

Нуклеиновые кислоты.ppt
- Количество слайдов: 11

 цитозин Аденин тимин Гуанин урацил
цитозин Аденин тимин Гуанин урацил
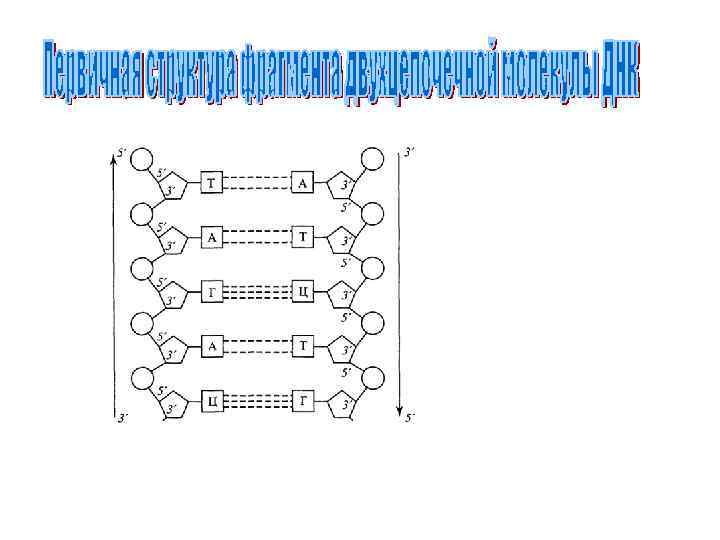
= мономерам (нуклеотидам). • Фосфат • Пентоза • Пурин/Пиримидин Пентоза") • Нуклеиновая кислота (полинуклеотид)= мономерам (нуклеотидам). • Фосфат • Пентоза • Пурин/Пиримидин Пентоза Пурин/Пиримидин
• Нуклеиновая кислота (полинуклеотид)= мономерам (нуклеотидам). • Фосфат • Пентоза • Пурин/Пиримидин Пентоза Пурин/Пиримидин
 Строение нуклеозида • N-гликозидная связь Ос но 5’ ва ни е О 4 4’ ’ С Пентоза 3’ 1’ 2’ Н
Строение нуклеозида • N-гликозидная связь Ос но 5’ ва ни е О 4 4’ ’ С Пентоза 3’ 1’ 2’ Н
 5’ • Фосфодиэфирная связь фосфат углевод основание 3’
5’ • Фосфодиэфирная связь фосфат углевод основание 3’
 Многие двухцепочечные ДНК в природе являются циклическими: плазмиды, ДНК митохондрий и хлоропластов, ДНК многих вирусов и бактерий. Такие ДНК, как правило, существуют в суперспиральном состоянии. При этом двойная спираль закручивается сама на себя, как показано на рисунке 196, количество витков образующейся суперспирали зависит от внешних условий. Суперспирализация циклических ДНК приводит к сильному изменению физических свойств молекулы, в особенности гидродинамических и электрофоретических. В клетках суперспирализация осуществляется особыми ферментами, которые для бактерий сравнительно хорошо изучены и называются ДНК-гиразами (или топоизомеразами II). Другие ферменты —топоизомеразы I — могут уменьшать число супервитков в кольцевых молекулах, давая набор «изомеров» с различным числом витков.
Многие двухцепочечные ДНК в природе являются циклическими: плазмиды, ДНК митохондрий и хлоропластов, ДНК многих вирусов и бактерий. Такие ДНК, как правило, существуют в суперспиральном состоянии. При этом двойная спираль закручивается сама на себя, как показано на рисунке 196, количество витков образующейся суперспирали зависит от внешних условий. Суперспирализация циклических ДНК приводит к сильному изменению физических свойств молекулы, в особенности гидродинамических и электрофоретических. В клетках суперспирализация осуществляется особыми ферментами, которые для бактерий сравнительно хорошо изучены и называются ДНК-гиразами (или топоизомеразами II). Другие ферменты —топоизомеразы I — могут уменьшать число супервитков в кольцевых молекулах, давая набор «изомеров» с различным числом витков.
 В качестве матрицы в реакции полимеразного копирования использовался одноцепочечный фрагмент ДНК, в качестве праймеров синтетические олигонуклеотиды или природные субфрагменты, получаемые при гидролизе рестрицирующими эндонуклеазами, а в качестве фермента - фрагмент Кленова ДНК полимеразы I (Pol. I) из E. coli. Метод включал два этапа. Сначала в ограниченных условиях проводили полимеразную реакцию в присутствии всех четырех типов d. NTP (один из них был мечен по альфа-положению фосфата), получая на выходе набор продуктов неполного копирования матричного фрагмента. Смесь очищали от несвязавшихся дезоксинуклеозидтрифосфатов и делили на восемь частей. После чего в "плюс" системе проводили четыре реакции в присутствии каждого из четырех типов нуклеотидов, а в "минус" системе - в отсутствие каждого из них. В результате, в "минус" системе терминация происходила перед d. NTP данного типа, а в "плюс" системе - после него. Полученные таким образом восемь образцов разделяли с помощью электрофореза, "считывали" сигнал и определяли последовательность исходной ДНК.
В качестве матрицы в реакции полимеразного копирования использовался одноцепочечный фрагмент ДНК, в качестве праймеров синтетические олигонуклеотиды или природные субфрагменты, получаемые при гидролизе рестрицирующими эндонуклеазами, а в качестве фермента - фрагмент Кленова ДНК полимеразы I (Pol. I) из E. coli. Метод включал два этапа. Сначала в ограниченных условиях проводили полимеразную реакцию в присутствии всех четырех типов d. NTP (один из них был мечен по альфа-положению фосфата), получая на выходе набор продуктов неполного копирования матричного фрагмента. Смесь очищали от несвязавшихся дезоксинуклеозидтрифосфатов и делили на восемь частей. После чего в "плюс" системе проводили четыре реакции в присутствии каждого из четырех типов нуклеотидов, а в "минус" системе - в отсутствие каждого из них. В результате, в "минус" системе терминация происходила перед d. NTP данного типа, а в "плюс" системе - после него. Полученные таким образом восемь образцов разделяли с помощью электрофореза, "считывали" сигнал и определяли последовательность исходной ДНК.
 Более мощный и более технологичный, этот способ, несколько модифицированный, применяется до сих пор. В основе метода тоже лежало ферментативное копирование с помощью фрагмента Кленова ДНК полимеразы I из E. coli. В качестве праймеров использовали синтетические олигонуклеотиды. Специфическую терминацию синтеза обеспечивали добавлением в реакционную смесь помимо четырех типов d. NTP (один из которых был радиоактивно мечен по альфа положению фосфата) еще и одного из 2', 3'дидезоксинуклеозидтрифосфатов (dd. ATP, dd. TTP, dd. CTP или dd. GTP), который способен включаться в растущую цепь ДНК, но не способен обеспечивать дальнейшее копирование из-за отсутствия 3'-ОН группы. Отношение концентраций d. NTP/dd. NTP авторы подбирали экспериментально, так, чтобы в итоге получить набор копий ДНК различной длины. Таким образом, для определения первичной структуры исследуемого фрагмента ДНК требовалось провести четыре реакции копирования: по одному типу терминаторов в каждой из реакций. После этого полученные продукты разгонялись в полиакриламидном геле на соседних дорожках и по расположению полос определялась последовательность нуклеотидов.
Более мощный и более технологичный, этот способ, несколько модифицированный, применяется до сих пор. В основе метода тоже лежало ферментативное копирование с помощью фрагмента Кленова ДНК полимеразы I из E. coli. В качестве праймеров использовали синтетические олигонуклеотиды. Специфическую терминацию синтеза обеспечивали добавлением в реакционную смесь помимо четырех типов d. NTP (один из которых был радиоактивно мечен по альфа положению фосфата) еще и одного из 2', 3'дидезоксинуклеозидтрифосфатов (dd. ATP, dd. TTP, dd. CTP или dd. GTP), который способен включаться в растущую цепь ДНК, но не способен обеспечивать дальнейшее копирование из-за отсутствия 3'-ОН группы. Отношение концентраций d. NTP/dd. NTP авторы подбирали экспериментально, так, чтобы в итоге получить набор копий ДНК различной длины. Таким образом, для определения первичной структуры исследуемого фрагмента ДНК требовалось провести четыре реакции копирования: по одному типу терминаторов в каждой из реакций. После этого полученные продукты разгонялись в полиакриламидном геле на соседних дорожках и по расположению полос определялась последовательность нуклеотидов.
 Рестрикция ДНК • это разрезание молекулы ДНК специфическими ферментами, которые называются эндонуклеазами рестрикции.
Рестрикция ДНК • это разрезание молекулы ДНК специфическими ферментами, которые называются эндонуклеазами рестрикции.
 Схема рестрикции
Схема рестрикции