Цитогенетика 16-18.ppt
- Количество слайдов: 52



Цитогенетика НГУ 2012 Рубцов Николай Борисович, д. б. н. , зав. лабораторией, ИЦи. Г СО РАН.

Краткая система описания структурных хромосомных аномалий. При описании хромосомных перестроек с двумя точками разрывов в одной хромосоме первыми указывается точка разрыва в р-плече, или если они локализованы в одном плече, то проксимальная точка разрыва: 46, XX, inv(2)(p 21 q 31); 46, XX, inv(2)(p 13 p 23).

Структурные аномалии. Хромосома-донор всегда стоит на последнем месте, даже если это половая хромосома или хромосома с более низким номером: 46, XX, ins(5; X)(p 14; q 21 q 25) означает инсерцию фрагмента Х хромосомы (q 21→q 25) в бэнд р14 хромосомы 5. 46, XX, ins(5; 2)(p 14; q 21 q 25) - инсерция фрагмента (q 21→q 25) хромосомы 2 в бэнд р14 хромосомы 5. Если подобная перестройка затрагивает только одну хромосому, то сначала указывается бэнд вставки, затем, в случае прямой инсерции, более близкая к центромере точка разрыва фрагмента инсерции и, наконец, дистальная точка разрыва этого фрагмента: 46, XX, ins(2)(q 13 p 23) Запись 46, XX, ins(2)(q 13 p 23 p 13), в отличие от предыдущей, означает вставку фрагмента p 13→p 23 в противоположной ориентации.

Для транслокаций, в которые вовлечены три хромосомы при одной точке разрыва в каждой из них, существует следующий порядок перечисления хромосом: · первой приводится половая хромосома или хромосома с наименьшим номером; · затем хромосома-реципиент фрагмента хромосомы, поставленной первой; · и, наконец, хромосома, являющаяся донором фрагмента первой хромосомы
(q 34; q 11; q 22) означает, что")
Расшифровка записи 46, XX, t(9; 22; 17)(q 34; q 11; q 22) означает, что фрагмент хромосомы 9, соответствующий району дистальному относительно 9 q 34, перенесен на хромосому 22 с точкой разрыва-соединения в 22 q 11, фрагмент хромосомы 22, соответствующий району дистальному относительно 22 q 11, перенесен на хромосому 17 с точкой разрыва-соединения в 17 q 22, и, наконец, фрагмент хромосомы 17, соответствующий району дистальному относительно 17 q 22, перенесен на хромосому 9 с точкой разрыва-соединения в 9 q 34.

Полная система описания структурных хромосомных аномалий. Описание перестроенной хромосомы начинается с конца рплеча или ближайшего к нему бэнда и продолжается до конца q-плеча. Бэнды указываются в том порядке, в котором они располагаются в хромосоме. Если перестроенная хромосома содержит материал одной хромосомы, то номер хромосомы при обозначении бэндов не повторяется. В следующих разделах приведены примеры различных хромосомных перестроек.
(q 22 q 25), 46, XX, dup(1)(pter→q 25: : q 22→qter) –")
46, XX, dup(1)(q 22 q 25), 46, XX, dup(1)(pter→q 25: : q 22→qter) – прямая дупликация, 46, XX, dup(1)(q 25 q 22), 46, XX, dup(1)(pter→q 25: : q 25→q 22: : q 25→qter) или 46, XX, dup(1)(pter→q 22: : q 25→q 22: : q 22→qter) – инвертированная дупликация. 46, XX, ins(2)(p 13 q 21 q 31), 46, XX, ins(2)(pter→p 13: : q 31→q 21: : p 13→q 21: : q 31→qter) или 46, XX, dir ins(2)(pter→p 13: : q 31→q 21: : p 13→q 21: : q 31→qter). 46, XX, ins(2)(p 13 q 31 q 21), 46, XX, ins(2)(pter→p 13: : q 31→q 21: : p 13→q 21: : q 31→qter) или 46, XX, dir ins(2)(pter→p 13: : q 21→q 31: : p 13→q 21: : q 31→qter).
(q 13) (краткая система записи), 46, XX, del(5)(pter→q 13: ) (полная")
Делеции: 46, XX, del(5)(q 13) (краткая система записи), 46, XX, del(5)(pter→q 13: ) (полная система записи). Потерян район q 13→qter. 46, XX, del(5)(q 13 q 33) (краткая система записи), 46, XX, del(5)(petr→q 13: : q 33→qter) (полная система записи). Потерян район q 13→q 33. Инверсии: 46, XX, inv(3)(q 21 q 26); 46, XX, inv(3)(pter→q 21: : q 26→qter - парацентирическая инверсия района 3 q 21→q 26. 46, XX, inv(3)(p 13 q 21); 46, XX, inv(3)(pter→p 13: : q 21→qter) перицентрическая инверсия района 3 p 13 q 21
t(1; 3)(p 22; q 13) (краткая система")
Примеры описания производных хромосом: 46, XX, der 1(1)t(1; 3)(p 22; q 13) (краткая система записи), 46, XX, der 1(1)t(1; 3)(1 qter→ 1 p 22: : 3 q 13→ 3 qter) (полная система записи). der(1)t(1; 3)(p 32; q 21)t(1; 11)(q 25; q 13) (краткая система записи), der(1)t(3 qter→ 3 q 21: : 1 p 32→ 1 q 25: : 11 q 13→ 11 qter) (полная система записи). der(1)t(1; 3)(p 32; q 21)t(3; 7)(q 28; q 11), der(1)(1 qter→ 1 p 32: : 3 q 21→ 3 q 28: : 7 q 11→ 7 qter). der(9)del(9)(p 12)t(9; 11)(q 34; q 11), der(9)(: 9 p 12→ 9 q 34: : 13 q 11→ 13 qter).
 растянутого (распластанного) хроматина или ДНК;")
fib ish - гибридизация in situ c нитями (волокнами) растянутого (распластанного) хроматина или ДНК; fish - флуоресцентная гибридизация in situ; nuc ish - флуоресцентная гибридизация in situ с ядрами интерфазных клеток; pcp - районоспецифические ДНК-пробы, полученные методом микродиссекции метафазных хромосом или представляющие собой контиги клонированных фрагментов ДНК (partitial chromosome paint); rev ish - обратная гибридизация in situ (reverse in situ hybridization) - гибридизация in situ ДНК-проб, приготовленных из анализируемого материала, с нормальными хромосомами человека; wcp - хромоспецифичные ДНК-пробы (whole chromosome paint).
(q 11. 2)(D 22 S 75 -) -")
Порядок использования «ish номенклатуры» : ish del(22)(q 11. 2)(D 22 S 75 -) - рутинный цитогенетический анализ не проводился, но делеция суббэнда 22 q 11. 2, характерная для синдрома Di. George, была выявлена проведением гибридизации in situ ДНК-пробы локуса D 22 S 75. ish del(22)(q 11. 2)(D 22 S 75 -x 2) рутинный цитогенетический анализ не проводился, но делеция суббэнда 22 q 11. 2 была выявлена в обоих гомологах хромосомы 22 проведением гибридизации in situ ДНК-пробы локуса D 22 S 75. 46, XX, ish der(4)t(4; 11)(p 16. 3; p 15)mat(wcp 11+, D 4 F 26 -, D 4 S 96+, D 4 Z 1+) девочка с одной нормальной хромосомой 4, двумя нормальными хромосомами 11 и хромосомой der(4)t(4; 11)(p 16. 3; p 15), унаследованной от матери, являющейся носителем сбалансированной t(4; 11). Хромосома der(4)t(4; 11)(p 16. 3; p 15) была детально описана благодаря использованию гибридизации in situ ДНК-проб wcp 11, D 4 F 26, D 4 S 96 и D 4 Z 1.

Краткая система описания структурных хромосомных аномалий. При описании хромосомных перестроек с двумя точками разрывов в одной хромосоме первыми указывается точка разрыва в р-плече, или если они локализованы в одном плече, то проксимальная точка разрыва: 46, XX, inv(2)(p 21 q 31); 46, XX, inv(2)(p 13 p 23).

Структурные аномалии. Хромосома-донор всегда стоит на последнем месте, даже если это половая хромосома или хромосома с более низким номером: 46, XX, ins(5; X)(p 14; q 21 q 25) означает инсерцию фрагмента Х хромосомы (q 21→q 25) в бэнд р14 хромосомы 5. 46, XX, ins(5; 2)(p 14; q 21 q 25) - инсерция фрагмента (q 21→q 25) хромосомы 2 в бэнд р14 хромосомы 5. Если подобная перестройка затрагивает только одну хромосому, то сначала указывается бэнд вставки, затем, в случае прямой инсерции, более близкая к центромере точка разрыва фрагмента инсерции и, наконец, дистальная точка разрыва этого фрагмента: 46, XX, ins(2)(q 13 p 23) Запись 46, XX, ins(2)(q 13 p 23 p 13), в отличие от предыдущей, означает вставку фрагмента p 13→p 23 в противоположной ориентации.

Для транслокаций, в которые вовлечены три хромосомы при одной точке разрыва в каждой из них, существует следующий порядок перечисления хромосом: · первой приводится половая хромосома или хромосома с наименьшим номером; · затем хромосома-реципиент фрагмента хромосомы, поставленной первой; · и, наконец, хромосома, являющаяся донором фрагмента первой хромосомы
(q 34; q 11; q 22) означает, что")
Расшифровка записи 46, XX, t(9; 22; 17)(q 34; q 11; q 22) означает, что фрагмент хромосомы 9, соответствующий району дистальному относительно 9 q 34, перенесен на хромосому 22 с точкой разрыва-соединения в 22 q 11, фрагмент хромосомы 22, соответствующий району дистальному относительно 22 q 11, перенесен на хромосому 17 с точкой разрыва-соединения в 17 q 22, и, наконец, фрагмент хромосомы 17, соответствующий району дистальному относительно 17 q 22, перенесен на хромосому 9 с точкой разрыва-соединения в 9 q 34.

Полная система описания структурных хромосомных аномалий. Описание перестроенной хромосомы начинается с конца рплеча или ближайшего к нему бэнда и продолжается до конца q-плеча. Бэнды указываются в том порядке, в котором они располагаются в хромосоме. Если перестроенная хромосома содержит материал одной хромосомы, то номер хромосомы при обозначении бэндов не повторяется. В следующих разделах приведены примеры различных хромосомных перестроек.
(q 22 q 25), 46, XX, dup(1)(pter→q 25: : q 22→qter) –")
46, XX, dup(1)(q 22 q 25), 46, XX, dup(1)(pter→q 25: : q 22→qter) – прямая дупликация, 46, XX, dup(1)(q 25 q 22), 46, XX, dup(1)(pter→q 25: : q 25→q 22: : q 25→qter) или 46, XX, dup(1)(pter→q 22: : q 25→q 22: : q 22→qter) – инвертированная дупликация. 46, XX, ins(2)(p 13 q 21 q 31), 46, XX, ins(2)(pter→p 13: : q 31→q 21: : p 13→q 21: : q 31→qter) или 46, XX, dir ins(2)(pter→p 13: : q 31→q 21: : p 13→q 21: : q 31→qter). 46, XX, ins(2)(p 13 q 31 q 21), 46, XX, ins(2)(pter→p 13: : q 31→q 21: : p 13→q 21: : q 31→qter) или 46, XX, dir ins(2)(pter→p 13: : q 21→q 31: : p 13→q 21: : q 31→qter).
(q 13) (краткая система записи), 46, XX, del(5)(pter→q 13: ) (полная")
Делеции: 46, XX, del(5)(q 13) (краткая система записи), 46, XX, del(5)(pter→q 13: ) (полная система записи). Потерян район q 13→qter. 46, XX, del(5)(q 13 q 33) (краткая система записи), 46, XX, del(5)(petr→q 13: : q 33→qter) (полная система записи). Потерян район q 13→q 33. Инверсии: 46, XX, inv(3)(q 21 q 26); 46, XX, inv(3)(pter→q 21: : q 26→qter - парацентирическая инверсия района 3 q 21→q 26. 46, XX, inv(3)(p 13 q 21); 46, XX, inv(3)(pter→p 13: : q 21→qter) перицентрическая инверсия района 3 p 13 q 21
t(1; 3)(p 22; q 13) (краткая система")
Примеры описания производных хромосом: 46, XX, der 1(1)t(1; 3)(p 22; q 13) (краткая система записи), 46, XX, der 1(1)t(1; 3)(1 qter→ 1 p 22: : 3 q 13→ 3 qter) (полная система записи). der(1)t(1; 3)(p 32; q 21)t(1; 11)(q 25; q 13) (краткая система записи), der(1)t(3 qter→ 3 q 21: : 1 p 32→ 1 q 25: : 11 q 13→ 11 qter) (полная система записи). der(1)t(1; 3)(p 32; q 21)t(3; 7)(q 28; q 11), der(1)(1 qter→ 1 p 32: : 3 q 21→ 3 q 28: : 7 q 11→ 7 qter). der(9)del(9)(p 12)t(9; 11)(q 34; q 11), der(9)(: 9 p 12→ 9 q 34: : 13 q 11→ 13 qter).

Элементы хромосомы

Структурные единицы организации хромосомы Теломера Теломерные повторы Теломера-ассоциированные повторы Эухроматиновые районы: G- и R-бэнды Центромерный район Центромера Прицентромерный гетероматин Эухроматиновые районы: G- и R-бэнды Теломера Теломерные повторы Теломера-ассоциированные повторы Ядрышкообразующие районы Кластеры повторенных последовательностей Кластеры дуплицированных последовательностей

Взаимоотношения между G-, R-, C- и Т-бэндами Эухроматиновые районы: G- и R-бэнды Бэнд. Что это? Бэнды: время репликации или изохоры? Насколько велики различия между структурными элементами в пределах одного бэнда? Что такое граница между бэндами? Т-бэнды и гигантские «блуждающие» петли. Случайность и закономерность локализации хромосом в ядре, бэндов в хромосоме. Т-? -? -? -R-? -? -? -G

Nokkala 1994 Rattner 1992

Изохоры ДНК «Геном млекопитающих организован как мозаика изохор, протяженных участков ДНК с определенным нуклеотидным составом. Изохоры представляют собой основу хромосомного бэндинга, связанного с многочисленными структурными и функциональными характеристиками хромосомных районов. Мы недавно показали, что переход от GC-бедной изохоры к GC-богатой в районе гена NF 1 (neurofibromatosis 1) происходит в пределах 5 kb и демаркируют геномные районы с высокой и низкой рекомбинационной частотой. В настоящей статье мы показали, что та же переходная зона разделяет рано и поздно реплицирующийся хроматин. В зоне перехода изохор репликационная вилка задерживается в средней S фазе и может быть визуализована fiber-FISH как Y-структура. Смена изохор консервативна при сравнении геномов человека и мыши, что подчеркивает важность переходных зон в качестве знаков в структурной организации хромосомных районов и определения их функциональной активности» . Genomics xx (2005) xxx – xxx An isochore transition zone in the NF 1 gene region is a conserved landmark of chromosome structure and function Claudia Schmegner et al.
 GC состав района NF 1 гена человека")
Границы «структурных элементов» и изохоры ДНК (a) GC состав района NF 1 гена человека (синяя) и мыши (красная). Ген NF 1 беден GC у обоих видов (среденее значение GC NF 1 гена у человека - 37%, у мыши- 39%). В соседних районах (гены RAB 11 -FIP 4) оно выше (~ 51%). Район перехода занимает около 10 kb. Его локализация идентична у обоих видов. Космиды, использованные для интерфазной FISH (A–F) и фибер -FISH (G, H) показаны ниже. Результаты FISH с интерфазными ядрами лимфоцитов человека (c, d) и фибробластов мыши (b, e). Окраска ДНК DAPI. Красным показана ДНК, включившая Budr в процессе репликации. Двойные сигналы FISH (зеленые) указывают на прошедшую репликацию (d, e), одиночные на непрошедшую (b, c). (f) Процент с непрошедшей репликацией соответствующего фрагмента ДНК. Известно, что PYGM ген реплицируется Genomics xx (2005) xxx – xxx раньше, а ген CFTR – позже гена An isochore transition zone in the NF 1 gene region is a NF 1. conserved landmark of chromosome structure and function Claudia Schmegner et al.
")
Границы «структурных элементов» и изохоры ДНК Структурный анализ репликационной вилки в транзитной зоне. (a-d) лимфоциты человека. (e-g) фибробласты мыши. (а, е) двухцветный FISH ДНК-проб, фланкирующих переходную зону, с интерфазными ядрами синхронизированных клеток при альтернативных вариантах мечения. (b-d, f, g) fiber-FISH с распластанным хроматином. (b, f, g) fiber-FISH с распластанным хроматином ДНК-проб, перекрывающих переходную зону. (c, d) fiber-FISH с распластанным хроматином, фланкирующих переходную зону: (с) GC-богатая проба- зеленый сигнал, GC-бедная проба – красный сигнал; (d) GC- богатая проба- красный сигнал, GC -бедная проба- зеленый сигнал. Genomics xx (2005) xxx – xxx An isochore transition zone in the NF 1 gene region is a conserved landmark of chromosome structure and function Claudia Schmegner et al.
: RESEARCH 0025.")
PNAS, vol. 102, no. 18: 6419– 6424, 2005 Genome Biol. 2001; 2(7): RESEARCH 0025. Temporal profile of replication of human chromosomes A draft annotation and overview of the human genome Yesu Jeon et al. F. A. Wright et al.
 Отсутствие корреляции между G-бэндами и GC-бедными районами, R-бэндами и")
Реконструкция G-бэндинга in silico (A) Отсутствие корреляции между G-бэндами и GC-бедными районами, R-бэндами и GCбогатыми районами хромосом 21 (слева) и 22 (справа). Значение содержания GC оценивали в 2. 5 Mb районе и сравнивали со средним по хромосоме (черная линия показывает GC пар при шаге 10 -kb). (B) Корреляции между G-бэндами и in silico бэндами хромосом 21 (слева) и 22 (справа). In silico Gбэнды рассчитывали как превышение процента GC пар в окружающем в 2, 5 -Mb районе над процентом над GC пар в окружающем в 9, 3 -Mb районе. (C) Наилучшее соответствие между G-бэндами и in silico Gбэндами было обнаружено для 3 q. PNAS, 2002, v 99, n 2: 797– 802 In silico chromosome staining: Reconstruction of Giemsa bands from the whole human genome sequence Yoshihito Niimura and Takashi Gojobori

Закономерности локализации хромосомных территорий Хромосомные территории и межхроматиновое пространство Рано и поздно реплицирующиеся хромосомные домены. a ) Средний оптический срез через ядро клетки нейробластомы SH-EP N 14, фиксированной через 20 часов после двухцветного мечения ДНК Cy 3 - и Cy 5 коньюгированных с нуклеотидами (синий сигнал – начало S -фазы, красный – середина S-фазы). b, c ) Средний оптический срез через то же ядро клетки нейробластомы с визуализацией ламина В (зеленый). Nat Rev Genet. 2001 Apr; 2(4): 292 -301 CHROMOSOME TERRITORIES, NUCLEAR ARCHITECTURE AND GENE REGULATION IN MAMMALIAN CELLS T. Cremer and C. Cremer

Локализация ДНК в метафазной хромосоме и интерфазном ядре

Владивосток, 29. 04. 2009

Топология хромосомных территорий и регуляция транскрипционной активности генов


Организация центромеры

Определения Исходно центромерой считалось место первичной перетяжки хромосомы. И в настоящее время этот пункт входит во многие определения, которые можно найти в статьях, словарях и в интернете ( «место первичной перетяжки в хромосоме» ; www. mhhe. com/biosci/genbio/life/glossaryc. mhtml). Можно найти много других определений, которые будут от этого: l специализированный район конденсированного хроматина в каждой из хромосом, в котором во время митоза сохраняется контакт хроматид, создавая Х-образную форму хромосомы; «центромеру сложно секвенировать» (http: //wordnetweb. princeton. edu/perl/webwn); l центромера – район ДНК обычно расположенный недалеко от центра хромосомы, в котором сестринские хроматиды находятся в контакте. Он вовлечен в клеточное деление, как место крепления митотического веретена (http: //en. wikipedia. org/wiki/Centromere); l район эукариротической хромосомы, где собирается кинетохор; l район первичной перетяжки, включающий микротрубочки веретена, ответственный за движение хромосом во время митоза и мейоза (http: //biology. usgs. gov/genetics_genomics/glossary_c. html); l перетяжка в хромосоме, делящая ее на плечи, служит местом соединения сестринских хроматид и прикрепления нитей веретена www. biologylessons. sdsu. edu/classes/lab 8/glossary. html l место на хромосоме, где соприкасаются хроматиды, и крепятся микротрубочки. www. knowledgerush. com/kr/encyclopedia/Chromosome/ l район перетяжки, соединяющий две сестринские хроматиды, обуславливающий Х-форму хромосомы, место формирования кинетохора. www. biology-online. org/dictionary/Centromere
: центромера – «локус, состоящий из хроматина,")
l Шулер и Сулливан (Schueler and Sullivan, 2006): центромера – «локус, состоящий из хроматина, необходимого для правильной сегрегации хромосом» , - или более подробное: «место сборки кинетохора, белковой структуры, присутствующей на всех хромосомах, координирующей прикрепление и их движение вдоль микротубочек» . l Подгорная с коллегами (2009): центромера – «это структура, обеспечивающая удержание хромосом, правильность выстраивания хромосом в метафазной пластинке и их прикрепление к веретену; участок, ответственный за контроль наступления анафазы» .
 l центромера определена как «участок хромосомы, характеризующийся специфической последовательностью")
«Академик» (www. academic. ru) l центромера определена как «участок хромосомы, характеризующийся специфической последовательностью нуклеотидов и структурой. Центромера играет важную роль в процессе деления клеточного ядра и в контроле экспрессии генов» , но уже через пару абзацев написано, что «у большинства эукариот центромера не имеет определённой, соответствующей ей нуклеотидной последовательности» . l Вероятно, наиболее просто было бы определить центромеру, как участок ДНК, на которой происходит сборка кинетохора, и отметить при этом, что последовательность нуклеотидов этого участка не является ни достаточным, ни необходимым элементом для проявления активной центромеры.

Определения Шулера и Сулливана связанных с центромерой или описывающих часть ее элементов. l Кинетохор: белковая структура, собранная на центромерном хроматине, связывающая ДНК центромеры с протеинами, управляющими движением хромосомы и определяющими прикрепление хромосомы к микротрубочкам веретена. l Центромерный район: район, включающий различные домены, расположенные рядом с центромерным локусом, имеющие соответствующие функции. l α-сателлит: семейство сателиттной ДНК на основе повторенной единицы размером 171 пар нуклеотидов, представленной в центромере у всех исследованных видов приматов. l Мономер: наименьшая по размеру единица повторенной сателлитной ДНК. l Повтор более высокого порядка: единица повторенной сателлитной ДНК, состоящая из набора копий сателлитных мономеров. l Центромерный хроматин: специализированный хроматин в центромерном районе, являющийся основой кинетохора.
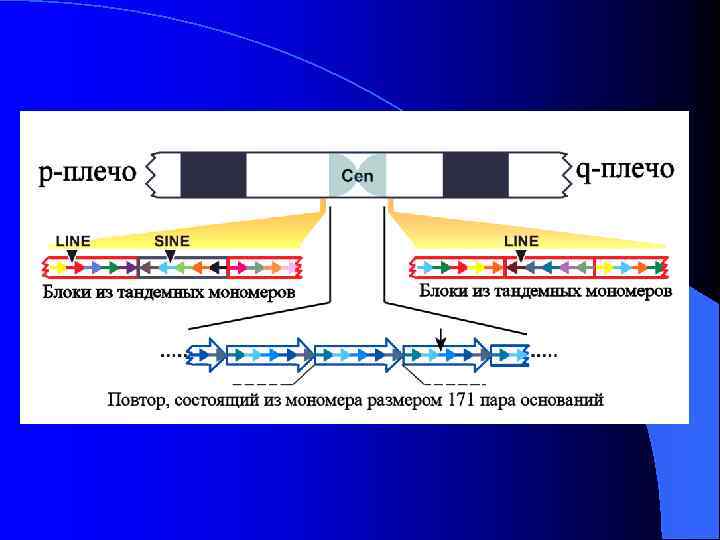
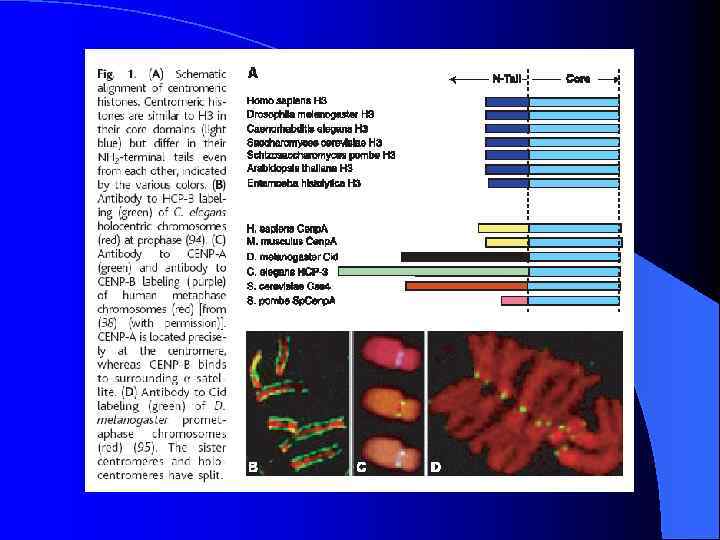
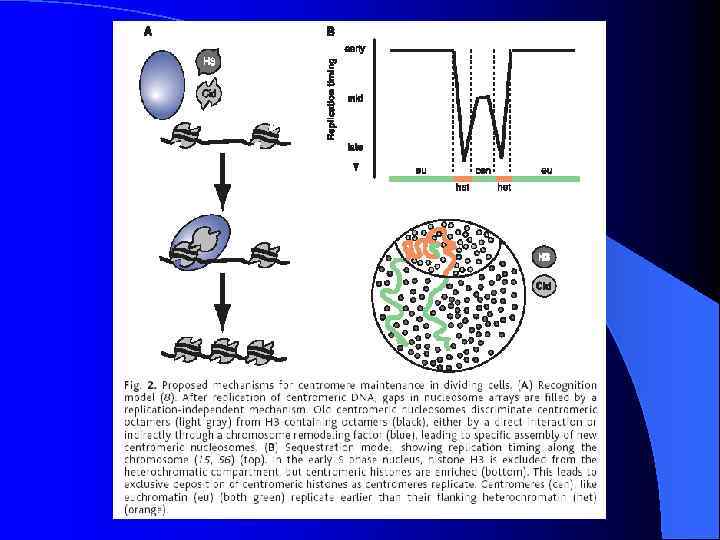
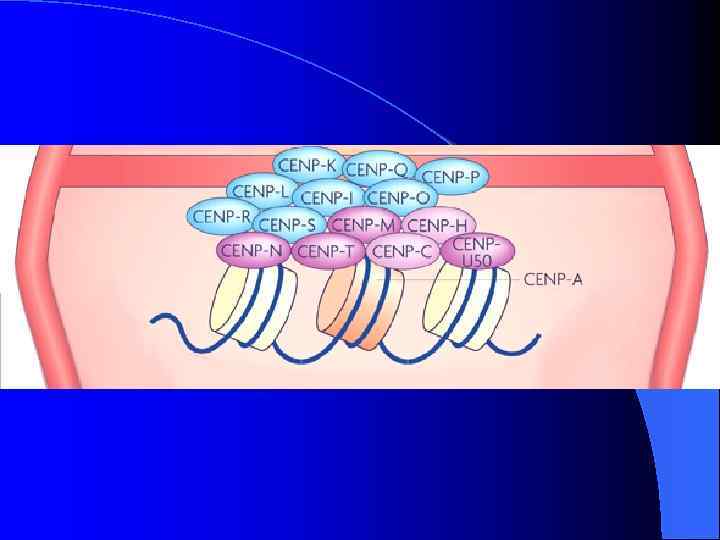
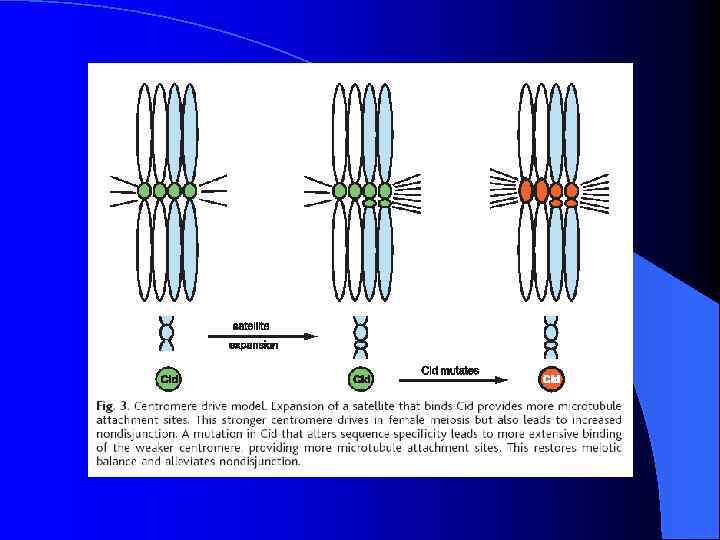
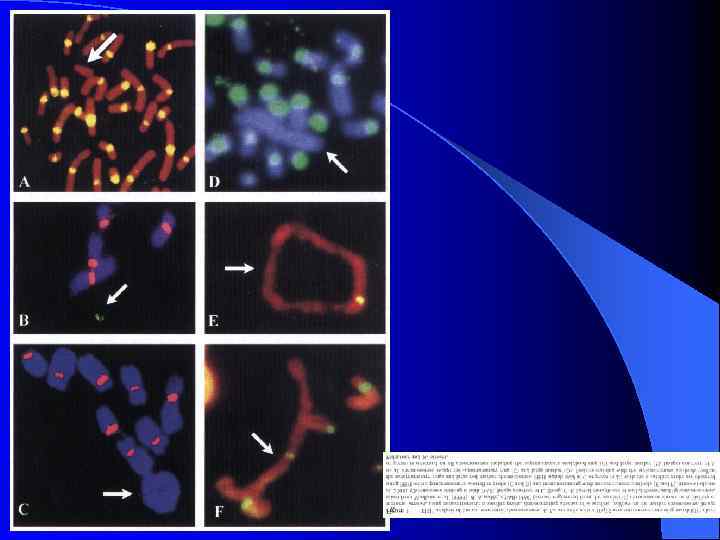
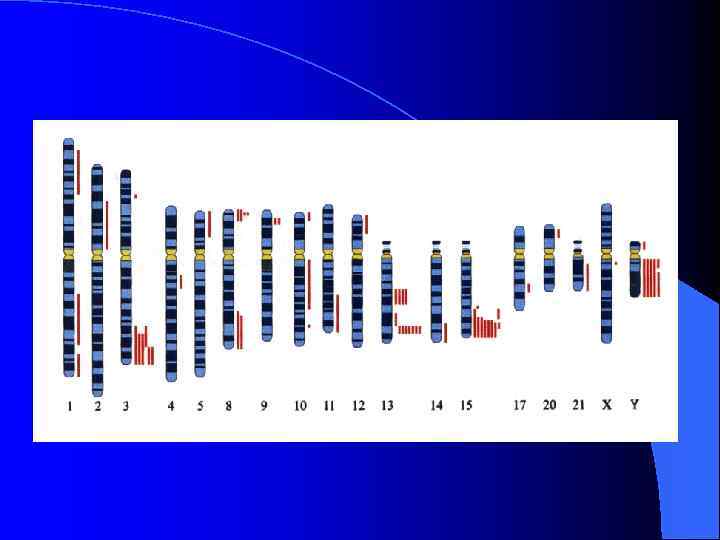
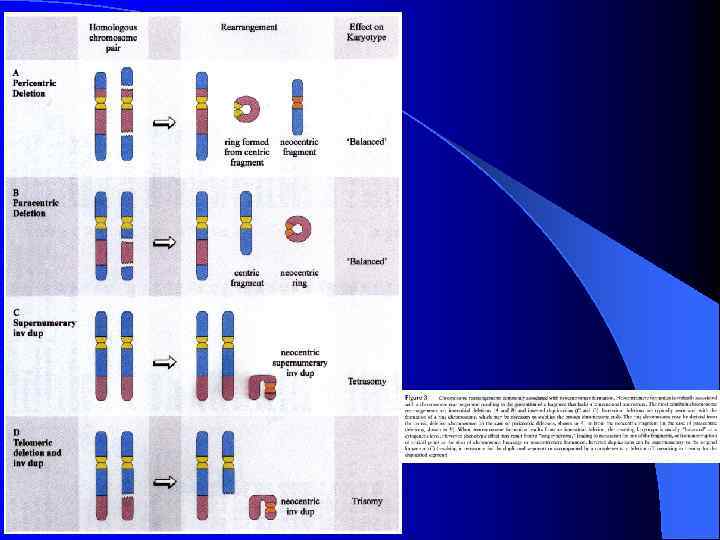
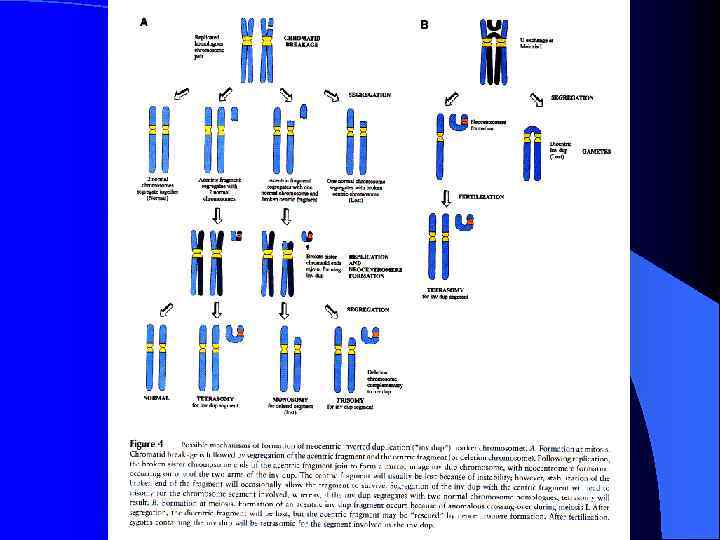
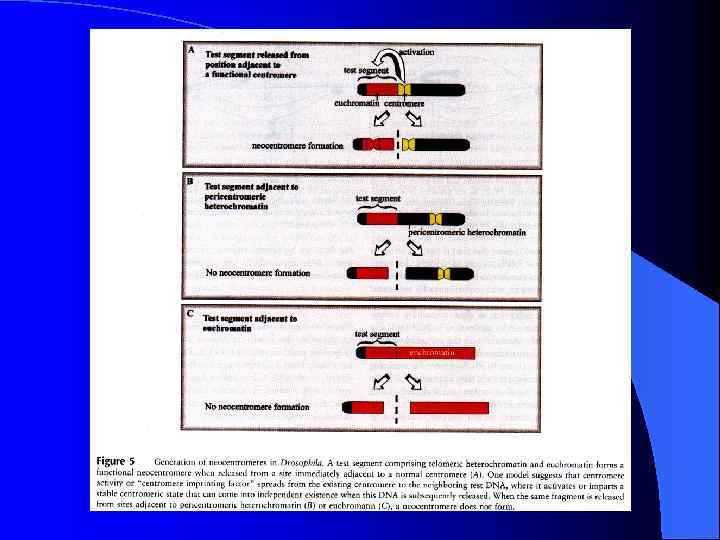
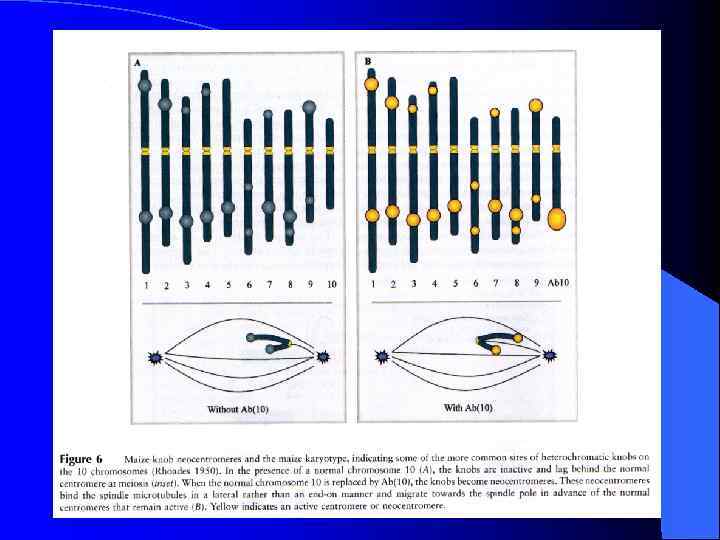

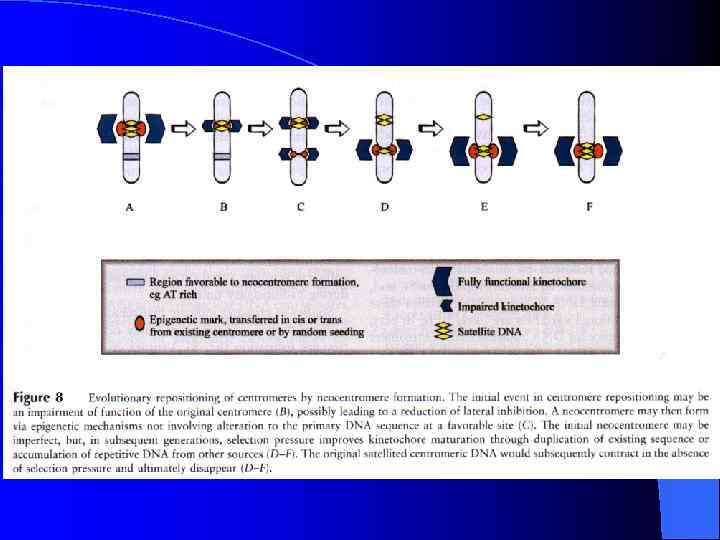

Неоцентромеры: • • • Возникновение неоцентромер – редкое событие Неоцентромеры могу возникать как митозе, так и в мейозе Неоцентромеры формируют функционально активный кинетохор и могут стабильно наследоваться в митозе и мейозе Формирование неоцентромер возможно в различных хромосомных сайтах, не связано с уникальной последовательностью, но сайты имеют некоторые общие характеристики (АТ обогащенность) Формирование неоцентромер обычно негативно сказывается на индивиде Происхождение неоцентромер человека, поведение в митозе и мейозе. Места локализации неоцентромер Состав протеинов в неоцентромерах Время репликации неоцентромерного района Maize Knobs или факультативные центромеры
Цитогенетика 16-18.ppt