Клеточный цикл-2011.ppt
- Количество слайдов: 44



Деление эукариотической клетки или Клеточный цикл

НОБЕЛЕВСКИЕ ПРЕМИИ 2001 ГОДА. КЛЕТОЧНЫЙ ЦИКЛ: УНИВЕРСАЛЬНЫЙ МЕХАНИЗМ ДЛЯ ДРОЖЖЕЙ И ЧЕЛОВЕКА

Фазы клеточного цикла G 0 старт

ДНК

Эукариоты митоз Активирование белков митоза Переход из метафазы в анафазу Обычный клеточный цикл Ранний эмбриональный цикл Стимулирование синтеза ферментов репликации ДНК репликация

Сверочные точки Выход клеток из цикла происходит в естественных условиях после митоза: вместо G-периода они вступают в так называемый Go-период, или состояние покоя, хотя этот покой относителен. Go-период - это время выполнения клеткой ее специализированных функций.

Контрольно-пропускные пункты сверочные точки Способность клетки надолго задерживаться в G 0 фазе свидетельствует о наличии регуляторного механизма, который принимает решение о выходе из клеточного цикла или его продолжении. Точка клеточного цикла, в которой принимается соответствующее решение, получила название ограничительной точки (restriction point) у высших эукариот и СТАРТ (START) у почкующихся дрожжей S. cerevisiae. Пройдя эту точку клеточного цикла клетка необратимо выходит из G 1 фазы и проходит все остальные его фазы вплоть до следующей точки, где принимается новое решение. Старт Или RP У дрожжей переход через СТАРТ регулируется размером клетки и наличием во внешней среде питательных веществ. Переход через ограничительную точку высших эукариот в основном зависит от наличия во внешней среде факторов роста. Так, например, выход фибробластов кожи из фазы покоя и запуск их клеточного цикла осуществляется под действием тромбоцитарного фактора роста (PDGF), который появляется во внешней среде при свертывании крови в зоне ранения.

Как изучали? Вводили цитоплазму из клеток в М-фазе Веретено легко детектировалось на поверхности клетки Ооцит переходил в М-фазу Вводили цитоплазму из клеток в интерфазе Ооцит переходил в G 2 -фазу

Как изучали? Клетки в митозе Клетки в G 1 -фазе Клетки в митозе Клетки в S -фазе Клетки в митозе Клетки в G 2 -фазе Вывод: в М-фазе есть фактор, стимулирующий конденсацию хромосом Индуцируется конденсация хромосом из клеток G 1 -фазы Индуцируется конденсация хромосом из клеток S-фазы Метафазные хромосомы Индуцируется конденсация хромосом из клеток G 2 -фазы Метафазные хромосомы

Как изучали? Ооцит - развивающаяся яйцеклетка Существуют специальные механизмы, останавливающие такую клетку в профазе 1 мейоза Xenopus laevis

• Эксперименты, проведенные в начале 1970 -х показали, что яйца взрослых лягушек Xenopus laevis производят фактор, который будучи введенным в незрелые ооциты, находящиеся в G 2 фазе, запускает в них мейоз, таким образом, готовя их для оплодотворения. Обычно, это "взросление" вызывает прогестерон. Фактору, который достоверно не был прогестероном, дали имя maturation-promoting factor или MPF. • Так называемый "анализ лягушачьих ооцитов" позволил проверить многие клеточные экстракты на наличие MPF. Успех анализа зависел от появления в течение взросления белого пятна, которое появляется, потому что митотическое веретено перемещается из центра клетки на новую позицию, на которой обычно находятся пигменты. Экстракты из многих клеток от дрожжей до человека имели MPF активность, но она проявлялась не на всех стадиях клеточного цикла. Экстракты из клеток в G 1 и S фазе не содержали MPF. Однако когда клетка приближалась к митозу, активность появлялась, а после деления резко исчезала.

Дрожжи – удобная модельная система

Делящиеся дрожжи S. pombe оказались удобным объектом исследования. Было выделено множество мутантных штаммов с нарушенным клеточным циклом. Одна из мутаций, сdc 2 (сell division cycle), была обнаружена в начале 1980 -х Paul Nurse. Продукт гена cdc 2 играет важную роль в работе молекулярного аппарата клеточного цикла. Теперь известно, что это протеин киназа с молекулярным весом 34000 д. Очень близкая к ней протеинкиназа, являющаяся продуктом гена CDC 28, с аналогичной функцией была обнаружена у почкующихся дрожжей S. cerevisiae. Эти киназы были названы Cdk (Cyclin dependent kinase), то есть циклин-зависимыми киназами.

Основа контроля клеточного цикла: Cdk в комплексе с циклинами стимулируют различные процессы, активность Cdk исчезает при деградации циклинов. Активность есть только в комплексе с циклином!!

Циклин: -это белок!! - Постепенно синтезируется в течение всей интерфазы быстро деградирует в анафазе (протеосомы, APC, деградация когезинов) -Каждый тип циклинов, обозначенных от A до H, имеет гомологичный участок (100 а. а. , называемый " циклиновый бокс " , отвечает за связывание с CDK ). -14 белков Подсемейства циклинов : 1. G 1 -циклины ( C , D и E ) 2. митотические циклины ( A и B ).

-деградация циклина проходит быстро: Узнается убиквитин –лигазой и расщепляется протеосомой
,")
«Черная метка» Убиквитин — небольшой белок (74 аминокислотных остатка, 8. 5 к. Да), есть во всех эукариотических организмах, высококонсервативен у всех эукариот. (от англ. ubiquitous — присутствующий везде): он обнаруживается абсолютно во всех частях организма — от кожи до мозга; во всех частях живой клетки — от мембраны до ядра.

Через сульфидную группу белок Е 1 связывает убиквитин. Для активации убиквитина используется молекула АТФ. Е 1 «передаёт» активированный убиквитин ферменту Е 2. Последний узнаёт и связывает фермент Е 3 — убиквитин-лигазу, которая специфически распознаёт белки-субстраты. В результате совместного действия Е 2 и Е 3 происходит модификация субстрата молекулой убиквитина. Дальше цикл повторяется.

Внутриклеточная концентрация периодически изменяется по мере прохождения клеток через клеточный цикл
 -вариабельная часть (красный, желтый)")
-пучек спиралей -общий кор из 100 а. к. (голубой, зеленый) -вариабельная часть (красный, желтый) Все циклины имеют общую последовательность длиной 100 -150 аминокислотных остатков, называемую циклиновым боксом, которая необходима для связывания с CDK. На основании анализа кристаллической структуры циклин А 1 состоит из двух компактных центральных доменов, каждый из которых представлен пятью спиралями, и двух дополнительных спиралей у С- и N-конца.

У дрожжей один Cdk, но взаимодействует с различными циклинами

-Сами по себе циклины не могут полностью активировать соответствующие CDK. -Для завершения процесса активации должно произойти специфическое фосфорилирование и дефосфорилирование определенных остатков аминокислот в полипептидных цепях этих протеинкиназ. - Большую часть таких реакций осуществляет киназа, активирующая CDK (CAK - CDK activating kinase). CDK активны в клеточном цикле: 1. после их взаимодействия с соответствующими циклинами и 2. осуществления посттрансляционных модификаций под действием CAK и других аналогичных белков-регуляторов клеточного цикла.

Cdc 2 киназа регулируется фосфорилированием Y 15 CAK Wee 1 Cdc 25 T 165 Не активен Активен Неактивен Активен

Комплекс с циклином приводит к активации в 10 000 раз -пучек спиралей -общий кор из 100 а. к. (голубой, зеленый) -вариабельная часть (красный, желтый)

Связывание циклина А с Cdk 2 увеличивает киназную активность последней на несколько порядков. Это объясняется конформационными измененими Cdk. В основном изменения касаются Т-петли Cdk. В несвязанном состоянии Т-петля закрывает сайт связывания с белком-субстратом, что препятствует его взаимодействию с АТФ. Кроме того, небольшая спираль на конце петли вызывает конформационные изменения АТФ-связывающего сайта, что не позволяет фосфату ориентироваться правильно относительно белка субстрата. Образование циклин-киназного комплекса происходит благодаря взаимодействию спирали PSTAIRE киназы со спиралями 3 и 5 циклина, а также С-концевой части киназы и спирали N циклина. При этом спираль распрямляется и превращается в бета-складку, что позволяет спирали PSTAIRE переместиться ближе к АТФ связывающему центру. Перемещение спирали PSTAIRE исправляет конформацию АТФ - связывающего сайта, делая его активным, так как теперь фосфат ориентирован правильно. Кроме того, Т-петля выравнивается, становясь плоской, и сайт связывания с белком-субстратом становится доступным. Дополнительное фосфорилирование треонина-161 Т-петли CAK (cyclin activating kinase) приводит к дополнительной активации Cdk 2 в 80 -300 раз. Механизм этой активации следующий: фосфат треонина-161 встроен в катионный карман под Тпетлей и действует как центральный узел для сети водородных связей, стабилизирующих взаимодействие Cdk 2 и циклина А. При этом Т-петля выравнивается в одной плоскости еще сильнее и приближается к циклину, что приводит к конформационным изменениям формы связывающего белок центра. Других конформационных изменений не происходит. Следовательно, фосфорилирование Cdk необходимо лишь для улучшения связывания с белковым субстратом.

У дрожжей один Cdk, но взаимодействует с различными циклинами 1. Конденсация хромосом -Фосфорилирование конденсинов -Гистона Н 1 2. Распад ядерной оболочки -фосфорил. Ламинов 3. Фосфорил. ассоциир. белков микротрубочек -веретено деления G 1 -Cdk фосфорилирует Факторы транскрипции – Комплекс S-Cdk –не активен -фосфорилируется – активен-белки регуляторных областей начала репликации У млекопитающих в реализации цикла 9 циклинов и 7 Cdk
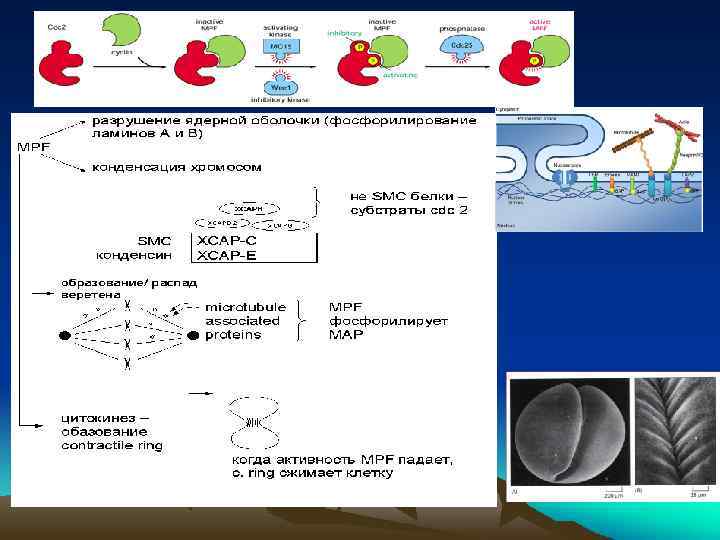

Ламина сформирована последовательностью одинаково ориентированных полимеров белков промежуточных филаментов, называемых ламинами. В клетках позвоночных она формируется в основном из ламина А , ламина В и ламина С. В-ламин имеет две формы: ламин В 1 и ламин В 2. Молекулярная масса трех главных полипептидов матрикса — примерно 60 -75 к. Да. В результате ассоциации трех главных полипептидов, путем димер-димерного взаимодействия происходит их укладка в 10 -нм структуры, присоединяющиеся к специфическим белкам ядерной мембраны через С-ламин.

Budding Yeast Fission Yeast Gene Product CDC 2 pol 3 Фенотип мутантов без функции катал. субед. ДНК-полимеразы аррест S-фазы аррест в G 2 с не качеств. реплециров. ДНК CDC 9 cdc 17 ДНК-лигаза CDC 28 cdc 2 ser/thr pr. k. SWI 6 cdc 10 регул. белок аррест на старте или при входе в митоз (G 2 checkpoint) для транскрипции G 1 цикл. аррест входа в S фазу CLN 1, 2, 3 ? G 1 циклины aаррест на старте, если все гены не активны CLB 1, 2, 3, 4 cdc 13 митотические циклины аррест на входе в митоз WEE 1 wee 1 tyr pr. K. Cdc 25 tyrosine фосфатаза ? ? RAD 9 Входят в митоз , несмотря на маленький размер аррест входа в митоз (G 2) вход в митоз при поврежденной ДНК (потеря обратной связи)

У млекопитающих в реализации цикла 9 циклинов и 7 Cdk
 белков, осуществляющих ингибирование Cdk.")
Ингибиторы Cdk. Существует два основных семейства CKI (cyclin kinase inhibitor) белков, осуществляющих ингибирование Cdk. Представители первого семейства Cip/Kip (CDK inhibitory protein) - р21, р27 и р57, ингибируют Cdk 2 и Cdk 4/6 циклиновые комплексы, осуществляя G 1 и G 1/S контроль. Представители второго семейства INK 4 (inhipitor of kinase 4) - р15, р16, р18 и р19, узкоспецифичны для CDK 4/6 -D комплексов и осуществляют аналогичные функции.
 белков, осуществляющих ингибирование Cdk.")
Ингибиторы Cdk. Существует два основных семейства CKI (cyclin kinase inhibitor) белков, осуществляющих ингибирование Cdk. Представители первого семейства Cip/Kip (CDK inhibitory protein) - р21, р27 и р57, ингибируют Cdk 2 и Cdk 4/6 циклиновые комплексы, осуществляя G 1 и G 1/S контроль. Представители второго семейства INK 4 (inhipitor of kinase 4) - р15, р16, р18 и р19, узкоспецифичны для CDK 4/6 -ц. D комплексов и осуществляют аналогичные функции. Лучше всего изучена структура молекул семейства Cip/Kip.

Обратная связь: клетка определяет, что один клеточный цикл закончился полностью до старта следующего, а так же целостность ДНК, полноту копирования ДНК, размер и сборку веретена деления

" Контактное торможение " Удалили ряд клеток Клетки на границе свободного пространства расширились, Слой клеток восстановлен за счет клеточного деления увеличили объем, то есть и свой биосинтез Важен не контакт клеток друг с другом, а степень их распластывания! -В отсутствие клеточных контактов чем меньше распластана клетка, тем больше времени занимает ее клеточный цикл. Суспензия клеток в агаре -Округление клеток сопровождается снижением общей интенсивности белкового синтеза. -Когда клетки контактируют друг с другом, это ограничивает для них возможность распластываться на субстрате, что ведет к уменьшению скорости синтеза белков, включая триггерные белки, регулирующие клеточный цикл. -Концентрация регуляторных белков в клетке падает и происходит остановка клеток в точке рестрикции. Клетки, закрепившиеся на небольшой поверхности Распластавшиеся клетки Вероятность вхождения в S-фазу

Белок ретинобластомы Функция p. Rb предотвратить чрезмерный рост клетки, запрещая продвижение по циклу, пока клетка не готова разделиться. Циклин подобные домены имеют некоторые p. Rb белки (р35) и фактор транскрипции TFIIB.

Как фосфорилирование белка ретинобластомы влияет на прохождение клеточного цикла?

Арест Продвижение
: 2363– 2368.")
Proc Natl Acad Sci U S A. 2003 March 4; 100(5): 2363– 2368.

p. Rb локализован в ядре
: 2165– 2167.")
Proc Natl Acad Sci U S A. 2003 March 4; 100(5): 2165– 2167.

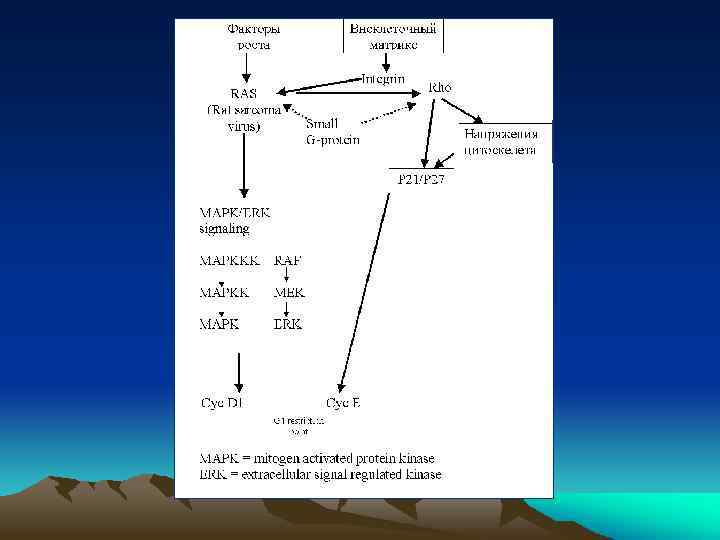
Cхема, отражающая участие некоторых белков в регуляции клеточного цикла. -это ингибиторы комплексов Cyclin/Cdk

До встречи в 9. 00 во вторник!

Genes Dev. 2008 22: 964 -969 Brigitte D. Lavoie p. Rb and condensin--local control of global chromosome structure
Клеточный цикл-2011.ppt