Лекция 1- end 2017 клеточная стенка .ppt
- Количество слайдов: 20



Цитология грибной клетки

• Динамика синтеза клеточной стенки и преобразования клеточных покровов в процессе морфогенеза

Синтез клеточной стенки у большинства клеток грибов высоко поляризован; почти весь синтез хитина и глюкана происходит в верхушке гифы величиной 1 мкм. Далее пластичные вновь синтезированные стенки в верхушках грибных гиф, в апикальном куполе постепенно созревают, превращаясь в жесткие боковые стенки гиф. Процесс созревания стенок идет в трех направлениях: • перекрестное связывание различных полимеров. Наиболее заметно образование ковалентных связей между хитином и разветвленным β-(1, 3)-глюканом; • процесс кристаллизации хитиновых микрофибрилл за счет увеличения числа водородных связей; • добавление нового материала, ведущее к утолщению микрофибрилл, и накопление матриксных материалов, покрывающих фибриллярный скелет.

• В процессе морфогенеза грибов происходят качественные и количественные преобразования клеточных покровов. Эти изменения определяют транспорт питательных веществ, метаболизм непроницаемых для клеточной стенки веществ и в большой степени зависят от функционального статуса клетки (клетка вегетативного мицелия, покоящиеся споры и т. д. ).

Мицелиально-дрожжевой диморфизм

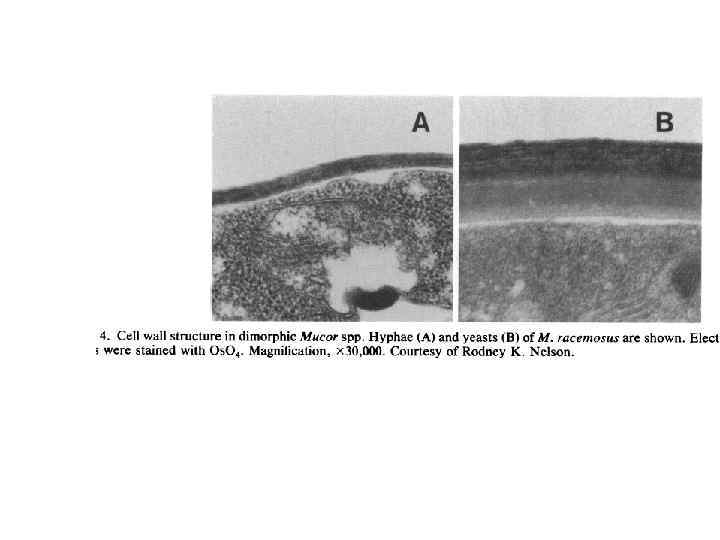
 Michalenko G.")
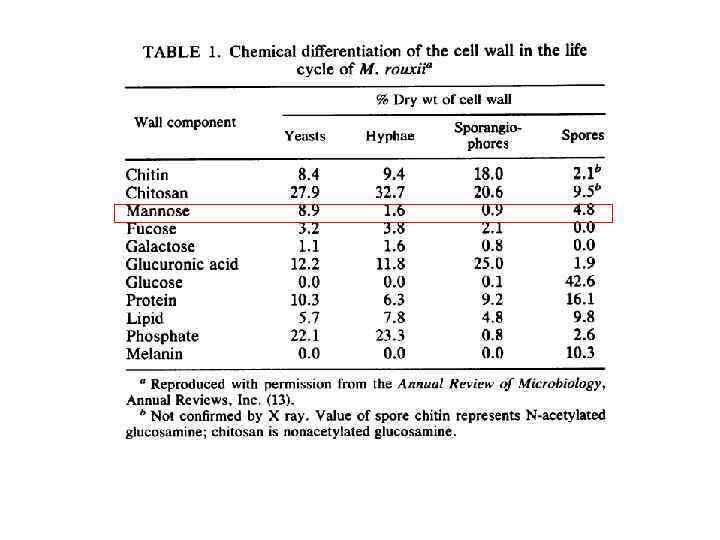
Очищенная фракция бесцветных клеточных оболочек вегетативного мицелия шампиньона содержит (% сух вес) Michalenko G. O. , Hohle H. R. , Rost D. Chemistry and architecture of the mycelial wall of Agaricus bisporus // J. Gen. Microbiol. 1976. V. 92. № 2. P. 251 -262. Таблица Компоненты клеточной стенки мицелий споры хитин 43 5 щелоченераствори мый глюкан 26 - щелочерастворим ый глюкан 14 - белок 16 26 липиды 15 13 следы - меланин - 41 хитозан - 15 ксилломаннан
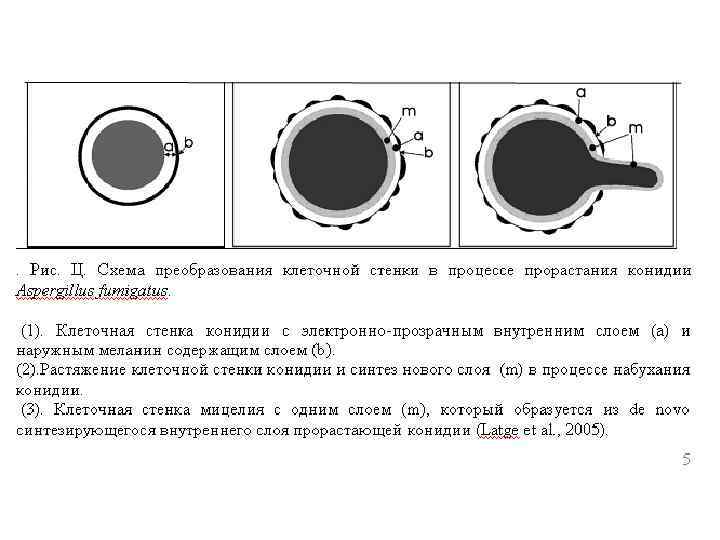

Стресс и компенсаторные механизмы


3 2 1

Цитоплазматическая мембрана

Цитоплазматическая мембрана
")
Жидкая мозаичная модель клеточной мембраны Singer SJ, Nicolson GL (1972 г)
, сфингомиелином")
Цитоплазматическая мембрана. Модель, имеющая липидный мост Наружный слой липидного моста обогащен гликосфинголипидами (GSLs), сфингомиелином (SMs) и холестерином (эргостерол у грибов), а внутренний слой обогащен насыщенными глицерофосфолипидами (GPLs). Обычная мембрана обогащена ненасыщенными глицерофосфолипидами GPLs.

• Важными компонентами грибных мембран являются стерины, они служат для регуляции текучести мембран, активности мембран-связанных ферментов и транспортных механизмов. • У растений наиболее распространенные стерин плазмалеммы - сигмастерин, ситостерин и кампестерин; • У клеток животных - холестерин • Пероноспоровые (из оомицетов/псевдогрибов) вообще не способны к синтезу стеринов и используют стерины растений хозяев, а сапролегниевые (представители оомицетов) имеют фукостерин, как и бурые водоросли и ланостерин (общий для грибов и животных). • В то время как у большинства грибов главный стерин плазмалеммы - эргостерин (за исключением Chytridiomycota, у которых доминантный стерин холестерин).

Функции плазмалеммы Контроль поглощения и секреции веществ • В плазмалемме находятся множество транспортных систем: ионные каналы, белки-переносчики и ионные насосы, которые участвуют в селективном транспорте ионов внутрь клетки и наружу. Через плазмалемму происходит также перенос макромолекул. Так, в периплазматическое пространство транспортируются строительные блоки клеточной стенки — полисахариды и структурные белки. Эти соединения обычно находятся в везикулах аппарата Гольджи и высвобождаются из них путем экзоцитоза. Запасание и использование энергии • Плазмалемма любой клетки является энергизованной мембраной, т. е. на ней существует градиент электрохимического потенциала ΔμH+, который используется для выполнения полезной работы, прежде всего для активного переноса веществ через мембрану. Размещение и обеспечение работы ферментов • В плазмалемме находится множество ферментов: ферменты построения клеточной стенки (хитинсинтаза, глюкансинтаза), ферменты сигнальных систем и др. Активность хитинсинтазы, которая очень высокая в очищенных препаратах плазматической мембраны грибов, используется как маркер плазматической мембраны при субклеточном фракционировании.

Функции плазмалеммы Рецепторные функции • Многие белки плазмалеммы являются рецепторами различных сигналов. Плазмалемма клетки — это комплекс различных рецепторов эндогенных сигналов и внешних воздействий (рецепторы физических факторов — температуры, соли, р. Н и др. ) Температура и засоленность изменяют текучесть мембраны. Изменение этих свойств плазмалеммы приводит к открытию или закрытию находящихся в ней механосенсорных ионных каналов. Таким образом, плазмалемма является местом рецепции (восприятия) различных сигналов химической и физической природы. Сигнальная функция • Многие компоненты плазмалеммы после восприятия сигналов служат источником вторичных мессенджеров — молекул, которые «передают» сигнал по эстафете и усиливают его. В качестве вторичных мессенджеров выступают инозитол-1, 4, 5 -трифосфат, диацилглицерин, фосфатидная кислота, продукты перекисного окисления липидов мембран. Все эти соединения образуются из липидов плазмалеммы под действием специальных ферментов, активируемых в результате воздействия сигнала на рецепторы. Таким образом, плазматическая мембрана является местом не только рецепции сигналов, но также их усиления и дифференцирования.
Лекция 1- end 2017 клеточная стенка .ppt