Физиология возб тканей.ppt
- Количество слайдов: 67



ДИСЦИПЛИНА: «НОРМАЛЬНАЯ ФИЗИОЛОГИЯ ЧЕЛЮСТНО-ЛИЦЕВОЙ ОБЛАСТИ» доцент кафедры нормальной физиологии, кандидат медицинских наук - Частоедова Ирина Александровна

«Физиология - это научный стержень, на котором держатся все науки. В сущности в медицине имеется лишь одна наука: наука о жизни или физиология» Клод Бернар

ФИЗИОЛОГИЯ РАСТЕНИЙ НОРМАЛЬНАЯ ЖИВОТНЫХ ЧЕЛОВЕКА ПАТОЛОГИЧЕСКАЯ КЛИНИЧЕСКАЯ

ОПРЕДЕЛЕНИЕ Нормальная физиология - наука о процессах жизнедеятельности целостного организма и механизмах ее регуляции для оптимального приспособления к меняющимся условиям окружающей среды

НОРМАЛЬНАЯ ФИЗИОЛОГИЯ ОБЩАЯ ЧАСТНАЯ СПЕЦИАЛЬНАЯ КЛЕТКИ ЭВОЛЮЦИОННАЯ ВОЗРАСТНАЯ ПО РОДУ ЗАНЯТИЙ И МЕСТУ ЖИЗНИ ЧЕЛОВЕКА ОРГАНОВ И СИСТЕМ

РАЗДЕЛ: Физиология возбудимых тканей ТЕМА: Биоэлектрические явления в тканях

ПЛАН ЛЕКЦИИ • Структурно-функциональная организация клеточной мембраны • Транспорт веществ через мембрану • Ионные каналы • Возбудимые ткани, основные свойства • Биоэлектрические явления в тканях

Строение клеточной мембраны Структурная основа клеточной мембраны — матрикс — бимолекулярный слой фосфолипидов: гидрофильные части которых (головки) направлены к поверхности мембраны, а гидрофобные части (хвосты, стабилизирующие мембрану в виде бислоя) внутрь мембраны

Строение клеточной мембраны Белки мембраны: p Интегральные – встроены в мембрану: образуют ионные каналы p Периферические белки расположены либо на наружной, либо на внутренней поверхности мембраны И — интегральные белки Т — трансмембранные белки П — периферические белки Один и тот же белок может быть рецептором, ферментом и насосом
 входят в состав гликопротеинов и гликолипидов мембраны, составляя")
Строение клеточной мембраны Углеводы (преимущественно олигосахариды) входят в состав гликопротеинов и гликолипидов мембраны, составляя 2 – 10% её массы: на наружной поверхности мембран клетки формируют поверхностную оболочку — гликокаликс
 2. Восприятие изменений внешней и внутренней среды организма")
Функции клеточной мембраны 1. Барьерная (защитная) 2. Восприятие изменений внешней и внутренней среды организма с помощью рецепторов 3. Создание электрического заряда 4. Выработка биологически активных веществ 5. Передача сигналов от одной клетки к другой 6. Транспортная функция

Транспорт веществ через мембрану

Виды транспорта веществ через мембрану: Пассивный транспорт: q простая диффузия (по градиенту концентрации, электрохимическому градиенту q Облегченная диффузия (с использованием переносчиков и через ионные каналы) q Осмос (по градиенту осмотического давления) Активный транспорт: qпервично-активный – непосредственно использует энергию АТФ qвторично-активный- используется энергия градиента какого-либо иона (чаще Nа) для транспорта других ионов

Пассивный транспорт через плазматическую мембрану

Пассивный транспорт: Для облегчённой диффузии веществ необходимы встроенные в мембрану белковые компоненты: p поры p переносчики p каналы

Активный транспорт через плазматическую мембрану

Виды активного транспорта: p Унипорт — однонаправленный транспорт одного вещества

Виды активного транспорта: p Симпорт - активный перенос вещества через мембрану, осуществляемый за счет энергии градиента концентрации другого вещества

Виды активного транспорта: p Антипорт - перемещение вещества против градиента своей концентрации. При этом другое вещество движется в противоположном направлении по градиенту своей концентрации.

Виды активного транспорта:

Ионные каналы

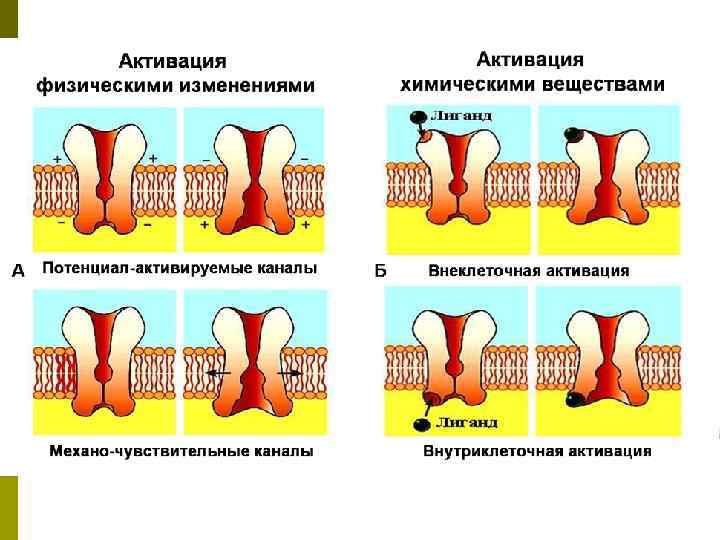
Классификация ионных каналов: По механизму управления: 1. Электровозбудимые потенциалозависимые каналы 2. Хемовозбудимые лиганд-рецепторзависимые каналы 3. Механочувствительные каналы p По скорости движения ионов: 1. Быстрые каналы 2. Медленные каналы p
 пронизывает всю толщу")
Модель калиевого канала Интегральный белок (на рисунке цифрами помечены фрагменты белка) пронизывает всю толщу липидного бислоя, формируя заполненную водой пору канала (на рисунке в канале видны 3 иона калия, нижний из ионов находится в полости поры)

Функциональные компоненты канала Воротный механизм p Сенсор p Избирательный фильтр p Пора открытого канала p

Состояния канала Состояние покоя — канал закрыт, но готов к открытию в ответ на химические, механические или электрические стимулы p Состояние активации — канал открыт и пропускает ионы p Состояние инактивации — канал закрыт и не способен к активации p

Виды ионных каналов Натриевые каналы - при открытом состоянии Na+ ‑канала ионы натрия устремляются из межклеточного пространства в цитозоль p Антагонисты натриевых каналов: местные анестетики (например, лидокаин) токсины (например, тетродотоксин) p

Виды ионных каналов p Калиевые каналы - при открытом состоянии K+ ‑канала ионы калия перемещаются из цитозоля во внеклеточное пространство - «утечка» калия из клетки p Избирательным блокирующим агентом К+ канала является тетраэтиламмоний (ТЭА), он действует только изнутри клетки

Виды ионных каналов p Кальциевые каналы —при открытом состоянии Ca 2+‑канала ионы кальция устремляются из внутриклеточных мембранных «депо кальция» и межклеточного пространства в цитозоль p Анионные каналы- через них происходит пассивный транспорт Cl– и в меньшей степени — НСО 3–. При открытом анионном канале ионы хлора диффундируют из цитозоля в межклеточное пространство

Возбудимые ткани: Мышечная Нервная Железистая

ОСНОВНЫЕ ХАРАКТЕРИСТИКИ ВОЗБУДИМЫХ ТКАНЕЙ p Раздражимость – свойство любой живой ткани отвечать на раздражитель p Возбудимость клетки - способность отвечать на раздражитель формированием потенциала действия и функциональной активностью p Возбуждение – процесс электрического, сократительного или секреторного ответа специализированной ткани, обязательным компонентом является быстрое колебание электрического потенциала клеточной мембраны
 p Проводимость –способность передавать")
ОСНОВНЫЕ СВОЙСТВА ВОЗБУДИМЫХ ТКАНЕЙ p Возбудимость (мера возбудимости- порог возбуждения) p Проводимость –способность передавать нервный импульс (мера проводимости- скорость проведения)
")
ОСНОВНЫЕ СВОЙСТВА ВОЗБУДИМЫХ ТКАНЕЙ Лабильность - способность реагировать на раздражитель с определенной скоростью (частотой) -мера лабильности - количество нервных импульсов, которое способна генерировать клетка в 1 с p Выделение секрета (медиатора) (мера – количество выделенного секрета или медиатора) p

Биоэлектрические явления в тканях

")
Первые наблюдения биоэлектрических явлений Первый опыт Гальвани В конце XVIII в. (1786 г. ) профессор анатомии Болонского университета Луиджи Гальвани провел опыты, положившие начало целенаправленным исследованиям биоэлектрических явлений

Второй опыт Гальвани Второй опыт Л. Гальвани считается опытом, в котором были получены абсолютные доказательства существования «животного электричества»
 –это разность потенциалов покоящейся клетки между")
Мембранный потенциал p Мембранный потенциал (потенциал покоя Ео) –это разность потенциалов покоящейся клетки между внутренней (отрицательной) и наружной (положительной) поверхностями клеточной мембраны.

Величина мембранного потенциала нервные клетки и кардиомиоциты от – 60 м. В до – 90 м. В p Скелетные мышцы — – 90 м. В p ГМК от - 30 до -70 м. В p Эритроциты - примерно - – 10 м. В. p

Мембранный потенциал мв +30 0 -90 Ео
 Na+ K+ ANa+ K+ K+ K+ ANa+")
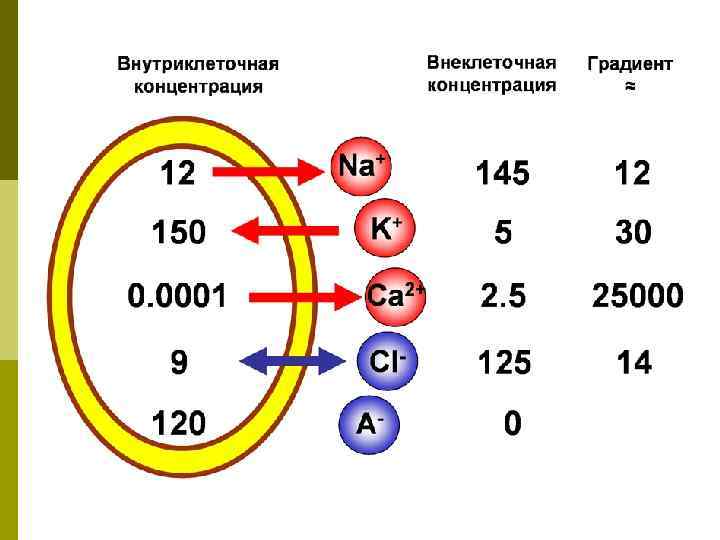
Происхождение мембранного потенциала (МП) Na+ K+ ANa+ K+ K+ K+ ANa+

Механизмы возникновения МП Главный механизм – диффузия K+ из клетки – величина МП рассчитывается по уравнению Нернста: [K+ out] Е 0 Ek 61, 5 log ----- -90 mv [K+ in] p Небольшая диффузия Na+ внутрь клетки делает реальный МП ниже, чем по формуле PK : PNa : PCl = 1 : 0, 04 : 0, 45

Механизмы возникновения МП p. Второй механизм –ассиметричная работа K+Na+-насоса K+ АТФ -аза Na+

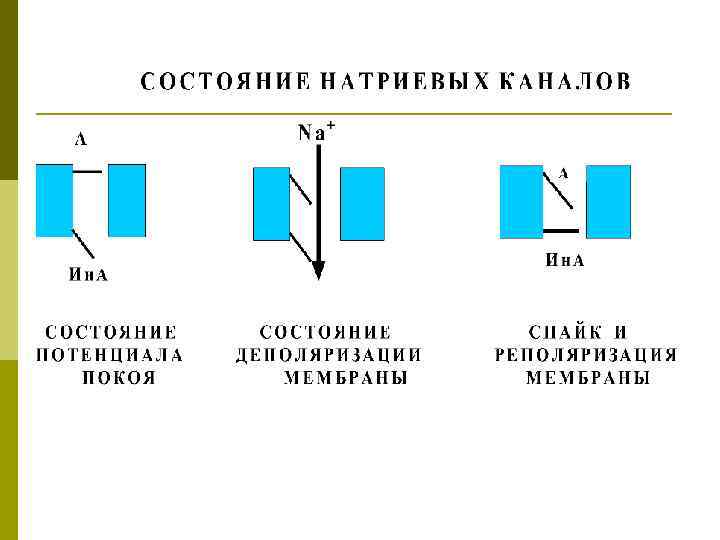
ТРИ СОСТОЯНИЯ МЕМБРАНЫ
 p Потенциал действия –кратковременное изменение МП, сопровождающееся, как правило, перезарядкой мембраны")
Потенциал действия (ПД) p Потенциал действия –кратковременное изменение МП, сопровождающееся, как правило, перезарядкой мембраны p Условия возникновения ПД: действие раздражителей пороговой и сверхпороговой силы в электровозбудимых участках мембраны

Фазы потенциала действия

Ионный механизм потенциала действия p Локальный ответ – начальная деполяризация мембраны до достижения КУД (критического уровня деполяризации) p Фаза деполяризации – открытие быстрых Na+ каналов и вход ионов Na+ внутрь клетки - перезарядка мембраны –инактивация каналов p Фаза реполяризации –открытие K+ каналов, выходящий из клетки K+ ток

Блокада калиевых каналов тетраэтиламмонием резко удлиняет процесс реполяризации
-связана с резким снижением K+ тока p")
Следовые потенциалы p Следовая деполяризация (следовой отрицательный потенциал)-связана с резким снижением K+ тока p Следовая гиперполяризация (следовой положительный потенциал) – является следствием еще сохраняющейся повышенной проницаемости клеточной мембраны для К+, она характерна для нейронов


Изменение возбудимости при возбуждении q. Фаза абсолютной рефрактерности –полная невозбудимость клетки (во время фазы деполяризации и большей части фазы реполяризации)-даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинства Na+‑каналов q Фаза относительной рефрактерности -в конце фазы реполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители
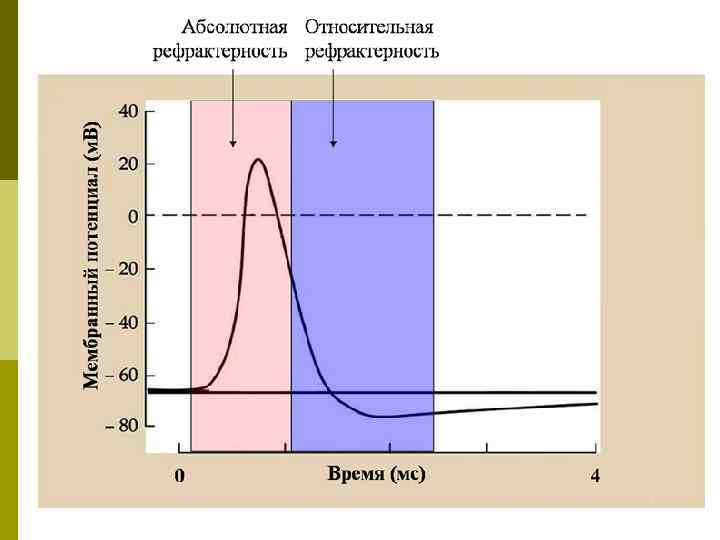

Изменение возбудимости при возбуждении q. Фаза супернормальной возбудимости –ПД можно вызвать подпороговыми раздражителями q. Фаза субнормальной возбудимости – соответствует следовой гиперполяризации - клетка способна генерировать ПД только в ответ на сверхпороговые раздражители

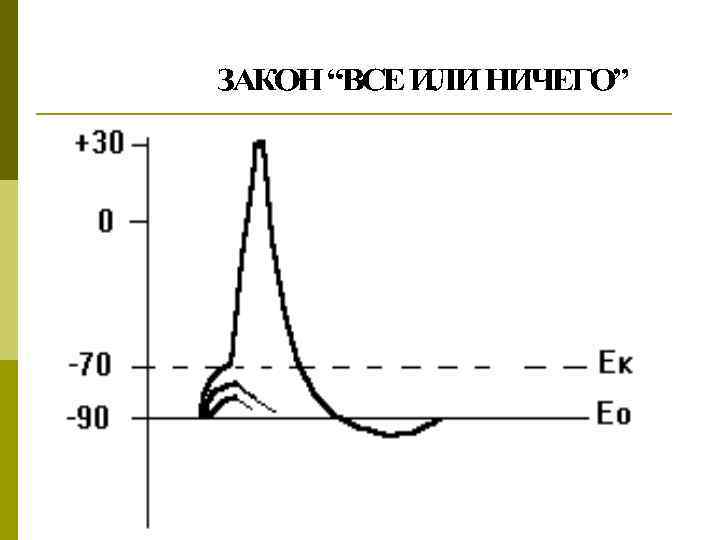
Условия, необходимые для возбуждения p Достаточная сила раздражителя (закон силы, правило «все или ничего» ) p Достаточное время действия раздражителя (закон времени) p Определенная скорость нарастания силы раздражителя (закон градиента)

Закон силы - ответная реакция в определенном диапазоне находится в прямой зависимости от силы раздражителя

Закон силы
, необходимое для снижения")
Виды раздражителей по силе действия Минимальное значение силы раздражителя (электрического тока), необходимое для снижения заряда мембраны от уровня покоя (Ео) до критического уровня (Ео), называется пороговым раздражителем. p Подпороговый раздражитель меньше по силе, чем пороговый p Сверхпороговый раздражитель - сильнее порогового p Порог раздражения или Еп = Ео - Ек p

Закон «все или ничего» Ответная реакция не зависит от силы сверхпороговых раздражителей: пороговый и сверхпороговый раздражители дают одинаковую амплитуду ответа

Взаимосвязь силы и времени действия раздражителя q. Кривая «сила -время» Гоорвега-Вейса. Лапика: q. В сверхпороговом диапазоне чем больше сила раздражителя, тем меньше время его действия, необходимое для возбуждения q. При сверхкратковременном действии раздражителя возбуждение не возникает, как бы ни была велика его сила

ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ»
 q. Для возникновения ПД необходима определенная")
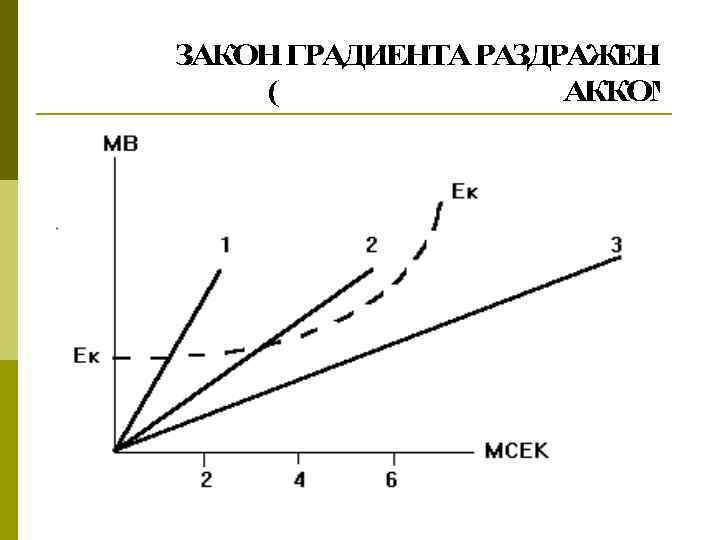
Закон градиента (крутизны нарастания Э. Дюбуа –Реймон, 1848) q. Для возникновения ПД необходима определенная скорость нарастания силы раздражителя, ниже которой возбуждение вообще не возникает q. При медленном нарастании силы возникает аккомодация (привыкание) тканей к действию раздражителя
 q. При средней силе тока возбуждение возникает под")
Закон полярности раздражения (Э. Пфлюгер, 1859) q. При средней силе тока возбуждение возникает под катодом при замыкании тока и под анодом при размыкании тока q. Механизм возникновения возбуждения под катодом при замыкании тока связан с деполяризацией мембраны, достигающей КУД q. Механизм возникновения возбуждения под анодом при размыкании тока связан с ликвидацией гиперполяризации, МП может достигать КУД

Катодическая депрессия Анодическая экзальтация
Физиология возб тканей.ppt