Лекция 1-2017 клеточная стенка .ppt
- Количество слайдов: 95



Цитология грибной клетки

Клеточные покровы и клеточная стенка грибов

• • • • • Введение. Общая характеристика клеточной стенки 1. 1. Функции и методы изучения. 1. 2. Строение клеточных покровов грибов. 1. 3. Структура и состав клеточной стенки грибов 1. 3. 1. Сахара, участвующие в построении клеточной стенки грибов 1. 3. 1. 1. Хитин 1. 3. 1. 2. Хитозан 1. 3. Глюканы 1. 3. 2. Белки 1. 3. 3. Внеклеточный чехол (капсула) или внеклеточный слизистый материал 1. 3. 4. Характеристика минорных компонентов клеточной стенки грибов 1. 4. Биосинтез клеточной стенки грибов 1. 4. 1. Синтез хитина 1. 4. 2. Синтез глюканов 1. 4. 3. Синтез гликопротеинов 1. 5. Преобразования клеточных покровов в процессе морфогенеза 1. 6. Клеточные стенки в установлении и развитии патогенных и симбиотических связей между грибами и другими организмами. 1. 7. Действие стресса на клеточную стенку

Введение. Общая характеристика клеточной стенки • Клеточная стенка грибов представляет собой сложную внеклеточную структуру, окружающую клетки. Стенка грибной клетки имеет уникальные черты организации, и состав клеточной стенки грибов является важным филогенетическим и таксономическим признаком представителей царства грибов. Она определяет жизненную форму грибов (мицелиальную или дрожжеподобную) и осуществляет связь грибной клетки с внешним миром.

Введение. Общая характеристика клеточной стенки • Стенка грибов — тонко организованный сложный комплекс разнообразных полисахаридов, белков, липидов, пигментов и минеральных солей. Молекулярный состав и организация полимеров клеточной стенки могут быть различными у представителей разных таксонов, на разных стадиях жизненного цикла, при различных условиях культивирования. • Клеточная стенка — динамичный компартмент, который изменяется в течение жизни грибов.
 • Клеточная стенка как пограничная органелла осуществляет")
Функции клеточной стенки и клеточных покровов (КС) • Клеточная стенка как пограничная органелла осуществляет взаимодействие с окружением и защиту от внешней среды обитания. Клеточная стенка обеспечивает взаимодействие с внешней средой, что особенно важно для грибов, питающихся осмотрофно. Кроме того, при участии компонентов клеточной стенки осуществляется узнавание грибных клеток между собой и узнавание хозяина в случае патогенных видов. Клеточная стенка первая подвергается опасности действия окружающих факторов или ингибиторов, которые образует хозяин, благодаря наличию рецепторов она осуществляет передачу сигналов в клетку и реагирует синтезом необходимых для нормального функционирования метаболитов. Обладает избирательной проницаемостью для питательных субстратов. В зависимости от условий окружающей среды полисахаридный состав, структура и толщина стенки сильно варьирует и скоординированы с клеточным циклом и циклом развития гриба

Узнавание экзогенных и эндогенных сигнальных молекул во время инфекции оомицетом приводит к активации иммунитета растения

Взаимодействие экзосом патогенного гриба Cryptococcus neoformans с клетками животных

Функции КС • Морфогенетическая роль в определении образцов роста клеток грибов. Благодаря разному составу и способу отложения материала клеточных стенок, образуются мицелиальные или дрожжевые клетки. Клеточная стенка должна оставаться пластичной для обеспечения роста, деления клеток и формирования разных типов клеток (вегетативный мицелий, споры, плодовые тела, склероции и т. д. ) в цикле развития. Ее разрушение приводит к нарушению роста и морфологии клеток грибов и увеличивает вероятность быстрой гибели клеток. Клеточная стенка является мишенью для антигрибных препаратов.
 грибов верхушки гиф быстро")
Функции КС • У обработанных полиоксином или никомицином (ингибиторы хитинсинтаз) грибов верхушки гиф быстро набухают и ”взрываются”; делящиеся дрожжевые клетки также формируют ”взрывающиеся пары” с нарушенным разделением дочерней клетки от материнской. Были попытки использовать полиоксины в качестве с/х фунгицидов, однако к ним быстро развивается устойчивость, т. к. обработанные грибы изменяют свою транспортную систему, которая прекращает поступление антибиотиков в клетку.

Функции КС • Стабилизирует внутреннее осмотическое давление, что очень важно для сохранения формы клетки. Внутри клеток грибов создается давление равное 2, 5 атмосфер. Высокое внутриклеточное давление необходимо для всасывания питательных веществ и продвижения в плотном субстрате.

Функции КС • Защищает против физического и химического стресса, функционирует как защитный покров. Комбинирование механической прочности и высокой эластичности за счет присутствия полисахаридов позволяет стенке перераспределять физический стресс, обеспечивает эффективную защиту от механического и ферментативного разрушения и позволяет гифе проникнуть и расти внутри агрессивного субстрата

Функции КС • • Клеточная стенка служит каркасом для белков. Основными компонентами клеточной стенки являются полисахариды и белки (например, гликопротеины, гидрофобины, ферменты др. ). Полисахариды обеспечивают механическую прочность стенки и выполняют функцию опоры для белков, прикрепленных к ним. Белки выполняют множество очень важных функций: Гликопротеины ограничивают проницаемость стенки, создают микроокружение во внутренних слоях стенки смежных с мембраной, участвуют в узнавании: субстрата, партнеров при спаривании, субстратов – хозяев для патогенных грибов, последующей адгезии и захвата субстрата. Гидрофобины предохраняют от высыхания.
")
Методы изучения структуры КС • световая микроскопия • просвечивающая и сканирующая электронная микроскопия (иммуноцитохимия) • биохимический анализ
 и структура поверхности клетки (B) Sсhizosaccharomyces pombe")
Ультраструктура клетки (A) и структура поверхности клетки (B) Sсhizosaccharomyces pombe

Структура и состав клеточной стенки у разных грибов

Факторы, влияющие на состав и структуру КС: - таксон-специфические признаки - стадия жизненного цикла - условия культивирования
 и глубинный мицелий")
Влияние условий культивирования на композицию клеточной стенки Agaricus bisporus (поверхностный (А) и глубинный мицелий (Б) А Б

Спора шампиньона двуспорового
 Срез фрагмента клетки мицелия (n+n). (B) Срез")
Электронная микроскопия срезов клеток Ustilago maydis (A) Срез фрагмента клетки мицелия (n+n). (B) Срез дрожжевой клетки (n). Масштабные отрезки A, 0. 05 μm; B, 0. 15 μm. Заметна разная структура КC. Стенка мицелия (A) многослойна. Слой b более электронно-плотный, чем a и c. Поверхность покрыта рыхлым электронно-плотным слоем (*). Стенка дрожжевой клетки более электронно-прозрачна, и только поверхностный слой окрашен (*).

Сахара, участвующие в построении клеточной стенки грибов
 Три главные группы полимеров: глюканы, маннаны и хитин Компоненты КС")
Компоненты КС Полисахариды (90%) Три главные группы полимеров: глюканы, маннаны и хитин Компоненты КС различаются по функциям: • скелетный компонент, который определяет жесткость КС (полисахариды) • матриксный компонент – связующий материал между фибриллами (полисахариды и белки) скелетный компонент • хитин (полимеры β-1, 4 -связанных остатков N-ацетилглюкозамин); хитозан (деацетилированный хитин) • нецеллюлозный глюкан – полимеры β-1, 3 -связанных остатков глюкозы с короткими β-1, 6 -связанными боковыми веточками • целлюлоза - β-1, 4 -связанные глюканы (очень редко) матриксные компоненты. Матриксный компонент связан химическими связями со скелетными полисахаридами и может состоять из: нецеллюлозных глюканов различных типов, маннанов, хитозана, галактанов, полимеров галактозамина и белков.

• По результатам биохимических и молекулярных исследований внутренний слой клеточной стенки грибов, видимый под электронным микроскопом (скелетный компонент), представлен хитином и глюканом (кроме зигомицетов, которые содержат хитин и хитозан), а наружный слой, составляющий матриксный компонент - гликопротеинами и глюканами у мицелиальных грибов или гликопротеинами и маннанами у дрожжей

Белки, участвующие в построении клеточной стенки грибов

Белки • белковый компонент присутствует во всех слоях клеточных покровов грибов. Белки S. cerevisiae и C. albicans могут составлять от 30 до 50% от сухой массы клеточной стенки. У мицелиальных грибов – 20 -30%.

• В клеточной стенке грибов присутствуют структурные и каталитические белки, которые принимают участие в выполнении ряда функций клеточной стенки на определенных стадиях жизненного цикла грибов и в ответ на условия окружающей среды. Структурные белки прочно связаны с клеточной стенкой, а каталитические - связаны слабо и даже секретируются в среду. •
")
Роль белков в качестве структурных элементов клеточной стенки реализуется в двух направлениях: • 1) Белок может скреплять отдельные молекулы полисахаридов, с которыми он связан ковалентными связями в единую структуру. Он может служить местом закрепления других белков клеточной стенки, соединенных с ним дисульфидными или другими типами связей. • 2) Роль белка определяется его ферментативной активностью. При этом важную структурную роль играют ферменты, осуществляющие включение вновь синтезированных элементов в клеточную стенку.

• Белки принимают непосредственное участие в росте и морфогенезе, адгезии, прикреплении к субстрату, спаривании и формировании биопленки, у патогенов они выполняют роль факторов вирулентности и иммуногенов. Некоторые из функций белков зависят от укладки полипептида, другие зависят от степени гликозилирования

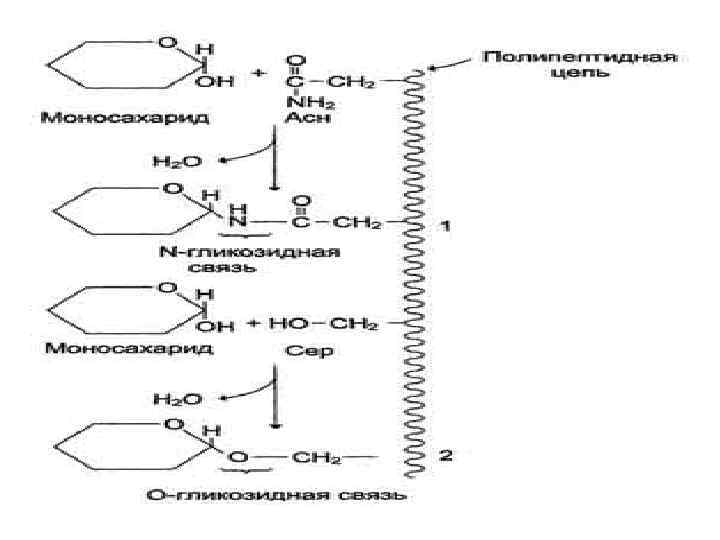
1 2 Белки клеточной стенки грибов: Большинство стеночных белков по свой природе гликопротеины, хотя есть и негликозилированные белки, такие как гликолитические ферменты, шаперон Hsp 40, гидрофобины и др. У грибов обнаружено 3 типа гликозилирования белков: 1 – О-Гликозилирование предполагает наличие О -гликозидной связи между остатком сахара и гидроксильной группой серина или треонина белка. 2 – N-Гликозилирование предполагает связывание разветвленных полисахаридных цепей с белком через хитобиозный мостик при помощи N-гликозидной связи между Nацетил-D-глюкозамином и β-амидным азотом аспарагина в белке. 3 – Белки, содержащие на С-конце гликозилфосфоинозитольный якорь (GPI якорь), который присоединяется к белку через фосфоэтаноламин.

3

• Белки клеточной стенки могут быть классифицированы согласно разным критериям, включая молекулярную структуру, функции или способ связывания с компонентами стенки. Так, белки клеточной стенки дрожжей делят на три группы, исходя из способа их экстракции.

• К первой группе относят белки, экстрагируемые при нагревании детергентами и тиоловыми реагентами, • ко второй – белки, выделяемые из клеточных стенок под действием глюканаз после удаления белков первой группы, • и к третьей - белки, выделяемые из клеточных стенок под действием щелочи после удаления белков первой группы. Такое деление отражает способ закрепления белковых молекул в клеточной стенке и отчасти позволяет сгруппировать их по характеру их функций.

• Анализ экстрагированных белков показал, что в первую группу попадают не структурные белки, а в большинстве своем ферменты, закрепленные в клеточной стенке нековалентными или дисульфидными связями. Среди ферментов значительное число гидролаз, ответственных за начальные стадии расщепления пищевых субстратов. До сих пор нет четкого представления о том, как эти ферменты встроены в клеточную стенку и как осуществляется их регуляция.

• Во второй и третьей группах находятся белки, ковалентно связанные с элементами полисахаридного каркаса клеточной стенки дрожжей. Белки второй группы связаны с β-1, 6 -глюканом клеточной стенки через фосфоэтаноламин-маннозный мостик GPI якоря. Наиболее изученным белком этой группы явлется α-агглютинин. Белки третьей группы – это белки с внутренними повторами (Pir-белки), характеризующиеся высокой степенью О-маннозилирования и отсутствием GPI якоря на С-конце. Их связь с полисахаридным каркасом осуществляется через О-связанные маннозные цепи.
 представляют собой пополняющийся")
Адгезины • Адгезины (сюда относятся белки, участвующие в процессе спаривания агглютинины) представляют собой пополняющийся постоянно класс белков клеточной адгезии, которые важны для широкого круга взаимоотношений грибной клетки с внешней/окружающей средой (Dranginis et al. , 2007). Это гликопротеины, которые секретируются в клеточную стенку, а затем закрепляются с помощью GPI якоря в цитоплазматической мембране и функционируют в поверхностном слое клеточной стенки.

Адгезины S. cerevisiae, C. albicans и C. glabrata
.")
Гидрофобины Структура гидрофобина HFBII (Armenante, 2008).

Гидрофобины • Наружная поверхность стенок гиф воздушного мицелия и спор обычно состоит из слоя палочковидных белков из семейства гидрофобинов шириной 10 нм, которые модифицируют биофизические свойства поверхности стенки. Гидрофобины принадлежат к большой и разнообразной группе родственных белков, широко представленных у грибов; при максимальной экспрессии они могут достигать 10% от общего белка стенки. Каждая молекула гидрофобина состоит из гидрофобного и гидрофильного доменов; поэтому они относятся к амфипатическим веществам (к ним же относятся фосфолипиды, составляющие мембраны клеток). Гидрофильность определяется аминогруппой. Амфипатическая структура гидрофобинов снабжает молекулы удивительным набором функций.

Гидрофобины • Молекулы гидрофобинов – относительно маленькие белки размером 7 -9 k. Da. Они содержат порядка 100 аминокислот, для них характерно присутствие сигнальной последовательности для секреции и восьми цистеиновых остатков. Эти восемь постоянных цистеиновых остатков образуют четыре дисульфидных мостика и обеспечивают правильную сборку гидрофобина. Гидрофобины не гликозилированы. Они уникальны для мицелиальных грибов, экспрессируются у представителей аскомицетов и базидиомицетов. Также имеются доказательства присутствия гидрофобинов у зигомицетов и хитридиомицетов. Каждый гриб имеет гены более одного, часто более 10 разных гидрофобинов, и эти гены обычно экспрессируются в разное время. У Schizophyllum commune гидрофобин в клеточной стенке вегетативной гифы отличается от гидрофобина, который экспрессируется в стенке гифы плодового тела.

Функции гидрофобинов • • способность гиф преодолевать поверхностное натяжение на границе раздела воздух-вода во влажных местах обитания; обеспечение гидрофобности, необходимой гифе и спорам в контакте с воздухом для защиты от излишнего испарения воды; механизм защитного действия гидрофобинов основан на свойстве самосборки на границе раздела гидрофобной/гидрофильной поверхности в амфипатическую мембрану участие в формировании репродуктивных структур (спор полового и бесполого размножения); важная роль в формировании ткани плодового тела, особенно в контролировании проницаемости для жидкости и воздуха; адгезия между гидрофильными клеточными стенками гриба и гидрофобными поверхностями растений и животных, что способствует внедрению в ткани хозяина и развитию инфекции; воздушные конидии Aspergillus, Penicillium и Cladosporium, которые имеют поверхностный слой гидрофобинов, не активируют иммунную систему; поэтому в медицине споры и гифы, покрытые слоем гидрофобинов, называют ‘иммунологически молчащей распространяющейся по воздуху плесенью’. Это означает, что хотя грибные споры повсеместны в воздухе, которым мы дышим, они никогда не активируют иммунитет и не индуцируют воспалительный процесс после вдыхания (Aimanianda et al. , 2009); опосредование симбиотических взаимоотношений гриба с корнями растений (микориза) и водорослями (лишайники).

Практическое применение гидрофобинов.


Структура и состав КС

")
Рис. И. Схема строения клеточной стенки дрожжей GPI, glycophosphatidylinositol (Selitrennikoff, 2001)

A. Интерпретация мицелиальной грибной клеточной стенки В. Интерпретация дрожжевой клеточной стенки
/капсулы sheath) Снаружи клетки грибов могут быть окружены дополнительно капсулярным материалом или чехлом.")
Чехлы (sheath)/капсулы sheath) Снаружи клетки грибов могут быть окружены дополнительно капсулярным материалом или чехлом.

• Биохимический анализ внеклеточного чехла у ряда грибов выявил присутствие полисахаридов (β-1, 6 -глюканов, β-1, 3 глюканов) и гликопротеинов, то есть по химической структуре он близок к наружному слою клеточной стенки и соответственно может быть развит в большей или меньшей степени. В последнем случае его принято рассматривать как самостоятельный слой клеточной стенки и называть чехлом или капсулой.

• При развитии чехла, окружающего клетки грибов, его наружные слои содержат белки (в том числе и гликозилированные): гидрофобины, адгезины и т. д. Капсула может иметь небольшую толщину (микрокапсула) или превышать диаметр клетки в 2 -3 раза. Размеры капсулы зависят от вида дрожжей и условий среды обитания.

• В последние годы появилось много работ по исследованию функций и строения слизистого чехла у разных грибов. • Ему приписывают роль в узнавании субстрата, адгезии, аккумуляции и сохранении компонентов клеточной стенки, модификации внеклеточного ионного окружения, сохранении от высыхания и неблагоприятных изменений окружающей среды, связывании гиф вместе, дополнительного источника питания и места локализации природных ассоциантов (например, бактерий).

• Гидрофобный внеклеточный матрикс патогена человека, возбудителя микоза легких Aspergillus fumigatus, состоит из комплекса веществ: галактоманнана, α 1, 3 -глюкана, моносахаридов, полиолов, меланинов и гидрофобинов (Beauvais et al. , 2007).
 M. Lars. et Lomb) – место локализации")
Чехлы дереворазрушающих грибов (Postia placenta (Fr. ) M. Lars. et Lomb) – место локализации экстраклеточных ферментов и грибных метаболитов

Чехол у Laetiporus sulphureus

Чехол у Pleurotus pulmonarius выполняет важную роль в межклеточных контактах

Капсула дрожжей Соединения, образующие капсулу, могут относится к следующим группам: • Фосфоманнаны – синтезируемые некоторыми аскомицетовыми дрожжами рода Pichia, образуют тонкий слой на поверхности клетки. • α-Глюканы, к числу которых относится пуллулан, образуемый дрожжеподобным грибом Aureobasidium pullulans. • β-Маннаны – образуются пигментированными дрожжами из родов Rhodotorula, Sporobolomyces. • Гетерополисахариды – характерны для многих дрожжей из родов Cryptococcus и Lipomyces. • • • Функции капсулы дрожжей: -прикрепление клеток к поверхности твердого субстрата, -резервный источник питания, -улучшение водного снабжения клетки (в том числе защита от высыхания) -в природных местах обитания – места аккумуляции спутников и ассоциантов грибов.
")
Капсула Cryptococcus neoformans играет главную роль в вирулентности Капсулярнй материал - глюкуроноксиломаннан, (88% ) (Cherniak and Sundstrom 1994).

Минорные компоненты клеточной стенки

• Липиды встречаются в клеточной стенке обычно в небольшом количестве (в основном в спорах). Липиды выполняют очень важную функцию для сохранения жизнеспособности спор (гидрофобные свойства). У спор, по сравнению с мицелием, липиды представляют менее насыщенную форму жирных кислот. Вместе с гидрофобинами липиды контролируют транспорт воды и осуществляют защиту от высыхания.

Кристаллы оксалата кальция на поверхности клеток мицелия Agaricus bisporus выделение излишних ионов Са 2+ и солей щавелевой кислоты
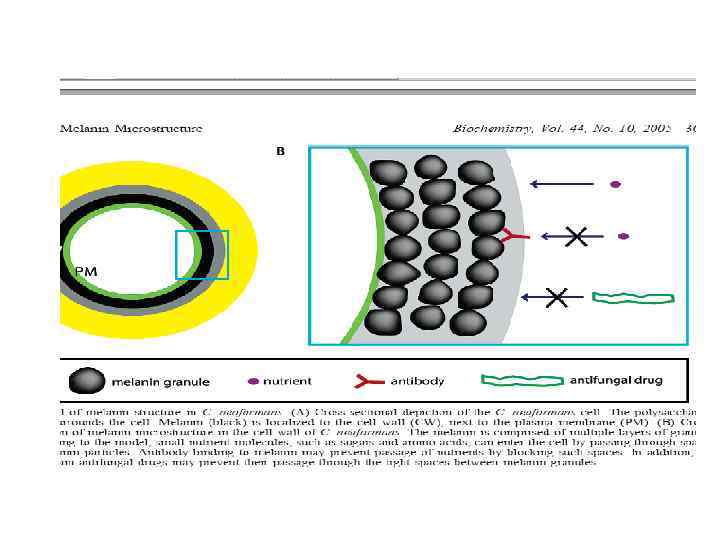
 TEM of C. neofomans a b TEM of (a)")
Меланин (защита от противогрибных препаратов) TEM of C. neofomans a b TEM of (a) melanin-lacking (500 w) and (b) melaninproducing (500 b) C. neofomans cells grown in L-DOPA medium for 7 days. Similar results were obtained with the other two pars of isolates. Scale bar, 500 nm, (Magnification X 15000).

Модель структуры меланина у C. neofomans

Функции алломеланина (продукт окисления и полимеризации безазотистых соединений фенольной природы, в его структуре присутствуют конденсированные ароматические ядра. ) : • защита от токсинов • устойчивость к экстремальным температурам • устойчивость к ультрафиолетовой радиации • защита от нематод и амеб • повышает резистентность к факторам иммунной системы • снижает восприимчивость к антигрибным препаратам

• Другой ароматический полимер, накапливающийся в оболочках спор грибов – спорополленин, (N. crassa, Mucor mucedo), который защищает споры от химических и физических факторов окружающей среды. Он представляет собой смесь биополимеров, содержащих главным образом цепи жирных кислот, фенилпропаноиды, фенольные соединения и следы каротиноидов.

Биосинтез клеточной стенки

• Биосинтез клеточной стенки грибов представляет собой комплексный процесс и происходит в трех местах: • в цитоплазме, • плазматической мембране и • самой клеточной стенке.
- и β-(1→ 3)- связанные глюканы")
• Структурные полимеры, такие как хитин, β-(1→ 4)- и β-(1→ 3)- связанные глюканы синтезируются векторно трансмембранными синтазами, локализованными в мембране, получающими предшественники в виде нуклеотидсахаров из цитозоля и снабжающими полимеризованными цепями клеточную стенку.
 синтезируются внутриклеточно и через цитоплазматический секреторный путь, включающий")
• Матриксные полимеры (глюканы, гликопротеины) синтезируются внутриклеточно и через цитоплазматический секреторный путь, включающий эндоплазматический ретикулум, везикулы Гольджи и секреторные гранулы доставляются к клеточной стенке. • Окончательная организация стенки, включающая модификацию полимеров и их ковалентное перекрестное связывание, происходят в самой стенке.
")
Биосинтез компонентов клеточной стенки. Синтез гликопротеина начинается в ЭПР (ER; белок показан фиолетовым цветом) с добавлением N-связанного олигосахарида (желтым) в процессе трансяции. GPI якорь (красным) также добавляются к некоторым белкам в ЭПР. В аппарате Гольджи, гликозилтрансферазы модифицируют белки в дальнейшем путем добавления сахаров к основным Oсвязанным олигосахаидам и к extend N-связанным олигосахаридам. Гликопротеины секретируются в пространство клеточной стенки, где они интегрируют в структуру клеточной стенки. Хитин (голубой) и глюкан (зеленыйgreen) компоненты КС синтезируются векторно на плазмалемме и продавливаются в пространство клеточной стенки в процессе синтеза. Различные компоненты стенки сшиваются вместе в простанстве КС ассоциированными с КС гликозилгидролазами и гликозилтрансферазами (основано на фиг Bowman & Free, 2006).

Структура и синтез клеточной стенки Saccharomyces cerevisiae


Синтез хитина у Saccharomyces cerevisiae

• Динамика синтеза клеточной стенки и преобразования клеточных покровов в процессе морфогенеза

Синтез клеточной стенки у большинства клеток грибов высоко поляризован; почти весь синтез хитина и глюкана происходит в верхушке гифы величиной 1 мкм. Далее пластичные вновь синтезированные стенки в верхушках грибных гиф, в апикальном куполе постепенно созревают, превращаясь в жесткие боковые стенки гиф. Процесс созревания стенок идет в трех направлениях: • перекрестное связывание различных полимеров. Наиболее заметно образование ковалентных связей между хитином и разветвленным β-(1, 3)-глюканом; • процесс кристаллизации хитиновых микрофибрилл за счет увеличения числа водородных связей; • добавление нового материала, ведущее к утолщению микрофибрилл, и накопление матриксных материалов, покрывающих фибриллярный скелет.

• В процессе морфогенеза грибов происходят качественные и количественные преобразования клеточных покровов. Эти изменения определяют транспорт питательных веществ, метаболизм непроницаемых для клеточной стенки веществ и в большой степени зависят от функционального статуса клетки (клетка вегетативного мицелия, покоящиеся споры и т. д. ).

Мицелиально-дрожжевой диморфизм


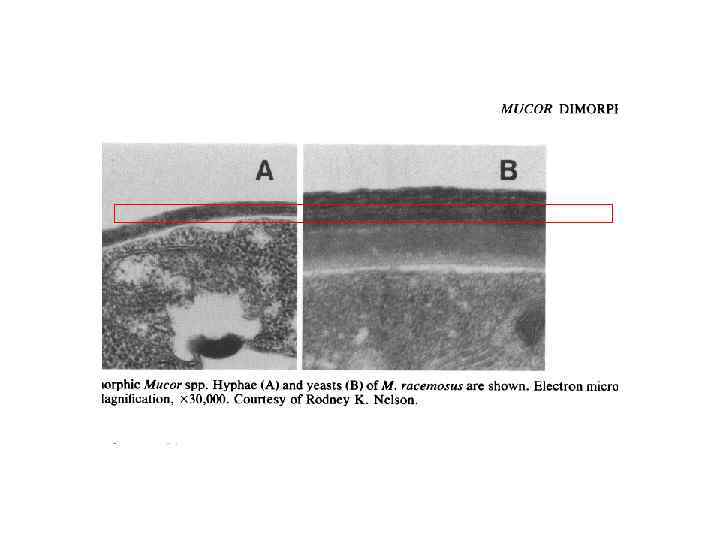
 Michalenko G.")
Очищенная фракция бесцветных клеточных оболочек вегетативного мицелия шампиньона содержит (% сух вес) Michalenko G. O. , Hohle H. R. , Rost D. Chemistry and architecture of the mycelial wall of Agaricus bisporus // J. Gen. Microbiol. 1976. V. 92. № 2. P. 251 -262. Таблица Компоненты клеточной стенки мицелий споры хитин 43 5 щелоченераствори мый глюкан 26 - щелочерастворим ый глюкан 14 - белок 16 26 липиды 15 13 следы - меланин - 41 хитозан - 15 ксилломаннан

Стресс и компенсаторные механизмы

3 2 1

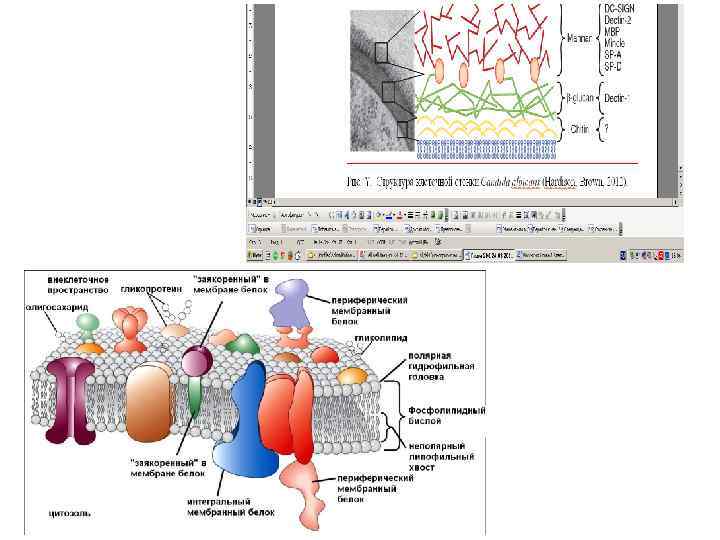
Цитоплазматическая мембрана

Цитоплазматическая мембрана
. «Жидкая мозаичная модель структуры")
Цитоплазматическая мембрана Singer SJ, Nicolson GL (февраль 1972 г. ). «Жидкая мозаичная модель структуры клеточных мембран»
, сфингомиелином (SMs) и холестерином (эргостерол")
Цитоплазматическая мембрана Наружный слой липидного моста обогащен гликосфинголипидами (GSLs), сфингомиелином (SMs) и холестерином (эргостерол у грибов), а внутренний слой обогащен насыщенными глицерофосфолипидами (GPLs). Обычная мембрана обогащена ненасыщенными глицерофосфолипидами GPLs.


• Важными компонентами грибных мембран являются стеролы, они служат для регуляции текучести мембран, активности мембран-связанных ферментов и транспортных механизмов. • У растений наиболее распространенные стеролы плазмалеммы - сигмастерол, ситостерол и кампестерол; • У клеток животных - холестерол • Пероноспоровые (из оомицетов/псевдогрибов) вообще не способны к синтезу стеролов и используют стеролы растений хозяев, а сапролегниевые (представители оомицетов) имеют фукостерол, как и бурые водоросли и ланостерол (общий для грибов и животных). • В то время как у большинства грибов главный стерол плазмалеммы - эргостерол (за исключением Chytridiomycota, у которых доминантный стерол холестерол).

Функции плазмалеммы Контроль поглощения и секреции веществ • В плазмалемме находятся множество транспортных систем: ионные каналы, белки-переносчики и ионные насосы, которые участвуют в селективном транспорте ионов внутрь клетки и наружу. Через плазмалемму происходит также перенос макромолекул. Так, в периплазматическое пространство транспортируются строительные блоки клеточной стенки — полисахариды и структурные белки. Эти соединения обычно находятся в везикулах аппарата Гольджи и высвобождаются из них путем экзоцитоза. Запасание и использование энергии • Плазмалемма любой клетки является энергизованной мембраной, т. е. на ней существует градиент электрохимического потенциала ΔμH+, который используется для выполнения полезной работы, прежде всего для активного переноса веществ через мембрану. Размещение и обеспечение работы ферментов • В плазмалемме находится множество ферментов: ферменты построения клеточной стенки (хитинсинтаза, глюкансинтаза), ферменты сигнальных систем и др. Активность хитинсинтазы, которая очень высокая в очищенных препаратах плазматической мембраны грибов, используется как маркер плазматической мембраны при субклеточном фракционировании.

Функции плазмалеммы Рецепторные функции • Многие белки плазмалеммы являются рецепторами различных сигналов. Плазмалемма клет-и — это комплекс различных рецепторов эндогенных сигналов и внешних воздействий (рецепторы физических факторов — температуры, соли, р. Н и др. ) Температура и засоленность изменяют текучесть мембраны. Изменение этих свойств плазмалеммы приводит к открытию или закрытию находящихся в ней механосенсорных ионных каналов. Таким образом, плазмалемма является местом рецепции (восприятия) различных сигналов химической и физической природы. Сигнальная функция • Многие компоненты плазмалеммы после восприятия сигналов служат источником вторичных мессенджеров — молекул, которые «передают» сигнал по эстафете и усиливают его. В качестве вторичных мессенджеров выступают инозитол-1, 4, 5 -трифосфат, диацилглицерин, фосфатидная кислота, продукты перекисного окисления липидов мембран. Все эти соединения образуются из липидов плазмалеммы под действием специальных ферментов, активируемых в результате воздействия сигнала на рецепторы. Таким образом, плазматическая мембрана является местом не только рецепции сигналов, но также их усиления и дифференцирования.
Лекция 1-2017 клеточная стенка .ppt