраться в механизмах работы участков начала репликации, надо научиться изучать индивидуальные клетки
типичная клетка в популяции (среднестатистическая клетка) – это миф Для того, чтобы разобраться в механизмах работы участков начала репликации, надо научиться изучать индивидуальные клетки
 Molecular Biology of the Cell Vol. 17, 5337– 5345, December 2006 DNA Replication Origin Interference Increases the Spacing between Initiation Events in Human Cells Ronald Lebofsky, Roland Heilig, Max Sonnleitner, Jean Weissenbach, and Aaron Bensimon* Для идентификации репликационных вилок и участков начала репликации ДНК метили сначала иододезоксиуридином, потом хлородезоксиуридином На растянутых на стекле молекулах ДНК (molecular combes) импульсную метку выявляли с помощью каскадов антител, конъюгированных с синим и красным флуорохромами соответственно. Для того, чтобы привязать картированные участки начала репликации к изучаемой области проводили FISH с серией коротких проб, расположенных на разных расстояниях друг от друга. Для интерпретации результатов анализировали расстояния между пробами
Molecular Biology of the Cell Vol. 17, 5337– 5345, December 2006 DNA Replication Origin Interference Increases the Spacing between Initiation Events in Human Cells Ronald Lebofsky, Roland Heilig, Max Sonnleitner, Jean Weissenbach, and Aaron Bensimon* Для идентификации репликационных вилок и участков начала репликации ДНК метили сначала иододезоксиуридином, потом хлородезоксиуридином На растянутых на стекле молекулах ДНК (molecular combes) импульсную метку выявляли с помощью каскадов антител, конъюгированных с синим и красным флуорохромами соответственно. Для того, чтобы привязать картированные участки начала репликации к изучаемой области проводили FISH с серией коротких проб, расположенных на разных расстояниях друг от друга. Для интерпретации результатов анализировали расстояния между пробами
 Кластерный анализ позволил идентифицировать 41 зону инициации репликации, между которыми распределялись 307 индивидуальных участков начала репликации в пределах изучаемой области протяженностью ~ 1, 5 MB 100 KB зоны инициации репликации индивидуальные инициации репликации (белые вертикальные линии)
Кластерный анализ позволил идентифицировать 41 зону инициации репликации, между которыми распределялись 307 индивидуальных участков начала репликации в пределах изучаемой области протяженностью ~ 1, 5 MB 100 KB зоны инициации репликации индивидуальные инициации репликации (белые вертикальные линии)
 Репликационные вилки, двигающиеся от активного ori супрессируют другие ori, находящиеся в пределах зоны 55+/-38 KB в обе стороны от активного ori. Внутри области инициации наблюдается полная суппрессия всех потенциальных ori На картинке суппрессированные области инициации показаны серыми горизонтальными линиями
Репликационные вилки, двигающиеся от активного ori супрессируют другие ori, находящиеся в пределах зоны 55+/-38 KB в обе стороны от активного ori. Внутри области инициации наблюдается полная суппрессия всех потенциальных ori На картинке суппрессированные области инициации показаны серыми горизонтальными линиями
 При снижении скорости репликации происходит активация «спящих» ori
При снижении скорости репликации происходит активация «спящих» ori
 функциональный анализ участков начала репликации ДНК высших эукариот проводят посредством внедрения их мутантных копий в эктопические позиции ori 1 2 клонирование 3 4 пробы 0 направленный мутагенез вырезание и интеграция в новую геномную позицию работает 1 2 3 4 или не работает пробы 1 2 3 4 пробы
функциональный анализ участков начала репликации ДНК высших эукариот проводят посредством внедрения их мутантных копий в эктопические позиции ori 1 2 клонирование 3 4 пробы 0 направленный мутагенез вырезание и интеграция в новую геномную позицию работает 1 2 3 4 или не работает пробы 1 2 3 4 пробы
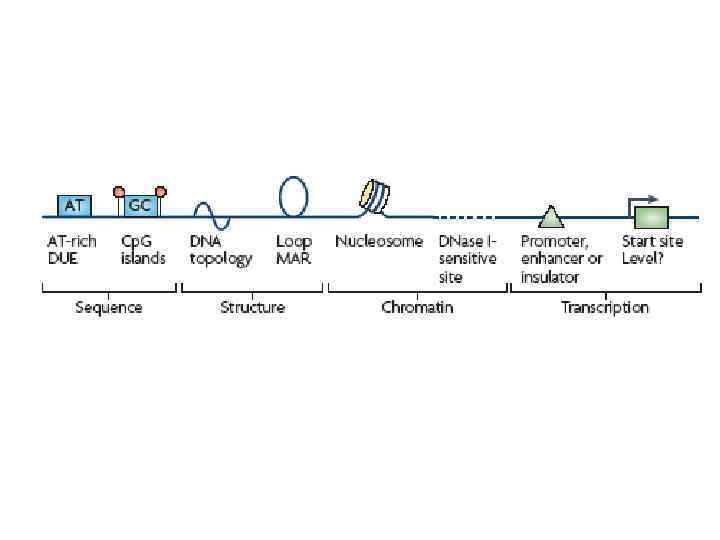
 В клетках высших эукариот инициация репликации в принципе может происходить где угодно. В эмбриональных клетках (до стадии средней бластулы) и в экстрактах из клеток ксенопуса инициация происходит через короткие интервалы (на молекуле ДНК) и без всякой специфичности в отношении последовательностей В неэмбриональных клетках сушествуют предпочтительные места инициации которые могут быть достаточно короткими (репликатор домена бета-глобиновых генов) и достаточно протяженными (зона инициации репликации локуса DHFR) Короткие репликаторы характеризуются наличием ряда необходимых элементов (АТ-богатые последовательности, полупуриновые/полипиримидиновые блоки, изогнутые последовательности ДНК). Набор и сочетание этих элементов могут быть разными в разных репликаторах. Вопрос о том, нужна ли транскрипция близлежащих генов для активации репликатора остается пока неясным
В клетках высших эукариот инициация репликации в принципе может происходить где угодно. В эмбриональных клетках (до стадии средней бластулы) и в экстрактах из клеток ксенопуса инициация происходит через короткие интервалы (на молекуле ДНК) и без всякой специфичности в отношении последовательностей В неэмбриональных клетках сушествуют предпочтительные места инициации которые могут быть достаточно короткими (репликатор домена бета-глобиновых генов) и достаточно протяженными (зона инициации репликации локуса DHFR) Короткие репликаторы характеризуются наличием ряда необходимых элементов (АТ-богатые последовательности, полупуриновые/полипиримидиновые блоки, изогнутые последовательности ДНК). Набор и сочетание этих элементов могут быть разными в разных репликаторах. Вопрос о том, нужна ли транскрипция близлежащих генов для активации репликатора остается пока неясным
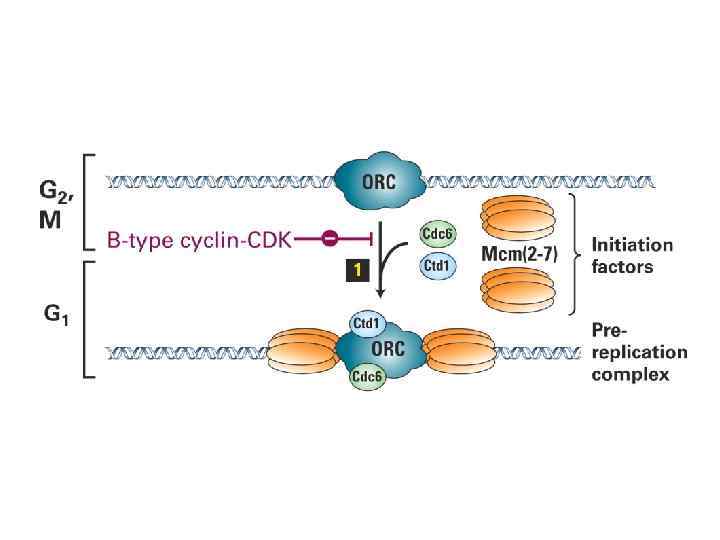
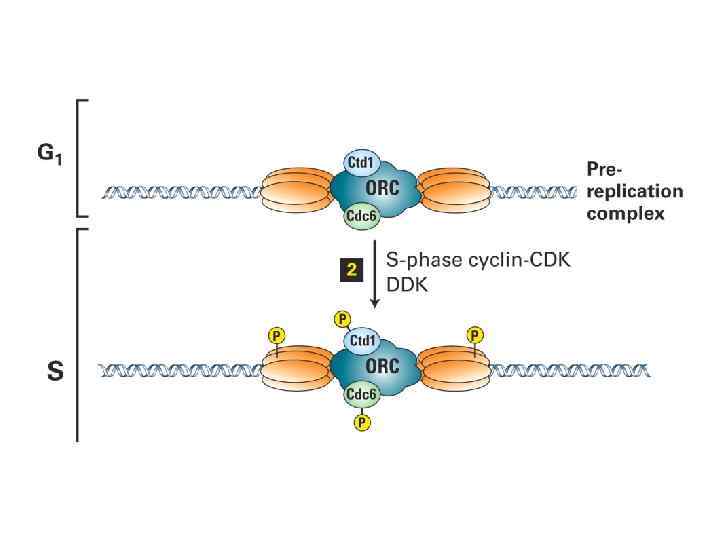
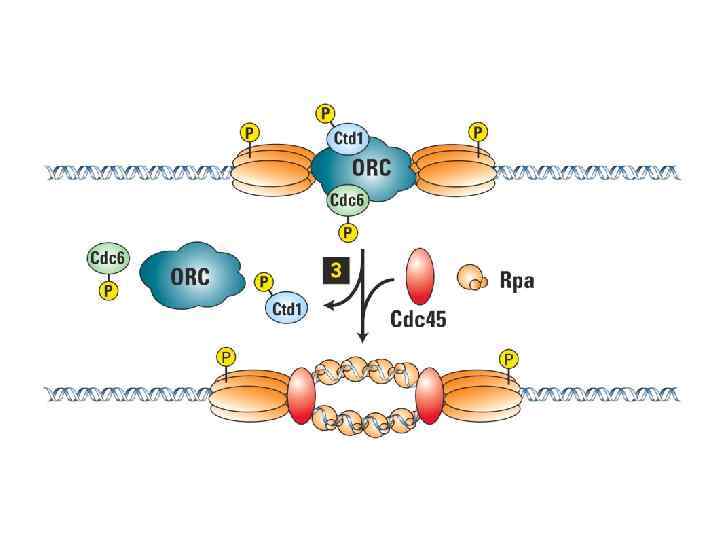
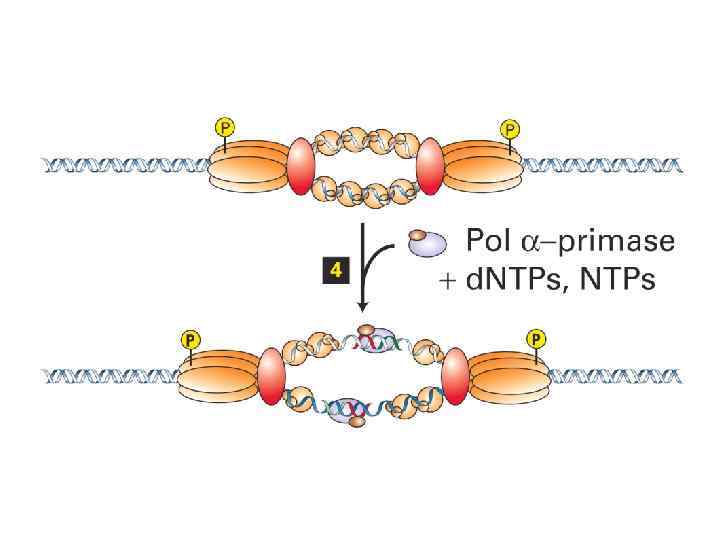
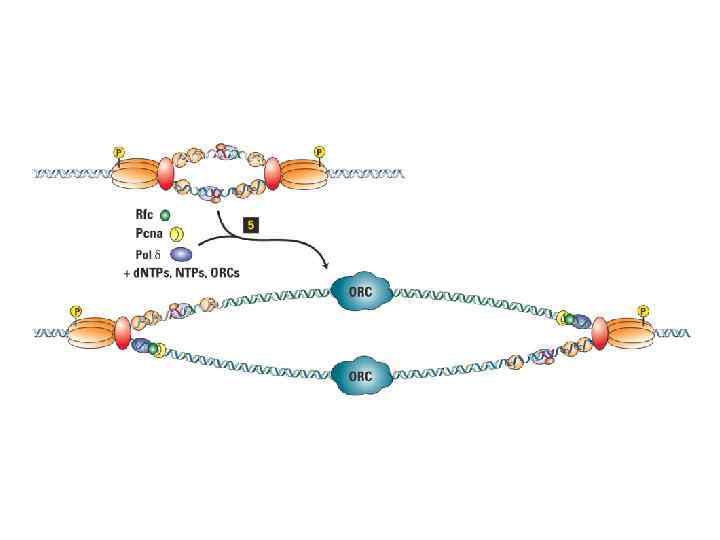
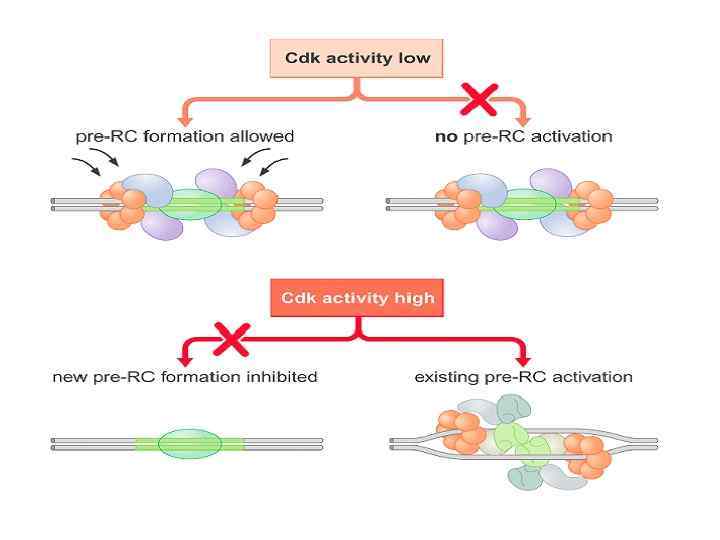
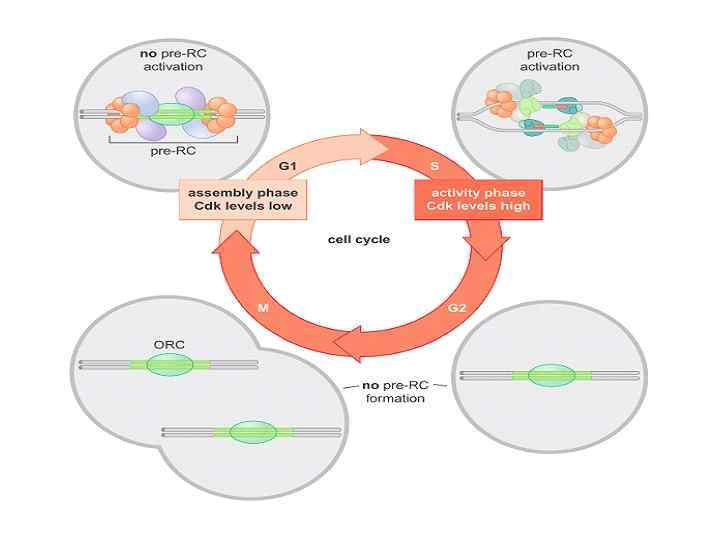
 ORC цикл, исключающий возможность повторной инициации до прохождения митоза
ORC цикл, исключающий возможность повторной инициации до прохождения митоза
 cdc-гены, сеll division control • • Реакции обратимого фосфорования Киназы, активируемые циклинами Циклины Фосфатазы • протеолиз • Убиквитинилирование (poly. Ub и mono. Ub) – протеолиз и изменение белок-белковых изаимодействий (соответственно)
cdc-гены, сеll division control • • Реакции обратимого фосфорования Киназы, активируемые циклинами Циклины Фосфатазы • протеолиз • Убиквитинилирование (poly. Ub и mono. Ub) – протеолиз и изменение белок-белковых изаимодействий (соответственно)
 Cell Cycle Checkpoints… Gap 2 Gap 1
Cell Cycle Checkpoints… Gap 2 Gap 1
 Перенос цитоплазмы из митотической клетки в интерфазную стимулирует вхождение интерфазной клетки в митоз
Перенос цитоплазмы из митотической клетки в интерфазную стимулирует вхождение интерфазной клетки в митоз
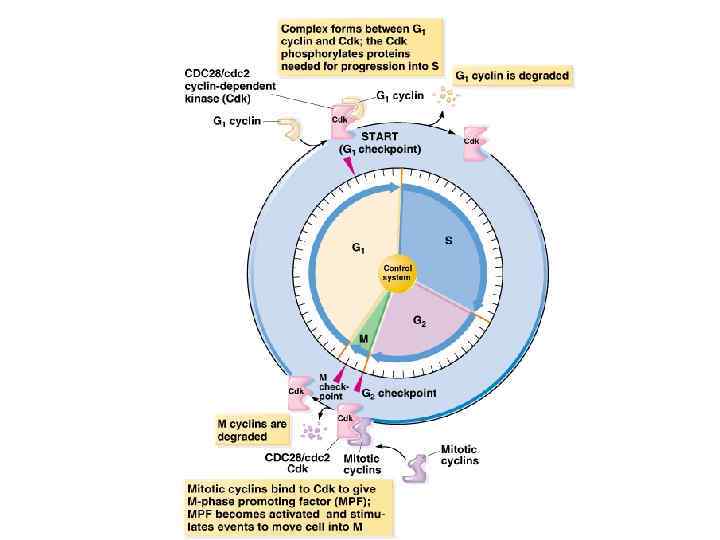
 MPF - Mitotic Promoting Factor MPF = Cdk 1 + Cyclin B Активная Cdk 1 фосфорилирует комплекс мишеней, участвующих в начале митоза (белки хромосом, ядерной оболочки, ядрышка, центросом и т. д. ) Активность Cdk 1 регулируется на двух уровнях: Присоединение циклина B = стимуляция активности Фосфорилирование треонина 14 и тирозина 15 киназой Wee 1 = инактивация (стерически препятствует связыванию АТФ с активным центром) G 2/M дефосфорилирование фосфатазой Cdc 25 M/G 1 циклин В убиквитинилируется белками APC и подвергается протеасомной деградации
MPF - Mitotic Promoting Factor MPF = Cdk 1 + Cyclin B Активная Cdk 1 фосфорилирует комплекс мишеней, участвующих в начале митоза (белки хромосом, ядерной оболочки, ядрышка, центросом и т. д. ) Активность Cdk 1 регулируется на двух уровнях: Присоединение циклина B = стимуляция активности Фосфорилирование треонина 14 и тирозина 15 киназой Wee 1 = инактивация (стерически препятствует связыванию АТФ с активным центром) G 2/M дефосфорилирование фосфатазой Cdc 25 M/G 1 циклин В убиквитинилируется белками APC и подвергается протеасомной деградации
 Cdk 1, как и другие киназы, регулирующие клеточный цикл, экспрессируется постоянно, тогда как уровень экспрессии циклинов изменяется в зависимости от фазы клеточного цикла
Cdk 1, как и другие киназы, регулирующие клеточный цикл, экспрессируется постоянно, тогда как уровень экспрессии циклинов изменяется в зависимости от фазы клеточного цикла
 контроль начала S фазы Cdk 2 - циклин E Сdk 4 Cdk 6 циклин D Cdk 4 активирует работу транскрипционного фактора E 2 F-1, включающего гены, работа которых необходима для репликации ДНК. Cdk 4 инактивирует белок Rb, который является ингибитором E 2 F-1 Cdk 2, 4. 6 являются сенсорами различных сигналов. Частью сенсорной системы являются ингибиторы Cdk (CKI).
контроль начала S фазы Cdk 2 - циклин E Сdk 4 Cdk 6 циклин D Cdk 4 активирует работу транскрипционного фактора E 2 F-1, включающего гены, работа которых необходима для репликации ДНК. Cdk 4 инактивирует белок Rb, который является ингибитором E 2 F-1 Cdk 2, 4. 6 являются сенсорами различных сигналов. Частью сенсорной системы являются ингибиторы Cdk (CKI).
 транскрипционный фактор, активирующий работу генов, участвующих в репликации ДНК
транскрипционный фактор, активирующий работу генов, участвующих в репликации ДНК
 По ходу осуществления репликации сестринские хроматиды скрепляются когезиновыми сшивками
По ходу осуществления репликации сестринские хроматиды скрепляются когезиновыми сшивками
. Секурин") В анафазе митоза клейзин расщепляется сепаразой, которая становится активной после удаления ингибитора (секурина). Секурин убиквитинилируется при посредстве APC (Anaphase Promoting Complex)
В анафазе митоза клейзин расщепляется сепаразой, которая становится активной после удаления ингибитора (секурина). Секурин убиквитинилируется при посредстве APC (Anaphase Promoting Complex)
 Индивидуальные гены реплицируются в разное время по ходу S фазы. Предложено несколько методических подходов, позволяющих определить предпочтительное время репликации гена (replication timing) Определение относительной представленности пробы в препаратах реплицирующейся ДНК, изолированных на различных стадиях S фазы Подсчет относительного количества клеток (в асинхронной культуре), содержащих две копии представляющего интерес фрагмента ДНК
Индивидуальные гены реплицируются в разное время по ходу S фазы. Предложено несколько методических подходов, позволяющих определить предпочтительное время репликации гена (replication timing) Определение относительной представленности пробы в препаратах реплицирующейся ДНК, изолированных на различных стадиях S фазы Подсчет относительного количества клеток (в асинхронной культуре), содержащих две копии представляющего интерес фрагмента ДНК
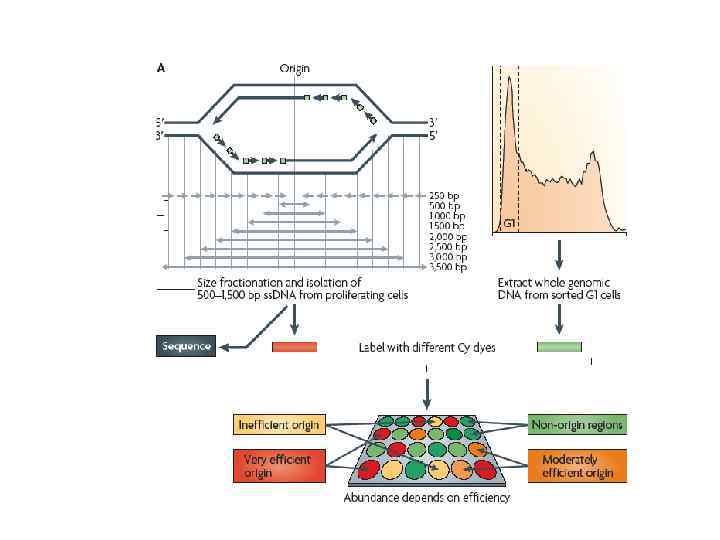
 Использование техники гибридизации с олигонуклеотидными чипами позволяет анализировать время репликации протяженных участков ДНК в рамках всего генома, либо индивидуальных хромосом Figure 1. Experimental strategy for assessing genome wide replication timing using genomic microarrays. (A) Cell cycle profile of cycling human lymphoblastoid cell line after staining with Hoechst 33259. Cells in G 1 and S phase of the cell cycle are sorted. (B) The sorted S and G 1 phase fractions are checked for purity by re-analysis on the flow sorter and DNA extracted from the fractions. (C) The extracted DNA is differentially labeled with d. CTP-Cy 3 or d. CTP-Cy 5 using random primed labeling. (D) The labeled DNA is cohybridized to the array after preannealing with Cot 1 DNA to suppress repeats. Early replicating sequences correspond to the spots with an increased S : G 1 ratio.
Использование техники гибридизации с олигонуклеотидными чипами позволяет анализировать время репликации протяженных участков ДНК в рамках всего генома, либо индивидуальных хромосом Figure 1. Experimental strategy for assessing genome wide replication timing using genomic microarrays. (A) Cell cycle profile of cycling human lymphoblastoid cell line after staining with Hoechst 33259. Cells in G 1 and S phase of the cell cycle are sorted. (B) The sorted S and G 1 phase fractions are checked for purity by re-analysis on the flow sorter and DNA extracted from the fractions. (C) The extracted DNA is differentially labeled with d. CTP-Cy 3 or d. CTP-Cy 5 using random primed labeling. (D) The labeled DNA is cohybridized to the array after preannealing with Cot 1 DNA to suppress repeats. Early replicating sequences correspond to the spots with an increased S : G 1 ratio.
 The") Replication timing profiles of two example chromosomes at a 1 Mb resolution. (A) The replication timing of chromosome 6 at a 1 Mb resolution. (B) The replication timing of chromosome 13 at a 1 Mb resolution. The black arrows indicate the gene deserts on chromosome 13.
Replication timing profiles of two example chromosomes at a 1 Mb resolution. (A) The replication timing of chromosome 6 at a 1 Mb resolution. (B) The replication timing of chromosome 13 at a 1 Mb resolution. The black arrows indicate the gene deserts on chromosome 13.
 В ядре эукариотической клетки репликоны организованы в кластеры После включения в новосинтезированную ДНК бромодезоксиуридина эти кластеры можно увидеть с использованием иммунофлуоресцентного окрашивания. Распределение активных центров репликации изменяется по ходу S-фазы
В ядре эукариотической клетки репликоны организованы в кластеры После включения в новосинтезированную ДНК бромодезоксиуридина эти кластеры можно увидеть с использованием иммунофлуоресцентного окрашивания. Распределение активных центров репликации изменяется по ходу S-фазы
 Разные типы фокусов репликации можно увидеть и анализируя распределение PCNA Мышиние клетки 3 Т 6 в геном которых интегрирован ген, кодирующий GFP-PCNA Early S Mid S Late S
Разные типы фокусов репликации можно увидеть и анализируя распределение PCNA Мышиние клетки 3 Т 6 в геном которых интегрирован ген, кодирующий GFP-PCNA Early S Mid S Late S
 В начале S-фазы в клетках присутствует несколько сотен фокусов репликации
В начале S-фазы в клетках присутствует несколько сотен фокусов репликации
 Структура эукариотического генома
Структура эукариотического генома
 Temp > 80 o C Cooling or p. H > 11 or lower p. H
Temp > 80 o C Cooling or p. H > 11 or lower p. H
 молекулы ДНК") скорость ренатурации завист от 1. концентрации ДНК 2. сложности (размеров) молекулы ДНК
скорость ренатурации завист от 1. концентрации ДНК 2. сложности (размеров) молекулы ДНК
 0") Критическими параметрами, определяющими полноту ренатурации являются концентрация ДНК и время инкубации (C T) 0 При изучении кинетики ренатурации ДНК дробят до фрагментов размером 100 -200 пн 1 2 Если взять в реакцию одинаковое количество ДНК, то фактическая концентрация «красного» фрагмента в препарате 2 будет в 10 раз меньше, чем в препарате 1. Это позволяет использовать анализ кинетики ренатурации для оценки размеров ДНК
Критическими параметрами, определяющими полноту ренатурации являются концентрация ДНК и время инкубации (C T) 0 При изучении кинетики ренатурации ДНК дробят до фрагментов размером 100 -200 пн 1 2 Если взять в реакцию одинаковое количество ДНК, то фактическая концентрация «красного» фрагмента в препарате 2 будет в 10 раз меньше, чем в препарате 1. Это позволяет использовать анализ кинетики ренатурации для оценки размеров ДНК
 for denaturation/renaturation of DNA") Cot curves (concentration x time) for denaturation/renaturation of DNA
Cot curves (concentration x time) for denaturation/renaturation of DNA
 Кривая ренатурации ДНК высших эукариот имеет несколько перегибов
Кривая ренатурации ДНК высших эукариот имеет несколько перегибов
 Наличие перегибов в кривой ренатурации ДНК высших эукариот свидетельствует о том, что она состоит из уникальных и повторяющихся последовательностей На основании степени повторенности выделяют три основных класса последовательностей: 1. Уникальные последовательности: 1 - 10 копий на гаплоидный геном (50 -60% генома млекопитающих) 2. умеренно повторяющиеся последовательности: 10 - 105 копий на гаплоидный геном (25 -40% генома млекопитающих) 3. Высокоповторяющиеся последовательности: более 105 копий на гаплоидный геном (10 -15% генома млекопитающих)
Наличие перегибов в кривой ренатурации ДНК высших эукариот свидетельствует о том, что она состоит из уникальных и повторяющихся последовательностей На основании степени повторенности выделяют три основных класса последовательностей: 1. Уникальные последовательности: 1 - 10 копий на гаплоидный геном (50 -60% генома млекопитающих) 2. умеренно повторяющиеся последовательности: 10 - 105 копий на гаплоидный геном (25 -40% генома млекопитающих) 3. Высокоповторяющиеся последовательности: более 105 копий на гаплоидный геном (10 -15% генома млекопитающих)
 2. Процессированные псевдогены 3. Повторы,") Пять классов повторяющихся последовательностей 1. Перемежающиеся повторы (Interspersed repeats) 2. Процессированные псевдогены 3. Повторы, состоящие из простых последовательностей (минисателлиты и микросателлиты) 4. Сегментные дупликации 5. Блоки тандемных повторов (сателлиты)
Пять классов повторяющихся последовательностей 1. Перемежающиеся повторы (Interspersed repeats) 2. Процессированные псевдогены 3. Повторы, состоящие из простых последовательностей (минисателлиты и микросателлиты) 4. Сегментные дупликации 5. Блоки тандемных повторов (сателлиты)
 Наибольшим числом копий представлены сателлитные ДНК Сателлитные ДНК часто отделяются от основной массы ДНК при центрифугировании в градиенте плотности Cs. Cl.
Наибольшим числом копий представлены сателлитные ДНК Сателлитные ДНК часто отделяются от основной массы ДНК при центрифугировании в градиенте плотности Cs. Cl.
 сателлитные ДНК млекопитающих состоят из вырожденных повторов, построенных по иерархическому принципу
сателлитные ДНК млекопитающих состоят из вырожденных повторов, построенных по иерархическому принципу
 Сателлитные ДНК – Размер области, содержащей повторяющиеся блоки составляет от 100 Kb до 1 Mb и более. – Альфоидный сателлит располагается в центромерных участках всех хромосом человека. Размер повторяющейся единицы 171 bp. – В ДНК каждой из хромосм человека альфоидный сателлит составляет 3 -5%
Сателлитные ДНК – Размер области, содержащей повторяющиеся блоки составляет от 100 Kb до 1 Mb и более. – Альфоидный сателлит располагается в центромерных участках всех хромосом человека. Размер повторяющейся единицы 171 bp. – В ДНК каждой из хромосм человека альфоидный сателлит составляет 3 -5%
 МИНИСАТЕЛЛИТЫ • Протяженность варьирует от 1 kb to 20 kb. • Размер повторяющейся единицы от ~10 до 500 bp. • Количество повторов может варьировать у разных особей (полиморфизм на основании которого возможено осуществление ДНК - фингерпринтинга. • Теломерные последовательности также относят к числу минисателлитов (у человека протяженность ~ 15 kb, повторяющаяся единица - GGGTTA.
МИНИСАТЕЛЛИТЫ • Протяженность варьирует от 1 kb to 20 kb. • Размер повторяющейся единицы от ~10 до 500 bp. • Количество повторов может варьировать у разных особей (полиморфизм на основании которого возможено осуществление ДНК - фингерпринтинга. • Теломерные последовательности также относят к числу минисателлитов (у человека протяженность ~ 15 kb, повторяющаяся единица - GGGTTA.
) Длина повторяющейся единицы 2") Микросателлиты также называют короткими тандемными повторами (short tandem repeats (STR)) Длина повторяющейся единицы 2 -8 bp, протяженность блока повторяющихся единиц не более 150 bp. Протяженность блоков различных микросателлитов варьирует у разных особей (основа для ДНК фингерпринтинга) Примеры микросателлитов: (A)n, (CGG)n Из динуклеотидных повторов чаще всего встречаются AC, AT, AG Экспансия микросателлитов может быть причиной возникновения различных заболеваний, связанных с нарушением функций генов
Микросателлиты также называют короткими тандемными повторами (short tandem repeats (STR)) Длина повторяющейся единицы 2 -8 bp, протяженность блока повторяющихся единиц не более 150 bp. Протяженность блоков различных микросателлитов варьирует у разных особей (основа для ДНК фингерпринтинга) Примеры микросателлитов: (A)n, (CGG)n Из динуклеотидных повторов чаще всего встречаются AC, AT, AG Экспансия микросателлитов может быть причиной возникновения различных заболеваний, связанных с нарушением функций генов
, которые дуплицированы") Сегментные дупликации Блоки от 1 до 300 Kb (чаще 10 -50 Kb), которые дуплицированы на одной хромосоме или на разных хромосомах. Могут иметь до 99% сходства по нуклеотидным последовательностям По некоторым оценкам у человека сегментные дупликации составляют ~3, 5 -5% от гаплоидного генома
Сегментные дупликации Блоки от 1 до 300 Kb (чаще 10 -50 Kb), которые дуплицированы на одной хромосоме или на разных хромосомах. Могут иметь до 99% сходства по нуклеотидным последовательностям По некоторым оценкам у человека сегментные дупликации составляют ~3, 5 -5% от гаплоидного генома
 Диспергированные повторы ~45% генома человека. Сюда входят: длина Число копий Ретроэлементы различных типов Long-terminal repeat transposons (LTRs; RNA-mediated) Long interspersed elements (LINEs); Short interspersed elements (SINEs) ДНК транспозоны (~3% от генома человека) Фракция в геноме
Диспергированные повторы ~45% генома человека. Сюда входят: длина Число копий Ретроэлементы различных типов Long-terminal repeat transposons (LTRs; RNA-mediated) Long interspersed elements (LINEs); Short interspersed elements (SINEs) ДНК транспозоны (~3% от генома человека) Фракция в геноме
 • В геноме человека есть несколько семейств LINE элементов; наиболее представлено семейство L 1. В геноме человека присутствует от 60, 000 to 100, 000 L 1 элементов. ORF 1 кодирует белок p 40, функции которого не известны. ORF 2 кодирует обратную транскриптазу, которая необходима для перемещения мобильных элементов. Красным цветом обозначены прямые повторы, которые есть у всех мобильных элементов.
• В геноме человека есть несколько семейств LINE элементов; наиболее представлено семейство L 1. В геноме человека присутствует от 60, 000 to 100, 000 L 1 элементов. ORF 1 кодирует белок p 40, функции которого не известны. ORF 2 кодирует обратную транскриптазу, которая необходима для перемещения мобильных элементов. Красным цветом обозначены прямые повторы, которые есть у всех мобильных элементов.
 SINEs Short Interspersed Nuclear Elements Длина ~ 300 bp Не содержат функциональных генов (LINE элементы с делетированной внутренней частью) У человека наиболее известо семейство Alu элементов (от 700. 000 до 1. 000 Alu элементов на гаплоидный геном)
SINEs Short Interspersed Nuclear Elements Длина ~ 300 bp Не содержат функциональных генов (LINE элементы с делетированной внутренней частью) У человека наиболее известо семейство Alu элементов (от 700. 000 до 1. 000 Alu элементов на гаплоидный геном)