Гидроб-л8(дополненная).ppt
- Количество слайдов: 43



Дэмэкология - Популяции: структура, динамика Популяция гидробионта как надорганизменная система. Основы популяционного анализа. Использованы некоторые слайды из презентаций Г. С. Розенберга, Л. В. Полищука, А. М. Гилярова
 стало общепринятым.")
Разделение экологии на аут-, дем- и синэкологию (экологию особей, популяций и сообществ) стало общепринятым. И если вопросы аутэкологии чаще всего «переадресовываются» блоку физиологических наук, то дем- и синэкология – это и есть, собственно, экология. И популяционный, и экосистемный подходы в экологии имеют достаточно развитые аппараты методов исследований и свои теоретические построения.

Популяция — совокупность особей одного вида, длительно занимающая определенное пространство, воспроизводящая себя в течение большого ряда поколений. Это элементарная экологическая и биохорологическая единица вида, элементарная эволюционная единица (Тимофеев-Ресовский и др. , 1973), форма существования вида (Наумов, 1963), форма утверждения вида во внешней среде (Шварц, 1967).
 – совокупность особей одного вида с общим")
Популяция (от лат. populus – народ, население) – совокупность особей одного вида с общим генофондом, которая формируется в результате взаимодействия потока генов и условий внешней среды в пределах определен- ного пространства. Генетическое единство популяции определяет ее основное положение как элементарной единицы эволюционного процесса.
 понятие локальной популяции – совокупность особей одного вида")
В экологии часто используется упрощенное (прагматическое) понятие локальной популяции – совокупность особей одного вида на определенной экологически однородной территории. Близким к этому является понятие ценопопуляция (предложено в 1961 г. В. В. Петровским) – совокупность особей вида в пределах сообщества (чаще используется в фитоценологии). А. М. Гиляров, 1987: Под популяцией например, в планктонологии, обычно понимается совокупность особей одного вида, населяющая водоем или его часть в течение некоторого времени. (Т. е здесь под популяцией понимается некая совокупность особей одного вида, ограниченная во времени и пространстве). Строго говоря, для пространственного разграничения в одном водоеме разных популяций планктонного организма существенное значение имеет отношение скоростей водообмена и скоростей внутрипопуляционнных процессов, определяющих динамику численности изучаемых организмов. А. М. Гиляров, 1990: популяция – любая способная к самовоспроизведению совокупность особей одного вида, более или менее изолированная в пространстве и времени от других аналогичных совокупностей того же вида.
 признаки, связанные со структурой, пространственным")
Количественные признаки, характеризующие популяцию, распадаются на две группы: 1) признаки, связанные со структурой, пространственным размещением и количественными соотношениями в популяции в каждый данный момент времени – статические характеристики популяции; 2) признаки, связанные с изменением популяции во времени – динамические характеристики популяции. Статические: плотность популяции (средняя плотность) = численность в динамике (экз/м 2 или м 3) (см. разъяснения в: Смуров, Полищук, 1989); характер размещения, возрастная структура. Динамические: скорость изменения численности популяции (R, r), Рождаемость (B, b), смертность (M, m или D, d), скорость продуцирования (Р).
 определяется как количество особей, приходящихся на единицу плотности")
В статике плотность популяции (средняя плотность) определяется как количество особей, приходящихся на единицу плотности или объема. Плотность обозначается обычно как и математически записывается как , где N – численность популяции, S – площадь или объем, занятый популяцией. Через плотность определяется численность популяции – все те особи, которые в данный момент живут на данном участке и отвечают определению популяции. Численность обычно обозначают как выше (N большое, от number) и имеют в виду численность в определенный момент времени t ( time). В динамике вместо плотности говорят численность, вкладывая в это понятие тот же смысл, что и в понятие плотности. Численность ( = плотность) в динамике обычно обозначают как N с указанием того момента времени t , к которому эта величина относится - Nt. Из контекста всегда ясно, какая численность (плотность или общее количество) имеется в виду.
 свойствами-")
Популяция, как и любая сложная система, характеризуется динамикой, структурой и системными (групповыми) свойствами- характеристиками. • Плотность популяции – число особей или биомасса популяции (реже используются другие показатели; например, содержание ДНК или РНК) в расчете на единицу площади или объема.

Другие статические характеристики популяции – возрастная структура и характер размещения. Возрастная структура - соотношение численности различных возрастных групп (особей разного возраста) в популяции. Обычно изображается графически в виде кривой численности особей разного возраста или пирамиды возрастов (рис. ) При описании возрастной структуры различают 3 возрастные группы: Пререпродуктивную, Репродуктивную, Пострепродуктивную. Возраст обычно определяют по морфологическим признакам, а часто – по размерно-весовым характеристикам (размерно- возрастные группы).

σ2 = x Случайное σ2 > x Агрегированное σ2 < x Регулярное σ2 / x = 1 Случайное σ2 / x >1 Агрегиров. σ2 / x <1 Регулярное В качестве математических аналогов характера пространственного размещения особей рассматриваются теоретические статистические распределения (ТСР). ТСР наиболее удобны и информативны при относительно малых выборках и исследовании размещений на территориях, соизмеримых с занимаемыми популяциями.


При средней концентрации эуфазиид Thysanoessa raschii 50 экз. /м 3 горбатый кит должен питаться более 800 часов для того, чтобы удовлетворить суточные потребности в энергии! http: //www. arcodiv. org/Euphausiids. html
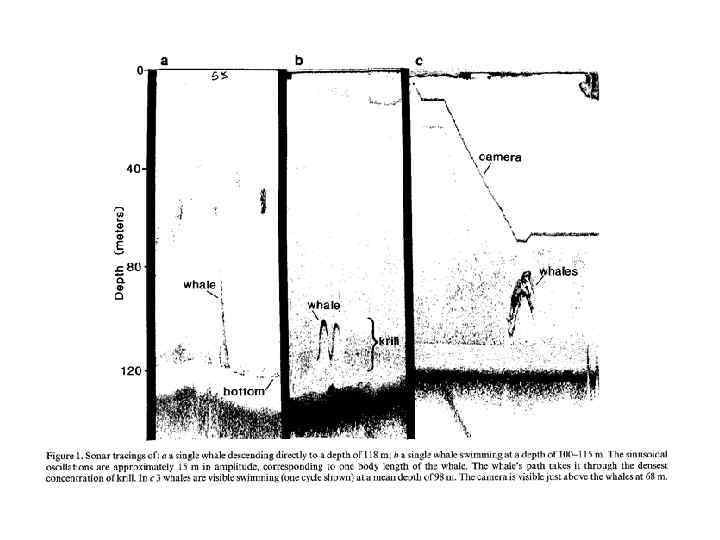

ПРИНЦИП АГРЕГАЦИИ ОСОБЕЙ Олли Скопление особей популяции, с одной стороны, усиливает конкуренцию между ними за пищевые ресурсы и жизненное пространство, с другой – приводит к повышению способности группы в целом к выживанию. Таким образом, как «перенаселенность» , так и «недонаселенность» может выступать в качестве лимитирующего фактора. Принцип сформулирован американским экологом Уардом Олли в 1931 г. Warder Clyde Allee (1885 -1955)

«Под экологической структурой популяции понимают определенное соотношение возрастных групп, определенное соотношение полов, сочетание оседлых животных с животными- мигрантами, наличие семейных, стадных и т. п. группировок. Чем сложнее структура популяции, тем выше ее приспо- собительные возможности. . . Единство приспособительных реакций популяций осуществляется с помощью сложной системы сигнализации и связи, информирующей отдельных особей о состоянии популяции в целом. Эта система информации основана на экологических и физиологических реакциях животных на внешние стимулы самой различной природы… Совокупность этих реакций спаивает особей популяции в единую функционирующую систему, обеспечивающую поддержание численности вида в разнообразной среде обитания» (Шварц, 1969, с. 14 -15). Станислав Семенович Шварц (1919 -1976)

Структура популяции Популяция состоит из особей? Да, но не с точки зрения демографии! В контексте демографии популяция состоит из когорт. Когорта – совокупность одновременно родившихся особей одного вида, обитающих в сходных условиях среды, т. е. совокупность особей-ровесников Когорта – это совокупность в точности одновозрастных особей, но на практике когорту нередко отождествляют с поколением – совокупностью особей, родившихся в течение определенного отрезка времени (например, года: сеголетки 0+, годовики 1+ и т. д. в популяции рыб). Таким образом, в каждый момент времени популяция – это совокупность когорт, родившихся в разные моменты времени в прошлом. Например, если в момент t когорта имеет возраст x лет, то она родилась x лет назад в момент t-x Когорту, входящую в состав популяции, называют также возрастной группой. То есть возрастная группа – это тоже когорта (хотя не всегда!). Используется также термин «возрастной класс» .

К динамическим характеристикам популяции относятся: - Скорость изменения численности популяции, т. е. изменение численности за определенный интервал времени, отнесенное к длине этого интервала ; - Мгновенная скорость изменения численности – производная численности по времени ; -Удельная скорость изменения численности – мгновенная скорость в расчете на одну особь ;

Экспоненциальный рост Nt = N 0 e rt r = const

Экспоненциальный рост r=b–d Удельная скорость популяционного роста как разность удельной рождаемости и удельной смертности

b– удельная рождаемость, число особей, родившихся за единицу времени в расчете на одну особь d– удельная смертность, число особей, погибших за единицу времени в расчете на одну особь Размерность b и d такая же, как и размерность r , - число особей, на единицу времени, на число особей, 1/время, или время-1

Кривые дожития кривые выживания lx – кривые

Вогнутые lx кривые: D. E. Sette в 1943 г. опубликовал данные о выживаемости ранних стадий макрели Scomber scombrus в Атлантике, в 1932 году, неблагоприятном по условиям для развития планктона Выяснилось, что в течении первых 70 суток жизни смертность макрели Scomber scombrus составила 99. 9994 % (100% - число отложенных яиц) Из миллиона икринок получалось 6 личинок, достигших длины 25 -39 мм
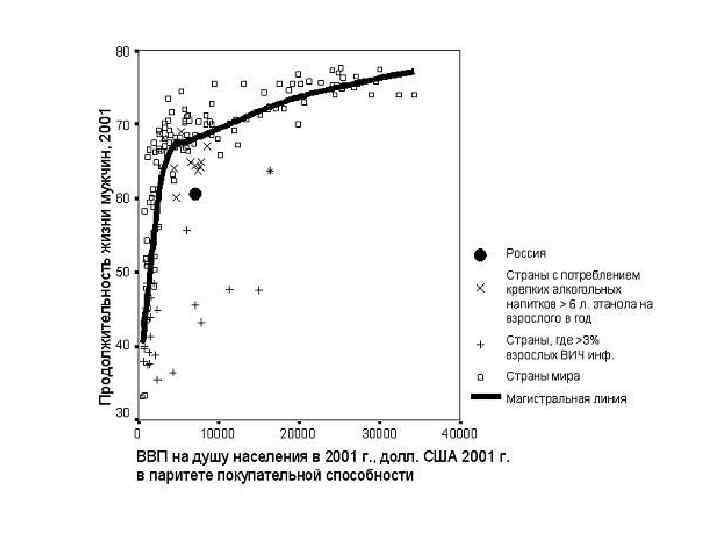
 экспоненциально пока окружающая среда по")
Первый принцип популяционной экологии: Численность популяции растет (или убывает) экспоненциально пока окружающая среда по отношению к каждой особи в этой популяции остается неизменной Среда - всё то в окружении организмов, что влияет на их выживаемость и размножение (абиотические факторы, скученность особей данного вида, а также плотности популяций всех других видов, с которыми данный вид может взаимодействовать).

ПРИНЦИП ЭКСПОНЕНЦИАЛЬНОГО РОСТА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ В БЛАГОПРИЯТНОЙ И НЕОГРАНИЧЕННОЙ СТАЦИОНАРНОЙ СРЕДЕ Это один из основных экологических принципов динамики попу- ляций. В. И. Вернадский называл этот процесс давлением жизни. Владимир Иванович Вернадский (1863 -1945)

ПРИНЦИП ЭКСПОНЕНЦИАЛЬНОГО РОСТА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ В БЛАГОПРИЯТНОЙ И НЕОГРАНИЧЕННОЙ СТАЦИОНАРНОЙ СРЕДЕ В природе экспоненциальный рост популяции практически никогда не наблюдается (если и происходит, то в течение очень непродолжительного времени, сменяясь спадом численности или выходом ее на некоторый стационарный уровень) – размер популяции всегда ограничен сверху.

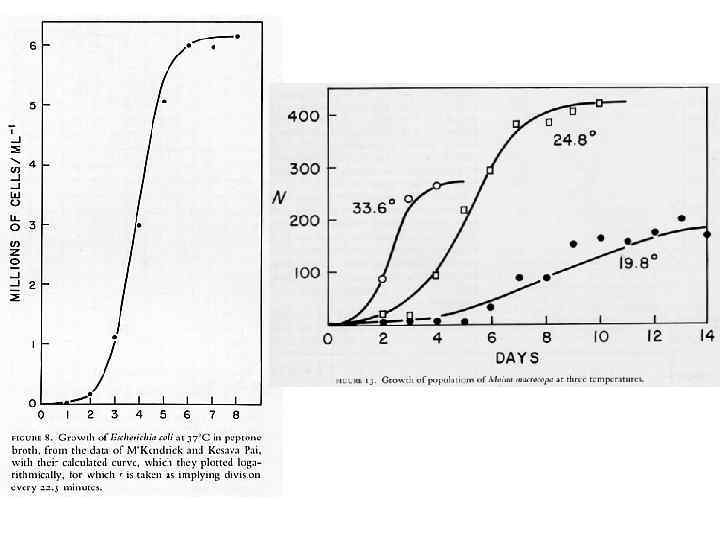

S – образный, близкий к логистическому, рост популяций может наблюдаться при самых разных механизмах ограничения плотности: 1) нехватки пищи (дафнии, водоросли, бактерии) 2) накопление продуктов метаболизма (дрожжи) 3) каннибализм (мучные жуки Tribolium) 4) поведенческие механизмы (домовые мыши в эксперименте) … и т. д. Однотипный тип математической зависимости, описывающей связь двух переменных, сам по себе не свидетельствует о том, что в основе связи лежит один и тот же биологический механизм

Разные формы зависимости удельной скорости популяционного роста r 0 от плотности популяции. Линейная зависимость при θ=1, вогнутая при θ<1
, показано, что скорость популяционного роста высока")
На 1780 сериях наблюдений (млекопитающие, птицы, рыбы, насекомые), показано, что скорость популяционного роста высока при низких значениях плотности, но она очень быстро падает при возрастании плотности. Зависимость описывается вогнутой кривой. Отсюда следует, что большинство популяций в природе существуют при плотностях, близких к ёмкости среды (величину K).

r- и K- жизненные стратегии Логистическая модель роста численности приводит к двум жизненным стратегиям (life-history strategies) – r K и Интересно, что r – буква маленькая (строчная), а K – большая (прописная), и оба этих обозначения происходят из обозначения параметров логистического уравнения

r- и K- жизненные стратегии – тайна имени K-виды, находятся в состоянии относительно стабильной численности, вблизи емкости среды K (на графике K = 1) r-виды, находятся в состоянии быстрого (близкого к экспоненциальному) роста численности с Однако эта картинка может создать ложное удельной скоростью ~ rm представление, что K-виды характеризуются высокой численностью, а r-виды – низкой. Все как раз наоборот!
 Признак r-виды K-виды Скорость роста численности")
Характеристики r- и K-видов (r- и K-стратегов) Признак r-виды K-виды Скорость роста численности Высокая, близкая к rm Низкая, много меньше rm Изменчивый во времени, Относительно постоянный во Размер популяции обычно значительно ниже времени, близкий к емкости емкости среды K Размер тела Мелкий Крупный Размер помета или кладки Большой Малый Размер потомков (отн. Мелкие Крупные размера тела взрослых) Развитие до половой Быстрое Медленное зрелости Число актов размножения за Часто один (semelparity) Часто много (iteroparity) время жизни Высокая, обычно не Низкая, обычно зависящая Смертность зависящая от плотности Продолжительность жизни Короткая Длинная Выживаемость молоди Низкая Высокая По: Pianka E. R. 1970. On r- and K-selection. Am. Nat. 104: 592 -597

Основные процессы динамики численности популяции Мерой скорости размножения является рождаемость (количество особей, родившихся в единицу времени), мерой темпа гибели является смертность (количество особей, погибших в единицу времени) Рождаемость + – Δ N – изменение Иммиграция Эмиграция численности – Смертность N – это плотность популяции, то есть численность Что означают плюсы и минусы на схеме? в расчете на единицу площади или объема (у водных организмов); размерность – число особей / км 2 или другое подобное выражение

Для чего нужно знать рождаемость и смертность? В конечном счете, как представляется, для выстраивания причинно-следственных цепочек от факторов окружающей среды к численности популяции, то есть для выявления всей совокупности внешних воздействий на популяцию с точки зрения их влияния на распространение и обилие организмов На следующем слайде показано воздействие внешних факторов на рождаемость и затем на изменение численности дафний , опосредованное внутрипопуляционными характеристиками, – плодовитостью, долей взрослых особей в популяции и скоростью развития яиц. Обратите внимание, насколько усложняется картина связей и влияний при переходе от самой грубой схемы на предыдущем слайде к схеме на следующем слайде (последняя заимствована из работы: Polishchuk L. V. Vijverberg J. Voronov D. A. Mooij W. M. Separating top-down from Bottom-up effects in Daphnia: Contribution analysis of birth rate…
 опосредовано скоростью")
Влияние среды на рождаемость b дафний b = V ln(1 + FA) опосредовано скоростью развития яиц V, плодовитостью F и долей взрослых A О К Р У скорость развития яиц температура V = 1/De Ж А Ю Щ плодовитость b А пища F = E/Na Я ΔN С размерно- доля взрослых Р избирательные A = N a/ N Е хищники Д А Факторы среды Внутрипопуляционные показатели


Возможности изложенного подхода на примере анализа динамики численности ветвистоусого рачка Bosmina coregoni из оз. Глубокого Периоды более или менее монотонного в общих чертах изменения смертности в точности совпадали по времени с периодами монотонного, но противоположного по знаку изменения рождаемости. Это наблюдение раскрывает «внутреннюю механику» основных популяционных изменений: спад численности в середине лета, пришедший на смену июньскому ее подъему, был обусловлен уменьшением рождаемости и одновременным увеличением смертности; окончание этого спада и начало августовского подъема численности связано с увеличением рождаемости и уменьшением смертности; наметившийся в конце августа спад численности вновь обусловлен уменьшением рождаемости и увеличением смертности.
, так")
Анализ по представленной на рис. 10. 4 схеме позволяет выявить как «внутренний» (внутрипопуляционный), так и «внешний» фактор, ответственный за изменение рождаемости. Ход рождаемости, не коррелируя ни с продолжительностью развития яиц, ни с долей взрослых особей, в точности повторял ход плодовитости (рис. 10. 6) и, как можно ожидать, определялся изменением трофических условий в водоеме. Это заключение подтверждается прямыми наблюдениями за сезонным ходом концентрации пищи: каждому пику рождаемости (и плодовитости) предшествовал по времени пик концентрации пригодного в пищу фитопланктона (рис. 10. 6). Соответствие кривых рождаемости и смертности указывает на существование единого внешнего фактора, управляющего изменением обоих этих показателей. На роль такого фактора есть один претендент – трофические условия. Поскольку недостаток пищи может быть непосредственной причиной гибели ветвистоусых раков, трудно сомневаться, что именно трофика, действуя через посредство рождаемости и смертности, определяла ход численности: улучшение трофических условий вызывало увеличение плодовитости и рождаемости, уменьшение смертности и, как следствие, – подъем численности; ухудшение трофических условий – наоборот, – уменьшение плодовитости и рождаемости, увеличение смертности и спад численности.
Гидроб-л8(дополненная).ppt