

!2013_Lecture-06.ppt
- Количество слайдов: 135
 Что такое популяция? • Популяция – от лат. populus – народ, население. • Хотя популяционный подход в биологии идет от Ч. Дарвина, слово “популяция” не было термином до начала XX века. • Термин «популяция» введен датским генетиком В. Иогансеном в 1903 г. для обозначения неоднородной в генетическом отношении группы особей одного вида в отличие от однородной чистой линии. • • Термин «популяция» получил очень широкое распространение в биологии и при этом его понимание весьма различно. До сих пор возникают жаркие дискуссии о том, что же такое популяция, и по этому вопросу отсутствует единство мнений. Существуют принципиально различные подходы к определению понятия “популяция”. Причины этих разногласий: 1. Различия в объекте исследования. 2. Различия в научном подходе и целях исследования. Wilhelm Ludvig Johannsen (1857 - 1927)
Что такое популяция? • Популяция – от лат. populus – народ, население. • Хотя популяционный подход в биологии идет от Ч. Дарвина, слово “популяция” не было термином до начала XX века. • Термин «популяция» введен датским генетиком В. Иогансеном в 1903 г. для обозначения неоднородной в генетическом отношении группы особей одного вида в отличие от однородной чистой линии. • • Термин «популяция» получил очень широкое распространение в биологии и при этом его понимание весьма различно. До сих пор возникают жаркие дискуссии о том, что же такое популяция, и по этому вопросу отсутствует единство мнений. Существуют принципиально различные подходы к определению понятия “популяция”. Причины этих разногласий: 1. Различия в объекте исследования. 2. Различия в научном подходе и целях исследования. Wilhelm Ludvig Johannsen (1857 - 1927)
: “Популяция – это группа") Что такое популяция? 1. Максимально широкое определение Raymond Pearl (1937): “Популяция – это группа живых особей, выделяемая в некоторых рамках пространства и времени” • • Согласно этому определению границы популяции можно провести так, как нам это удобно в данном случае. Во многих ситуациях этот подход удобен.
Что такое популяция? 1. Максимально широкое определение Raymond Pearl (1937): “Популяция – это группа живых особей, выделяемая в некоторых рамках пространства и времени” • • Согласно этому определению границы популяции можно провести так, как нам это удобно в данном случае. Во многих ситуациях этот подход удобен.
 Что такое популяция? 2. Эволюционно-генетический подход “Популяция – это совокупность особей одного вида, занимающая определенную территорию, обладающая общим генофондом в результате свободного скрещивания (панмиксии) между особями внутри популяции и в определенной степени генетически изолированная от других популяций” • • Популяция – это “репродуктивное сообщество”. Идеальная, или “менделевская популяция” – обладает полной панмиктичностью и удовлетворяет всем требованиям закона Харди. Вайнберга. • Эволюционно-генетическое определение популяции неприемлемо для экологии по следующим причинам: – пригодно только для видов, размножающихся половым путем, в то время как экология изучает популяции всех организмов; – множество популяций, изучаемых экологами, не удовлетворяет критериям эволюционно-генетического подхода.
Что такое популяция? 2. Эволюционно-генетический подход “Популяция – это совокупность особей одного вида, занимающая определенную территорию, обладающая общим генофондом в результате свободного скрещивания (панмиксии) между особями внутри популяции и в определенной степени генетически изолированная от других популяций” • • Популяция – это “репродуктивное сообщество”. Идеальная, или “менделевская популяция” – обладает полной панмиктичностью и удовлетворяет всем требованиям закона Харди. Вайнберга. • Эволюционно-генетическое определение популяции неприемлемо для экологии по следующим причинам: – пригодно только для видов, размножающихся половым путем, в то время как экология изучает популяции всех организмов; – множество популяций, изучаемых экологами, не удовлетворяет критериям эволюционно-генетического подхода.
 Что такое популяция? 3. Биоценологический подход “Популяция – это совокупность особей одного вида, входящих в некоторый биоценоз” • В фитоценологии даже используют термин “ценопопуляция”. • Такое определение популяции непригодно для общей экологии по следующим причинам: – границы биоценозов далеко не всегда можно однозначно установить; – границы популяций животных далеко не всегда совпадают с границами биоценозов: • популяции мелких животных могут быть меньше биоценоза; • популяции крупных и подвижных животных всегда охватывают множество разных биоценозов; • на разных стадиях жизненного цикла животные могут быть приурочены к совершенно разным биотопам и биоценозам.
Что такое популяция? 3. Биоценологический подход “Популяция – это совокупность особей одного вида, входящих в некоторый биоценоз” • В фитоценологии даже используют термин “ценопопуляция”. • Такое определение популяции непригодно для общей экологии по следующим причинам: – границы биоценозов далеко не всегда можно однозначно установить; – границы популяций животных далеко не всегда совпадают с границами биоценозов: • популяции мелких животных могут быть меньше биоценоза; • популяции крупных и подвижных животных всегда охватывают множество разных биоценозов; • на разных стадиях жизненного цикла животные могут быть приурочены к совершенно разным биотопам и биоценозам.
 Что такое популяция? 4. “Жесткое” определение – характерно для некоторых, особенно отечественных, биологов-эволюционистов Например, Алексей Владимирович Яблоков (1987): Популяция – это “минимальная самовоспроизводящаяся группировка особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая самостоятельную генетическую систему и формирующая собственное экологическое гиперпространство”. . . “это достаточно многочисленная группа особей, на протяжении большого числа поколений в высокой степени изолированная от других групп. ” Такое определение мало пригодно для целей экологии по следующим причинам: – содержит трудно проверяемые или неопределяемые параметры (выделены красным); – применимо только к видам, размножающимся половым путем; – множество популяций, изучаемых экологами, явно не удовлетворяет критериям данного определения.
Что такое популяция? 4. “Жесткое” определение – характерно для некоторых, особенно отечественных, биологов-эволюционистов Например, Алексей Владимирович Яблоков (1987): Популяция – это “минимальная самовоспроизводящаяся группировка особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая самостоятельную генетическую систему и формирующая собственное экологическое гиперпространство”. . . “это достаточно многочисленная группа особей, на протяжении большого числа поколений в высокой степени изолированная от других групп. ” Такое определение мало пригодно для целей экологии по следующим причинам: – содержит трудно проверяемые или неопределяемые параметры (выделены красным); – применимо только к видам, размножающимся половым путем; – множество популяций, изучаемых экологами, явно не удовлетворяет критериям данного определения.
 Что такое популяция? 5. “Мягкое” определение – наиболее распространено в экологии Первый вариант: “Популяция – это любая способная к самовоспроизведению совокупность особей одного вида, населяющая некоторую территорию и относительно изолированная от других таких же совокупностей” Второй вариант (еще более “мягкое” определение): Без слов “способная к самовоспроизведению”, т. е. “любая, существующая в течение многих поколений, совокупность особей одного вида …” Преимущества "мягкого" определения популяции: – возможность включения всего разнообразия популяционных группировок; – пригодность для всех организмов независимо от формы размножения, размеров, подвижности и проч.
Что такое популяция? 5. “Мягкое” определение – наиболее распространено в экологии Первый вариант: “Популяция – это любая способная к самовоспроизведению совокупность особей одного вида, населяющая некоторую территорию и относительно изолированная от других таких же совокупностей” Второй вариант (еще более “мягкое” определение): Без слов “способная к самовоспроизведению”, т. е. “любая, существующая в течение многих поколений, совокупность особей одного вида …” Преимущества "мягкого" определения популяции: – возможность включения всего разнообразия популяционных группировок; – пригодность для всех организмов независимо от формы размножения, размеров, подвижности и проч.
 Разнообразие популяций • Причины разнообразия биологических свойств популяций – кроются в бесконечном разнообразии организмов – их размеров, структуры, физиологии, поведения, способов размножения, продолжительности жизни и т. д.
Разнообразие популяций • Причины разнообразия биологических свойств популяций – кроются в бесконечном разнообразии организмов – их размеров, структуры, физиологии, поведения, способов размножения, продолжительности жизни и т. д.
 популяции Ареал Ящерица") Разнообразие популяций Иерархия популяционных систем: – Географические популяции – Местные (локальные) популяции Ареал Ящерица Lacerta agilis Местная популяция
Разнообразие популяций Иерархия популяционных систем: – Географические популяции – Местные (локальные) популяции Ареал Ящерица Lacerta agilis Местная популяция
 популяции – Элементарные") Разнообразие популяций Иерархия популяционных систем: – Географические популяции – Местные (локальные) популяции – Элементарные популяции (демы) – Парцеллы, семейные группы и т. п. Демы Семейные группы Ящерица Lacerta agilis Местные популяции
Разнообразие популяций Иерархия популяционных систем: – Географические популяции – Местные (локальные) популяции – Элементарные популяции (демы) – Парцеллы, семейные группы и т. п. Демы Семейные группы Ящерица Lacerta agilis Местные популяции
 *: Гемипопуляции образуют отдельные стадии жизненного цикла") Разнообразие популяций Гемипопуляции В. Н. Беклемишев (1976) *: Гемипопуляции образуют отдельные стадии жизненного цикла видов, развивающихся с метаморфозом в тех случаях, когда они обитают в разных биотопах. Например: головастики и взрослые лягушки, личинки стрекоз и взрослые стрекозы и т. п. Беклемишев Владимир Николаевич (1890– 1962) * Биогеоценологические основы сравнительной паразитологии. М. : Наука. 1976. 502 с.
Разнообразие популяций Гемипопуляции В. Н. Беклемишев (1976) *: Гемипопуляции образуют отдельные стадии жизненного цикла видов, развивающихся с метаморфозом в тех случаях, когда они обитают в разных биотопах. Например: головастики и взрослые лягушки, личинки стрекоз и взрослые стрекозы и т. п. Беклемишев Владимир Николаевич (1890– 1962) * Биогеоценологические основы сравнительной паразитологии. М. : Наука. 1976. 502 с.
 Разнообразие популяций Временные и постоянные популяции – Временные – спорадически или сезонно заселяемые территории, население на которых существует недолго (одно или несколько поколений) и в конце концов вымирает. Сезонные популяции – У перелетных птиц, кочующих млекопитающих и других мигрирующих организмов. Независимые и зависимые популяции – Независимые – самовоспроизводятся (r > 0), служат источником расселяющихся особей. – Зависимые – неспособны к самоподдержанию (r < 0), существуют благодаря притоку расселяющихся особей извне.
Разнообразие популяций Временные и постоянные популяции – Временные – спорадически или сезонно заселяемые территории, население на которых существует недолго (одно или несколько поколений) и в конце концов вымирает. Сезонные популяции – У перелетных птиц, кочующих млекопитающих и других мигрирующих организмов. Независимые и зависимые популяции – Независимые – самовоспроизводятся (r > 0), служат источником расселяющихся особей. – Зависимые – неспособны к самоподдержанию (r < 0), существуют благодаря притоку расселяющихся особей извне.
 Основные особенности популяционного подхода Различия в степени интегрированности популяций Две крайние точки зрения: 1. Популяция – в высшей степени интегрированная и целостная система, реагирующая на изменения окружающей среды подобно настоящему организму. Стратегическая задача популяции – поддерживать свое стабильное существование, для чего в ней есть развитые механизмы саморегуляции. Популяция – это просто совокупность особей и как целое вообще не представляет никакой реальности. “Поведение популяции” – это просто метафора, удобный нам способ описания. 2. Как разрешить это противоречие? В чем его причины? – – в различиях методологических подходов и типов мышления; в различиях между объектами исследования разных ученых. Решение противоречия – в компромиссе: – – популяции некоторых организмов являются достаточно интегрированными и саморегулируемыми системами; в популяциях других видов интеграция и саморегуляция развиты в меньшей степени, или даже вообще отсутствуют.
Основные особенности популяционного подхода Различия в степени интегрированности популяций Две крайние точки зрения: 1. Популяция – в высшей степени интегрированная и целостная система, реагирующая на изменения окружающей среды подобно настоящему организму. Стратегическая задача популяции – поддерживать свое стабильное существование, для чего в ней есть развитые механизмы саморегуляции. Популяция – это просто совокупность особей и как целое вообще не представляет никакой реальности. “Поведение популяции” – это просто метафора, удобный нам способ описания. 2. Как разрешить это противоречие? В чем его причины? – – в различиях методологических подходов и типов мышления; в различиях между объектами исследования разных ученых. Решение противоречия – в компромиссе: – – популяции некоторых организмов являются достаточно интегрированными и саморегулируемыми системами; в популяциях других видов интеграция и саморегуляция развиты в меньшей степени, или даже вообще отсутствуют.
 Основные особенности популяционного подхода Значение пространственно-временных масштабов • • • Каждая популяция существует в специфическом пространстве и времени. Даже понятие экосистемы будет очень разным «с точки зрения» различных организмов. Эти различия в основном определяются размерами организма, скоростью его размножения и плодовитостью. Примеры – обитатели леса Средней России: 1. Липовая тля
Основные особенности популяционного подхода Значение пространственно-временных масштабов • • • Каждая популяция существует в специфическом пространстве и времени. Даже понятие экосистемы будет очень разным «с точки зрения» различных организмов. Эти различия в основном определяются размерами организма, скоростью его размножения и плодовитостью. Примеры – обитатели леса Средней России: 1. Липовая тля
 Основные особенности популяционного подхода Значение пространственно-временных масштабов Примеры – обитатели леса Средней России: 1. Липовая тля 2. Рыжая лесная полевка
Основные особенности популяционного подхода Значение пространственно-временных масштабов Примеры – обитатели леса Средней России: 1. Липовая тля 2. Рыжая лесная полевка
 Основные особенности популяционного подхода Значение пространственно-временных масштабов Примеры – обитатели леса Средней России: 1. Липовая тля 2. Рыжая лесная полевка 3. Лось
Основные особенности популяционного подхода Значение пространственно-временных масштабов Примеры – обитатели леса Средней России: 1. Липовая тля 2. Рыжая лесная полевка 3. Лось
 Основные особенности популяционного подхода Значение пространственно-временных масштабов Примеры – обитатели леса Средней России: 1. Липовая тля 2. Рыжая лесная полевка 3. Лось Итак: различия пространственно-временных масштабов между разными организмами могут быть огромными, и поэтому динамика популяций этих организмов и их экосистемные отношения очень различны.
Основные особенности популяционного подхода Значение пространственно-временных масштабов Примеры – обитатели леса Средней России: 1. Липовая тля 2. Рыжая лесная полевка 3. Лось Итак: различия пространственно-временных масштабов между разными организмами могут быть огромными, и поэтому динамика популяций этих организмов и их экосистемные отношения очень различны.
 Основные особенности популяционного подхода Популяции унитарных и модулярных организмов • • • 1. 2. Из чего состоят популяции? Из особей. Но что такое особь? Длительное время никто не задавал этого вопроса, поскольку экология популяций занималась преимущественно животными. Растения же существенно отличаются от животных биологической природой особей. Существует два фундаментально различных типа организмов: Унитарные Модулярные В прошлом многие экологические и эволюционные обобщения были сделаны так, будто в мире существуют только унитарные организмы. И это привело ко многим заблуждениям.
Основные особенности популяционного подхода Популяции унитарных и модулярных организмов • • • 1. 2. Из чего состоят популяции? Из особей. Но что такое особь? Длительное время никто не задавал этого вопроса, поскольку экология популяций занималась преимущественно животными. Растения же существенно отличаются от животных биологической природой особей. Существует два фундаментально различных типа организмов: Унитарные Модулярные В прошлом многие экологические и эволюционные обобщения были сделаны так, будто в мире существуют только унитарные организмы. И это привело ко многим заблуждениям.
 Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Унитарные организмы • • Большинство животных, часть грибов и одноклеточных Имеют типичный онтогенез: – • • • зигота – эмбрион – организм (особь) Строение и размеры особи более или менее стандартны и предопределены генетически. Особи имеют ограниченное время жизни и всегда умирают. Поэтому возрастная структура популяций достаточно проста.
Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Унитарные организмы • • Большинство животных, часть грибов и одноклеточных Имеют типичный онтогенез: – • • • зигота – эмбрион – организм (особь) Строение и размеры особи более или менее стандартны и предопределены генетически. Особи имеют ограниченное время жизни и всегда умирают. Поэтому возрастная структура популяций достаточно проста.
 Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы • Большинство растений, многие грибы и некоторые животные: – • (гидроидные и коралловые полипы, мшанки, губки, колониальные асцидии) Их онтогенез: – зигота – модуль – новые модули (вегетативное размножение) – модулярный организм
Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы • Большинство растений, многие грибы и некоторые животные: – • (гидроидные и коралловые полипы, мшанки, губки, колониальные асцидии) Их онтогенез: – зигота – модуль – новые модули (вегетативное размножение) – модулярный организм
 Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы • Большинство растений, многие грибы и некоторые животные: – • (гидроидные и коралловые полипы, мшанки, губки, колониальные асцидии) Их онтогенез: – зигота – модуль – новые модули (вегетативное размножение) – модулярный организм
Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы • Большинство растений, многие грибы и некоторые животные: – • (гидроидные и коралловые полипы, мшанки, губки, колониальные асцидии) Их онтогенез: – зигота – модуль – новые модули (вегетативное размножение) – модулярный организм
 Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы • Большинство растений, многие грибы и некоторые животные: – • Их онтогенез: – • • • (гидроидные и коралловые полипы, мшанки, губки, колониальные асцидии) зигота – модуль – новые модули (вегетативное размножение) – модулярный организм Развитие очень пластично и не столь предопределено генетически. Строение, число входящих в организм модулей и его размеры очень сильно варьируют в зависимости от среды обитания. Тело организма имеет «возрастную структуру» - молодые и старые модули. Потенциально бессмертны. Возрастная структура популяций очень сложна.
Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы • Большинство растений, многие грибы и некоторые животные: – • Их онтогенез: – • • • (гидроидные и коралловые полипы, мшанки, губки, колониальные асцидии) зигота – модуль – новые модули (вегетативное размножение) – модулярный организм Развитие очень пластично и не столь предопределено генетически. Строение, число входящих в организм модулей и его размеры очень сильно варьируют в зависимости от среды обитания. Тело организма имеет «возрастную структуру» - молодые и старые модули. Потенциально бессмертны. Возрастная структура популяций очень сложна.
 Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы Популяции модулярных организмов можно изучать на 3 уровнях: 1. На уровне модулей Например: отдельные листья дерева, побеги злаков 2. На уровне групп модулей (организмов) Например: отдельные деревья, кусты злаков, куртинки и т. п. 3. На уровне клонов и генетов Клон – результат вегетативного размножения модуля. Генет – “генетический индивидуум” – все, что образовалось из одной зиготы.
Основные особенности популяционного подхода Популяции унитарных и модулярных организмов Модулярные организмы Популяции модулярных организмов можно изучать на 3 уровнях: 1. На уровне модулей Например: отдельные листья дерева, побеги злаков 2. На уровне групп модулей (организмов) Например: отдельные деревья, кусты злаков, куртинки и т. п. 3. На уровне клонов и генетов Клон – результат вегетативного размножения модуля. Генет – “генетический индивидуум” – все, что образовалось из одной зиготы.
 Методы оценки численности популяций • Прямой подсчет – Пригоден для крупных растений, сидячих или малоподвижных животных. – Учеты крупных животных с самолета или вертолета. • Метод повторного отлова A – отловлено, помечено и выпущено B – снова отловлено, из них оказалось C помеченных В этом случае: • Метод требует соблюдения множества условий и имеет поэтому ограниченное применение.
Методы оценки численности популяций • Прямой подсчет – Пригоден для крупных растений, сидячих или малоподвижных животных. – Учеты крупных животных с самолета или вертолета. • Метод повторного отлова A – отловлено, помечено и выпущено B – снова отловлено, из них оказалось C помеченных В этом случае: • Метод требует соблюдения множества условий и имеет поэтому ограниченное применение.
 Методы оценки численности популяций • Выборочные методы – – – Рандомизированное взятие из популяции количественных выборок. Каждая выборка берется с определенной площади или объема. Подсчет количества особей в каждой выборке. Вычисление средней плотности популяции и показателей ее варьирования. Это основной и важнейший метод.
Методы оценки численности популяций • Выборочные методы – – – Рандомизированное взятие из популяции количественных выборок. Каждая выборка берется с определенной площади или объема. Подсчет количества особей в каждой выборке. Вычисление средней плотности популяции и показателей ее варьирования. Это основной и важнейший метод.
 Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – Число мышей, попавших в ловушки за сутки.
Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – Число мышей, попавших в ловушки за сутки.
 Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – Число мышей, попавших в ловушки за сутки. – Число насекомых, прилетевших на светоловушку за ночь.
Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – Число мышей, попавших в ловушки за сутки. – Число насекомых, прилетевших на светоловушку за ночь.
 Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – Число мышей, попавших в ловушки за сутки. – Число насекомых, прилетевших на светоловушку за ночь. – Число насекомых, попавших в ловушку того или иного типа. Феромонная ловушка Почвенная ловушка (pit-fall trap) Ловушка для короедов
Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – Число мышей, попавших в ловушки за сутки. – Число насекомых, прилетевших на светоловушку за ночь. – Число насекомых, попавших в ловушку того или иного типа. Феромонная ловушка Почвенная ловушка (pit-fall trap) Ловушка для короедов
 Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – – – Число мышей, попавших в ловушки за сутки. Число насекомых, прилетевших на светоловушку за ночь. Число мух, привлеченных пищевой приманкой за час. Число животных, добытых в среднем одним охотником за сезон охоты. И т. д и т. п. Ловушка для омаров
Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – – – Число мышей, попавших в ловушки за сутки. Число насекомых, прилетевших на светоловушку за ночь. Число мух, привлеченных пищевой приманкой за час. Число животных, добытых в среднем одним охотником за сезон охоты. И т. д и т. п. Ловушка для омаров
 Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – – – Число мышей, попавших в ловушки за сутки. Число насекомых, прилетевших на светоловушку за ночь. Число мух, привлеченных пищевой приманкой за час. Число животных, добытых в среднем одним охотником за сезон охоты. И т. д и т. п.
Методы оценки численности популяций • Выборочные методы • Использование относительных показателей численности – – – Число мышей, попавших в ловушки за сутки. Число насекомых, прилетевших на светоловушку за ночь. Число мух, привлеченных пищевой приманкой за час. Число животных, добытых в среднем одним охотником за сезон охоты. И т. д и т. п.
 Пространственная структура популяций Три типа распределения особей в пространстве 1. Случайное Число проб (рассеянное, диффузное) Число особей в пробе – (число особей в пробе или расстояние до ближайшего соседа) – Дисперсия Случайное
Пространственная структура популяций Три типа распределения особей в пространстве 1. Случайное Число проб (рассеянное, диффузное) Число особей в пробе – (число особей в пробе или расстояние до ближайшего соседа) – Дисперсия Случайное
 Пространственная структура популяций Три типа распределения особей в пространстве 1. Случайное 2. Равномерное Число проб (регулярное, единообразное, сверхрассеянное) Число особей в пробе – (число особей в пробе или расстояние до ближайшего соседа) – Дисперсия Равномерное
Пространственная структура популяций Три типа распределения особей в пространстве 1. Случайное 2. Равномерное Число проб (регулярное, единообразное, сверхрассеянное) Число особей в пробе – (число особей в пробе или расстояние до ближайшего соседа) – Дисперсия Равномерное
 Пространственная структура популяций Три типа распределения особей в пространстве 1. Случайное 2. Равномерное 3. Агрегированное Число проб (скученное, конгрегированное) Число особей в пробе – (число особей в пробе или расстояние до ближайшего соседа) – Дисперсия Агрегированное
Пространственная структура популяций Три типа распределения особей в пространстве 1. Случайное 2. Равномерное 3. Агрегированное Число проб (скученное, конгрегированное) Число особей в пробе – (число особей в пробе или расстояние до ближайшего соседа) – Дисперсия Агрегированное
 Пространственная структура популяций Три типа распределения особей в пространстве Равномерное Случайное Агрегированное
Пространственная структура популяций Три типа распределения особей в пространстве Равномерное Случайное Агрегированное
 (распределение особей") Пространственная структура популяций Основные типы пространственной структуры популяций – Диффузные популяции (сплошные) (распределение особей внутри популяции может быть любым) Случайное Равномерное Агрегированное
Пространственная структура популяций Основные типы пространственной структуры популяций – Диффузные популяции (сплошные) (распределение особей внутри популяции может быть любым) Случайное Равномерное Агрегированное
 – Частично") Пространственная структура популяций Основные типы пространственной структуры популяций – Диффузные популяции (сплошные) – Частично дискретные популяции • границы между популяциями нередко достаточно условны
Пространственная структура популяций Основные типы пространственной структуры популяций – Диффузные популяции (сплошные) – Частично дискретные популяции • границы между популяциями нередко достаточно условны
 – Частично") Пространственная структура популяций Основные типы пространственной структуры популяций – Диффузные популяции (сплошные) – Частично дискретные популяции – Полностью дискретные популяции островного типа • границы между популяциями отчетливы; • как правило определяются тем, что популяции разделены территориями, непригодными для обитания данного вида.
Пространственная структура популяций Основные типы пространственной структуры популяций – Диффузные популяции (сплошные) – Частично дискретные популяции – Полностью дискретные популяции островного типа • границы между популяциями отчетливы; • как правило определяются тем, что популяции разделены территориями, непригодными для обитания данного вида.
 Частично") Пространственная структура популяций Основные типы пространственной структуры популяций – – Диффузные популяции (сплошные) Частично дискретные популяции Полностью дискретные популяции островного типа Популяции специальных типов: • ленточные • сетчатые • четковидные и т. д.
Пространственная структура популяций Основные типы пространственной структуры популяций – – Диффузные популяции (сплошные) Частично дискретные популяции Полностью дискретные популяции островного типа Популяции специальных типов: • ленточные • сетчатые • четковидные и т. д.
 Частично") Пространственная структура популяций Основные типы пространственной структуры популяций – – Диффузные популяции (сплошные) Частично дискретные популяции Полностью дискретные популяции островного типа Популяции специальных типов: • ленточные • сетчатые • четковидные и т. д. • Нередко в пределах ареала вида встречаются популяции различных типов
Пространственная структура популяций Основные типы пространственной структуры популяций – – Диффузные популяции (сплошные) Частично дискретные популяции Полностью дискретные популяции островного типа Популяции специальных типов: • ленточные • сетчатые • четковидные и т. д. • Нередко в пределах ареала вида встречаются популяции различных типов
 Пространственная структура популяций Примеры разных типов популяций в ареале вида: сплошные и островные
Пространственная структура популяций Примеры разных типов популяций в ареале вида: сплошные и островные
 Пространственная структура популяций Примеры разных типов популяций в ареале вида: ленточные и островные
Пространственная структура популяций Примеры разных типов популяций в ареале вида: ленточные и островные
 Динамика пространственной структуры популяций во времени
Динамика пространственной структуры популяций во времени
 Динамика пространственной структуры популяций во времени
Динамика пространственной структуры популяций во времени
 2. Динамика популяции, обладающей возрастной") Динамика популяций 1. Неограниченный рост популяции (независимый от плотности) 2. Динамика популяции, обладающей возрастной структурой 3. Ограниченный рост популяции (зависимый от плотности)
Динамика популяций 1. Неограниченный рост популяции (независимый от плотности) 2. Динамика популяции, обладающей возрастной структурой 3. Ограниченный рост популяции (зависимый от плотности)
 • “Отец демографии”. • Впервые исследовал динамику населения") Неограниченный рост популяции John Graunt (1662) • “Отец демографии”. • Впервые исследовал динамику населения Лондона. • Впервые указал на необходимость точного количественного измерения рождаемости, смертности, соотношения полов и возрастной структуры популяций человека. • Впервые высказал идею о возможности неограниченного роста популяций. • Впервые дал оценку потенциальной скорости роста популяций: – Население Лондона может удвоиться за 64 года. John Graunt 1620 -1674
Неограниченный рост популяции John Graunt (1662) • “Отец демографии”. • Впервые исследовал динамику населения Лондона. • Впервые указал на необходимость точного количественного измерения рождаемости, смертности, соотношения полов и возрастной структуры популяций человека. • Впервые высказал идею о возможности неограниченного роста популяций. • Впервые дал оценку потенциальной скорости роста популяций: – Население Лондона может удвоиться за 64 года. John Graunt 1620 -1674
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Когорта – совокупность особей, родившихся одновременно (в пределах необходимой точности измерений) – • Популяция с дискретными поколениями практически всегда (за исключением периода размножения) представлена одной когортой. Здесь продолжительность поколения примерно равна длительности существования когорты. Число особей • например, в течение 1 часа, 1 суток, 1 недели, 1 месяца, 1 года. Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Когорта – совокупность особей, родившихся одновременно (в пределах необходимой точности измерений) – • Популяция с дискретными поколениями практически всегда (за исключением периода размножения) представлена одной когортой. Здесь продолжительность поколения примерно равна длительности существования когорты. Число особей • например, в течение 1 часа, 1 суток, 1 недели, 1 месяца, 1 года. Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Динамику численности такой популяции можно изображать двумя способами: – – Второй способ используют чаще Число особей • По точкам максимальной численности каждой когорты (поколения) в момент появления ее на свет По точкам минимальной численности каждой когорты (поколения) в начале периода размножения Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Динамику численности такой популяции можно изображать двумя способами: – – Второй способ используют чаще Число особей • По точкам максимальной численности каждой когорты (поколения) в момент появления ее на свет По точкам минимальной численности каждой когорты (поколения) в начале периода размножения Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Рост подобной популяции можно описать уравнением: – выживаемость особей в поколении 0 до момента их размножения Число особей – средняя плодовитость особей в поколении 0 Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Рост подобной популяции можно описать уравнением: – выживаемость особей в поколении 0 до момента их размножения Число особей – средняя плодовитость особей в поколении 0 Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Или же вторым способом: Число особей – выживаемость особей в поколении 1 до момента их размножения Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Или же вторым способом: Число особей – выживаемость особей в поколении 1 до момента их размножения Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями Если условия среды стабильны, то s 1 = s 2 = s И тогда эти два уравнения выглядят одинаково: • Поэтому мы для упрощения будем использовать первое уравнение Число особей • • Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями Если условия среды стабильны, то s 1 = s 2 = s И тогда эти два уравнения выглядят одинаково: • Поэтому мы для упрощения будем использовать первое уравнение Число особей • • Когорта 2 Когорта 0 Поколение 0 Когорта 1 Поколение 2 Время
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • • Однако, если условия стабильны, то s 1 = s 2 = s И тогда оба уравнения выглядят одинаково: • • Поэтому мы для упрощения будем использовать первое уравнение Для раздельнополой популяции это уравнение должно быть видоизменено так: – средняя доля самок в популяции – средняя плодовитость одной самки • Коэффициент размножения популяции равен: • • И тогда: или: Это уравнение описывает так называемый “геометрический рост популяции” Поэтому – это также и “коэффициент геометрического роста популяции” •
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • • Однако, если условия стабильны, то s 1 = s 2 = s И тогда оба уравнения выглядят одинаково: • • Поэтому мы для упрощения будем использовать первое уравнение Для раздельнополой популяции это уравнение должно быть видоизменено так: – средняя доля самок в популяции – средняя плодовитость одной самки • Коэффициент размножения популяции равен: • • И тогда: или: Это уравнение описывает так называемый “геометрический рост популяции” Поэтому – это также и “коэффициент геометрического роста популяции” •
 Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Популяция растет, если: • Численность популяции не изменяется, если: • Популяция сокращает численность, если:
Неограниченный рост популяции Геометрический рост популяции с дискретными поколениями • Популяция растет, если: • Численность популяции не изменяется, если: • Популяция сокращает численность, если:
 Неограниченный рост популяции Геометрический рост популяции с перекрывающимися поколениями и дискретным размножением • • У видов с сезонной репродукцией, особи которых размножаются многократно, популяции состоят из нескольких последовательно родившихся когорт и растут дискретно, увеличиваясь только в периоды размножения. Дискретный рост подобных популяций лучше всего описывает следующее конечное разностное уравнение: Nt – величина популяции в момент t p – вероятность выживания особи за 1 интервал b – рождаемость на 1 самку на 1 интервал Преобразуем уравнение так: f – средняя доля самок в популяции • Тогда коэффициент геометрического роста будет равен: • В результате: • или иначе:
Неограниченный рост популяции Геометрический рост популяции с перекрывающимися поколениями и дискретным размножением • • У видов с сезонной репродукцией, особи которых размножаются многократно, популяции состоят из нескольких последовательно родившихся когорт и растут дискретно, увеличиваясь только в периоды размножения. Дискретный рост подобных популяций лучше всего описывает следующее конечное разностное уравнение: Nt – величина популяции в момент t p – вероятность выживания особи за 1 интервал b – рождаемость на 1 самку на 1 интервал Преобразуем уравнение так: f – средняя доля самок в популяции • Тогда коэффициент геометрического роста будет равен: • В результате: • или иначе:
 Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • У видов с непрерывным размножением мы не можем измерять время поколениями и коэффициент геометрического роста для них непригоден. Здесь необходим другой подход. • Скорость роста популяции равна: • Или в дифференциальном виде: • Удельная мгновенная скорость роста популяции составляет: • r – иначе называется “мальтузианским параметром” – – Имеет размерность 1/время Если r > 0 – популяция растет Если r = 0 – популяция не изменяет свою численность Если r < 0 – популяция сокращается
Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • У видов с непрерывным размножением мы не можем измерять время поколениями и коэффициент геометрического роста для них непригоден. Здесь необходим другой подход. • Скорость роста популяции равна: • Или в дифференциальном виде: • Удельная мгновенная скорость роста популяции составляет: • r – иначе называется “мальтузианским параметром” – – Имеет размерность 1/время Если r > 0 – популяция растет Если r = 0 – популяция не изменяет свою численность Если r < 0 – популяция сокращается
 Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • В полностью замкнутой популяции: – – • b – удельная мгновенная рождаемость d – удельная мгновенная смертность В незамкнутой популяции: – – i – удельная мгновенная иммиграция e – удельная мгновенная эмиграция
Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • В полностью замкнутой популяции: – – • b – удельная мгновенная рождаемость d – удельная мгновенная смертность В незамкнутой популяции: – – i – удельная мгновенная иммиграция e – удельная мгновенная эмиграция
 Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • Если проинтегрировать дифференциальное уравнение • то получится: • Это так называемое уравнение экспоненциального роста популяции, или уравнение Лотки Это уравнение впервые получено итальянским математиком Альфредом Лоткой в 1925 г. Alfred Lotka
Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • Если проинтегрировать дифференциальное уравнение • то получится: • Это так называемое уравнение экспоненциального роста популяции, или уравнение Лотки Это уравнение впервые получено итальянским математиком Альфредом Лоткой в 1925 г. Alfred Lotka
 Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • Графики уравнения Начальная численность N = 10 r = 0, 0. 06, 0. 08, 0. 14, 0. 22 Alfred Lotka
Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • Графики уравнения Начальная численность N = 10 r = 0, 0. 06, 0. 08, 0. 14, 0. 22 Alfred Lotka
 Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • После логарифмирования экспоненциального уравнения получаем: • На графике: Начальная численность N = 10 r = 0, 0. 06, 0. 08, 0. 14, 0. 22 Alfred Lotka
Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением • После логарифмирования экспоненциального уравнения получаем: • На графике: Начальная численность N = 10 r = 0, 0. 06, 0. 08, 0. 14, 0. 22 Alfred Lotka
 Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением Соотношения между параметрами λ и r • Уравнение геометрического роста: • Уравнение экспоненциального роста: • Это означает, что: • Отсюда: • Или же: Alfred Lotka
Неограниченный рост популяции Экспоненциальный рост популяции с непрерывным размножением Соотношения между параметрами λ и r • Уравнение геометрического роста: • Уравнение экспоненциального роста: • Это означает, что: • Отсюда: • Или же: Alfred Lotka
 Неограниченный рост популяции Возможности неограниченного роста популяций в природе Фазаны на о. Протекшн Кольчатая горлица на Британских островах
Неограниченный рост популяции Возможности неограниченного роста популяций в природе Фазаны на о. Протекшн Кольчатая горлица на Британских островах
 Неограниченный рост популяции Возможности неограниченного роста популяций в природе Овцы на о. Тасмания
Неограниченный рост популяции Возможности неограниченного роста популяций в природе Овцы на о. Тасмания
 Неограниченный рост популяции Возможности неограниченного роста популяций в природе Северные олени на о. Сент-Пол
Неограниченный рост популяции Возможности неограниченного роста популяций в природе Северные олени на о. Сент-Пол
 • Биотический потенциал —") Неограниченный рост популяции Биотический потенциал вида • R. Chapman (1928) • Биотический потенциал — характеристика внутренней потенциальной способности данной популяции к росту при стабильном возрастном составе и оптимальных условиях среды; выражается величиной прироста популяции за единицу времени в расчете на одну особь.
Неограниченный рост популяции Биотический потенциал вида • R. Chapman (1928) • Биотический потенциал — характеристика внутренней потенциальной способности данной популяции к росту при стабильном возрастном составе и оптимальных условиях среды; выражается величиной прироста популяции за единицу времени в расчете на одну особь.
") Неограниченный рост популяции Биотический потенциал вида • R. Chapman (1928)
Неограниченный рост популяции Биотический потенциал вида • R. Chapman (1928)
 Максимальное значение мальтузианского") Неограниченный рост популяции Биотический потенциал вида • • R. Chapman (1928) Максимальное значение мальтузианского параметра, характеризующее способность вида к размножению в наиболее благоприятных условиях: Вид Длит. поколения (сутки) 0. 014 rmax (сутки-1) 60 0. 3 -0. 5 1. 24 80 0. 12 150 0. 015 Собака 1000 0. 009 Человек 7000 0. 0003 Кишечная палочка Инфузория-туфелька Мучной хрущак Серая крыса
Неограниченный рост популяции Биотический потенциал вида • • R. Chapman (1928) Максимальное значение мальтузианского параметра, характеризующее способность вида к размножению в наиболее благоприятных условиях: Вид Длит. поколения (сутки) 0. 014 rmax (сутки-1) 60 0. 3 -0. 5 1. 24 80 0. 12 150 0. 015 Собака 1000 0. 009 Человек 7000 0. 0003 Кишечная палочка Инфузория-туфелька Мучной хрущак Серая крыса
 Неограниченный рост популяции Связь максимально возможной скорости роста популяции с продолжительностью генерации и величиной организма:
Неограниченный рост популяции Связь максимально возможной скорости роста популяции с продолжительностью генерации и величиной организма:
 Динамика популяции, обладающей возрастной структурой • • • Экспоненциальная модель уравнивает особей по выживаемости и плодовитости. Однако, особи в популяции очень различаются по этим параметрам, особенно в зависимости от их возраста. Поэтому необходимо учитывать возрастную структуру популяции.
Динамика популяции, обладающей возрастной структурой • • • Экспоненциальная модель уравнивает особей по выживаемости и плодовитости. Однако, особи в популяции очень различаются по этим параметрам, особенно в зависимости от их возраста. Поэтому необходимо учитывать возрастную структуру популяции.
 Динамика популяции, обладающей возрастной структурой
Динамика популяции, обладающей возрастной структурой
 Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – Календарного возраста особей Сосна Человек
Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – Календарного возраста особей Сосна Человек
 Динамика популяции, обладающей возрастной структурой Возрастная структура популяции человека
Динамика популяции, обладающей возрастной структурой Возрастная структура популяции человека
 Динамика популяции, обладающей возрастной структурой Возрастная структура популяции сельди
Динамика популяции, обладающей возрастной структурой Возрастная структура популяции сельди
 Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – – Календарного возраста особей Стадии жизненного цикла: Растения: семена – (p) проростки – (j) ювенильные – (im) имматурные – (v) виргинные (вегетативные) – (g) генеративные (молодые, средневозрастные, старые) – (ss) субсенильные – (s) сенильные. Периоды жизни растения - возрастные состояния растения на протяжении его онтогенеза. По Т. А. Работнову выделяют следующие возрастные состояния: - латентный период - период существования особи в состоянии семени; - период проростка - период появления всходов; - ювенильный период - период несформированности признаков и свойств, свойственных взрослой особи; - имматурный период - период усиленного образования вегетативных органов; - виргинильный период - период, когда растение готово к воспроизводству потомства, но еще не размножается половым путем; - репродуктивный период - период, когда растение цветет, плодоносит и дает семена; - сенильный период - период ослабления цветения, приостановления вегетативного роста и вегетативного размножения.
Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – – Календарного возраста особей Стадии жизненного цикла: Растения: семена – (p) проростки – (j) ювенильные – (im) имматурные – (v) виргинные (вегетативные) – (g) генеративные (молодые, средневозрастные, старые) – (ss) субсенильные – (s) сенильные. Периоды жизни растения - возрастные состояния растения на протяжении его онтогенеза. По Т. А. Работнову выделяют следующие возрастные состояния: - латентный период - период существования особи в состоянии семени; - период проростка - период появления всходов; - ювенильный период - период несформированности признаков и свойств, свойственных взрослой особи; - имматурный период - период усиленного образования вегетативных органов; - виргинильный период - период, когда растение готово к воспроизводству потомства, но еще не размножается половым путем; - репродуктивный период - период, когда растение цветет, плодоносит и дает семена; - сенильный период - период ослабления цветения, приостановления вегетативного роста и вегетативного размножения.
 Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – – Календарного возраста особей Стадии жизненного цикла: Растения: семена – (p) проростки – (j) ювенильные – (im) имматурные – (v) виргинные (вегетативные) – (g) генеративные (молодые, средневозрастные, старые) – (ss) субсенильные – (s) сенильные. Демографическая структура популяций ели, березы и осины на пробной площади 0, 04 га в окрестности оз. Кискинлампи. По имеющимся данным, территория не подвергалась вырубкам в течении по крайней мере 100 лет и пожарам в течении 200
Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – – Календарного возраста особей Стадии жизненного цикла: Растения: семена – (p) проростки – (j) ювенильные – (im) имматурные – (v) виргинные (вегетативные) – (g) генеративные (молодые, средневозрастные, старые) – (ss) субсенильные – (s) сенильные. Демографическая структура популяций ели, березы и осины на пробной площади 0, 04 га в окрестности оз. Кискинлампи. По имеющимся данным, территория не подвергалась вырубкам в течении по крайней мере 100 лет и пожарам в течении 200
 Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – – Календарного возраста особей Стадии жизненного цикла: Растения: семена – (p) проростки – (j) ювенильные – (im) имматурные – (v) виргинные (вегетативные) – (g) генеративные (молодые, средневозрастные, старые) – (ss) субсенильные – (s) сенильные. Насекомые: яйца – личинки 1 возраста – лич. 1 возр. – …… – куколки – имаго
Динамика популяции, обладающей возрастной структурой • Возрастную структуру описывают с использованием: – – Календарного возраста особей Стадии жизненного цикла: Растения: семена – (p) проростки – (j) ювенильные – (im) имматурные – (v) виргинные (вегетативные) – (g) генеративные (молодые, средневозрастные, старые) – (ss) субсенильные – (s) сенильные. Насекомые: яйца – личинки 1 возраста – лич. 1 возр. – …… – куколки – имаго
. Таблица") Динамика популяции, обладающей возрастной структурой Демографические таблицы • • Таблица выживания (“Life Table”). Таблица размножения (плодовитости). Демографические таблицы, как правило, строятся для когорты и отражают ее судьбу от момента появления до отмирания последней особи. Это так называемые когортные, или динамические таблицы выживания и размножения.
Динамика популяции, обладающей возрастной структурой Демографические таблицы • • Таблица выживания (“Life Table”). Таблица размножения (плодовитости). Демографические таблицы, как правило, строятся для когорты и отражают ее судьбу от момента появления до отмирания последней особи. Это так называемые когортные, или динамические таблицы выживания и размножения.
 Возраст или стадия x") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия x (0) Яйцо (1) Лич-1 (2) Лич-2 (3) Лич-3 (4) Лич-4 (5) Имаго (6)
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия x (0) Яйцо (1) Лич-1 (2) Лич-2 (3) Лич-3 (4) Лич-4 (5) Имаго (6)
 Возраст или стадия Число") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста x ax (0) Яйцо 44000 (1) Лич-1 3513 (2) Лич-2 2529 (3) Лич-3 1922 (4) Лич-4 1461 (5) Имаго 1300 (6) 0
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста x ax (0) Яйцо 44000 (1) Лич-1 3513 (2) Лич-2 2529 (3) Лич-3 1922 (4) Лич-4 1461 (5) Имаго 1300 (6) 0
 Возраст или стадия Число") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста x ax lx (0) Яйцо 44000 1. 000 (1) Лич-1 3513 0. 080 (2) Лич-2 2529 0. 058 (3) Лич-3 1922 0. 044 (4) Лич-4 1461 0. 033 (5) Имаго 1300 0. 030 (6) 0 0
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста x ax lx (0) Яйцо 44000 1. 000 (1) Лич-1 3513 0. 080 (2) Лич-2 2529 0. 058 (3) Лич-3 1922 0. 044 (4) Лич-4 1461 0. 033 (5) Имаго 1300 0. 030 (6) 0 0
 Возраст или стадия Число") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте x ax lx dx (0) Яйцо 44000 1. 000 0. 920 (1) Лич-1 3513 0. 080 0. 022 (2) Лич-2 2529 0. 058 0. 014 (3) Лич-3 1922 0. 044 0. 011 (4) Лич-4 1461 0. 033 0. 003 (5) Имаго 1300 0. 030 ---- (6) 0 0 ---- Значения dx можно складывать, при этом:
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте x ax lx dx (0) Яйцо 44000 1. 000 0. 920 (1) Лич-1 3513 0. 080 0. 022 (2) Лич-2 2529 0. 058 0. 014 (3) Лич-3 1922 0. 044 0. 011 (4) Лич-4 1461 0. 033 0. 003 (5) Имаго 1300 0. 030 ---- (6) 0 0 ---- Значения dx можно складывать, при этом:
 Возраст или стадия Число") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте Коэффициент смертности x ax lx dx qx (0) Яйцо 44000 1. 000 0. 92 (1) Лич-1 3513 0. 080 0. 022 0. 28 (2) Лич-2 2529 0. 058 0. 014 0. 24 (3) Лич-3 1922 0. 044 0. 011 0. 25 (4) Лич-4 1461 0. 033 0. 003 0. 11 (5) Имаго 1300 0. 030 ---- (6) 0 0 ---- qx – средний шанс для особи погибнуть на данной стадии px = (1 – qx) – средний шанс для особи выжить на данной стадии Значения qx нельзя складывать
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте Коэффициент смертности x ax lx dx qx (0) Яйцо 44000 1. 000 0. 92 (1) Лич-1 3513 0. 080 0. 022 0. 28 (2) Лич-2 2529 0. 058 0. 014 0. 24 (3) Лич-3 1922 0. 044 0. 011 0. 25 (4) Лич-4 1461 0. 033 0. 003 0. 11 (5) Имаго 1300 0. 030 ---- (6) 0 0 ---- qx – средний шанс для особи погибнуть на данной стадии px = (1 – qx) – средний шанс для особи выжить на данной стадии Значения qx нельзя складывать
 Возраст или стадия Число") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте Коэффициент Интенсивность смертности x ax lx dx qx kx (0) Яйцо 44000 1. 000 0. 92 1. 098 (1) Лич-1 3513 0. 080 0. 022 0. 28 0. 15 (2) Лич-2 2529 0. 058 0. 014 0. 24 0. 12 (3) Лич-3 1922 0. 044 0. 011 0. 25 0. 12 (4) Лич-4 1461 0. 033 0. 003 0. 11 0. 05 (5) Имаго 1300 0. 030 ---- (6) 0 0 ----
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте Коэффициент Интенсивность смертности x ax lx dx qx kx (0) Яйцо 44000 1. 000 0. 92 1. 098 (1) Лич-1 3513 0. 080 0. 022 0. 28 0. 15 (2) Лич-2 2529 0. 058 0. 014 0. 24 0. 12 (3) Лич-3 1922 0. 044 0. 011 0. 25 0. 12 (4) Лич-4 1461 0. 033 0. 003 0. 11 0. 05 (5) Имаго 1300 0. 030 ---- (6) 0 0 ----
 Возраст или стадия Число") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте x ax lx dx qx kx (0) Яйцо 44000 1. 000 0. 92 1. 098 (1) Лич-1 3513 0. 080 0. 022 0. 28 0. 15 (2) Лич-2 2529 0. 058 0. 014 0. 24 0. 12 (3) Лич-3 1922 0. 044 0. 011 0. 25 0. 12 (4) Лич-4 1461 0. 033 0. 003 0. 11 0. 05 (5) Имаго 1300 0. 030 ---- (6) 0 0 ---- Значения kx можно складывать, например: Коэффициент Интенсивность смертности
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus) Возраст или стадия Число особей в начале возраста Доля когорты, дожившая до начала возраста Доля погибших в данном возрасте x ax lx dx qx kx (0) Яйцо 44000 1. 000 0. 92 1. 098 (1) Лич-1 3513 0. 080 0. 022 0. 28 0. 15 (2) Лич-2 2529 0. 058 0. 014 0. 24 0. 12 (3) Лич-3 1922 0. 044 0. 011 0. 25 0. 12 (4) Лич-4 1461 0. 033 0. 003 0. 11 0. 05 (5) Имаго 1300 0. 030 ---- (6) 0 0 ---- Значения kx можно складывать, например: Коэффициент Интенсивность смертности
 • • Динамическая таблица размножения") Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) • • Динамическая таблица размножения cоставляется на основе когортной таблицы выживания и является ее продолжением. Статическая таблица размножения может быть составлена для всей популяции на основе данных о возрастном составе и средней плодовитости особей разного возраста.
Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) • • Динамическая таблица размножения cоставляется на основе когортной таблицы выживания и является ее продолжением. Статическая таблица размножения может быть составлена для всей популяции на основе данных о возрастном составе и средней плодовитости особей разного возраста.
 Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Пальма Astrocaryum mexicanum
Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Пальма Astrocaryum mexicanum
 Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Мятлик Poa annua
Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Мятлик Poa annua
 Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Белохвостый олень Odocoileus hemionus в штате Мичиган
Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Белохвостый олень Odocoileus hemionus в штате Мичиган
 Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Красный олень Cervus elaphus
Динамика популяции, обладающей возрастной структурой Примеры возрастной динамики средней плодовитости особей Красный олень Cervus elaphus
 Возраст Доля когорты, дожившая до") Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая до начала возраста x x lx 0 1 2 3 4 5 6 Σ
Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая до начала возраста x x lx 0 1 2 3 4 5 6 Σ
 Возраст Доля когорты, дожившая до") Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая до начала возраста x x lx 0 1 0. 8 2 0. 6 3 0. 4 4 0. 4 5 0. 2 6 0. 0 Σ ----
Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая до начала возраста x x lx 0 1 0. 8 2 0. 6 3 0. 4 4 0. 4 5 0. 2 6 0. 0 Σ ----
 Возраст Доля когорты, дожившая Средняя") Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая Средняя плодовитость до начала возраста x в возрасте x x lx mx 0 1. 0 0. 0 1 0. 8 0. 2 2 0. 6 0. 3 3 0. 4 1. 0 4 0. 6 5 0. 2 0. 1 6 0. 0 Σ ---- 2. 2 Валовая рождаемость – среднее число потомков на 1 особь, которые могли бы появиться, если бы все особи доживали до максимально возможного возраста.
Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая Средняя плодовитость до начала возраста x в возрасте x x lx mx 0 1. 0 0. 0 1 0. 8 0. 2 2 0. 6 0. 3 3 0. 4 1. 0 4 0. 6 5 0. 2 0. 1 6 0. 0 Σ ---- 2. 2 Валовая рождаемость – среднее число потомков на 1 особь, которые могли бы появиться, если бы все особи доживали до максимально возможного возраста.
 Возраст Доля когорты, дожившая Средняя") Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая Средняя плодовитость до начала возраста x в возрасте x Среднее число потомков на особь в возрасте x x lx mx l x mx 0 1. 0 0. 00 1 0. 8 0. 2 0. 16 2 0. 6 0. 3 0. 18 3 0. 4 1. 0 0. 40 4 0. 6 0. 24 5 0. 2 0. 1 0. 02 6 0. 00 Σ ---- 2. 2 1. 00 Чистая скорость размножения (воспроизводства) – среднее число потомков, оставляемых одной особью в течение жизни с учетом динамики вымирания когорты.
Динамика популяции, обладающей возрастной структурой Возрастная таблица размножения (плодовитости) Возраст Доля когорты, дожившая Средняя плодовитость до начала возраста x в возрасте x Среднее число потомков на особь в возрасте x x lx mx l x mx 0 1. 0 0. 00 1 0. 8 0. 2 0. 16 2 0. 6 0. 3 0. 18 3 0. 4 1. 0 0. 40 4 0. 6 0. 24 5 0. 2 0. 1 0. 02 6 0. 00 Σ ---- 2. 2 1. 00 Чистая скорость размножения (воспроизводства) – среднее число потомков, оставляемых одной особью в течение жизни с учетом динамики вымирания когорты.
") Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus)
Основные параметры когортной таблицы выживания (на примере саранчевого Chortippus brunneus)
 Основные параметры когортной таблицы выживания
Основные параметры когортной таблицы выживания
 – среднее число потомков, оставляемых") Динамика популяции, обладающей возрастной структурой Чистая скорость размножения (воспроизводства) – среднее число потомков, оставляемых одной особью в течение жизни с учетом динамики вымирания когорты. – – Имеет размерность числа особей Если R 0 > 1 – популяция растет Если R 0 = 1 – популяция не изменяет свою численность Если R 0 < 1 – популяция сокращается Поэтому R 0 называют также “скоростью замещения популяции”. Так, если в поколении 0 было N 0 особей, то в поколении 1 их будет: Откуда следует: В поколении 2: В общем виде: (где F – номер поколения)
Динамика популяции, обладающей возрастной структурой Чистая скорость размножения (воспроизводства) – среднее число потомков, оставляемых одной особью в течение жизни с учетом динамики вымирания когорты. – – Имеет размерность числа особей Если R 0 > 1 – популяция растет Если R 0 = 1 – популяция не изменяет свою численность Если R 0 < 1 – популяция сокращается Поэтому R 0 называют также “скоростью замещения популяции”. Так, если в поколении 0 было N 0 особей, то в поколении 1 их будет: Откуда следует: В поколении 2: В общем виде: (где F – номер поколения)
 Динамика популяции, обладающей возрастной структурой Таблица выживания и плодовитости Phlox drummondii
Динамика популяции, обладающей возрастной структурой Таблица выживания и плодовитости Phlox drummondii
 Динамика популяции, обладающей возрастной структурой • В тех случаях, когда невозможно построить таблицу выживания для когорты, используют данные о возрастной структуре популяции. Ясно, что число особей разного возраста примерно отражает усредненную динамику вымирания когорт и позволяет построить таблицу выживания. Такую таблицу выживания называют статической, а также моментальной, или вертикальной. Возрастная пирамида для самцов и самок чернохвостого оленя в Чаппарале Калифорнии (по Дажо, 1975)
Динамика популяции, обладающей возрастной структурой • В тех случаях, когда невозможно построить таблицу выживания для когорты, используют данные о возрастной структуре популяции. Ясно, что число особей разного возраста примерно отражает усредненную динамику вымирания когорт и позволяет построить таблицу выживания. Такую таблицу выживания называют статической, а также моментальной, или вертикальной. Возрастная пирамида для самцов и самок чернохвостого оленя в Чаппарале Калифорнии (по Дажо, 1975)
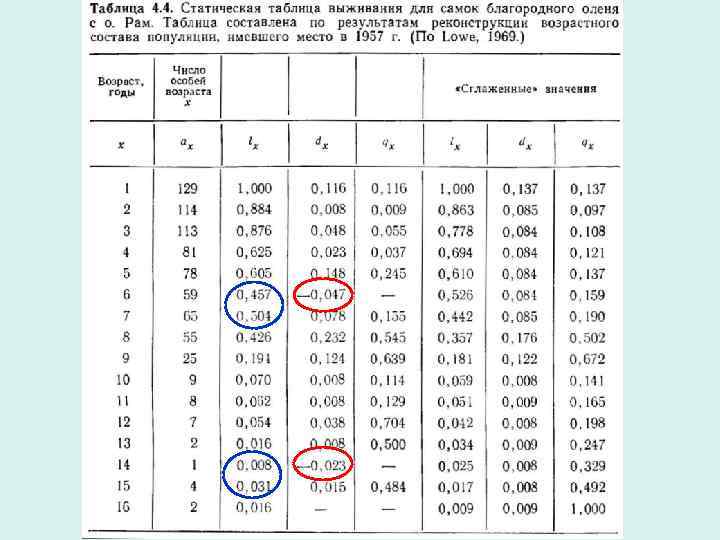
 Динамика популяции, обладающей возрастной структурой Кривые выживания • Можно построить на основе когортных таблиц выживания. Поэтому их называют когортными кривыми выживания. Первый метод построения – в арифметическом масштабе: – – – Используют абсолютные значения численности и возраста Этот метод не позволяет сравнивать популяции с различной численностью и долговечностью особей Форма таких кривых мало что говорит о характере смертности особей Число особей ax • Возраст x
Динамика популяции, обладающей возрастной структурой Кривые выживания • Можно построить на основе когортных таблиц выживания. Поэтому их называют когортными кривыми выживания. Первый метод построения – в арифметическом масштабе: – – – Используют абсолютные значения численности и возраста Этот метод не позволяет сравнивать популяции с различной численностью и долговечностью особей Форма таких кривых мало что говорит о характере смертности особей Число особей ax • Возраст x
 Динамика популяции, обладающей возрастной структурой Кривые выживания • Второй способ построения – в полулогарифмическом масштабе: – – Число особей выражают в логарифмическом масштабе по оси ординат. Поэтому наклон кривой относительно оси абсцисс в каждой точке пропорционален вероятности гибели особей в данном возрасте. Используют относительные значения численности (пересчет на 1000 особей) и возраста (в процентах от максимальной долговечности особей). Это позволяет легко сравнивать популяции с различной численностью и долговечностью особей. 1000 особей Наклон кривой Log ax Макс. возраст = 1
Динамика популяции, обладающей возрастной структурой Кривые выживания • Второй способ построения – в полулогарифмическом масштабе: – – Число особей выражают в логарифмическом масштабе по оси ординат. Поэтому наклон кривой относительно оси абсцисс в каждой точке пропорционален вероятности гибели особей в данном возрасте. Используют относительные значения численности (пересчет на 1000 особей) и возраста (в процентах от максимальной долговечности особей). Это позволяет легко сравнивать популяции с различной численностью и долговечностью особей. 1000 особей Наклон кривой Log ax Макс. возраст = 1
: 1. Тип") Динамика популяции, обладающей возрастной структурой Три основных типа кривых выживания (Pearl, 1928): 1. Тип дрозофилы, или человека – выпуклая кривая • 2. Вероятность гибели особей увеличивается с возрастом. Тип гидры – прямая линия • 3. Вероятность гибели особей постоянна и не зависит от возраста. Тип устрицы – вогнутая кривая – Вероятность гибели особей уменьшается с возрастом.
Динамика популяции, обладающей возрастной структурой Три основных типа кривых выживания (Pearl, 1928): 1. Тип дрозофилы, или человека – выпуклая кривая • 2. Вероятность гибели особей увеличивается с возрастом. Тип гидры – прямая линия • 3. Вероятность гибели особей постоянна и не зависит от возраста. Тип устрицы – вогнутая кривая – Вероятность гибели особей уменьшается с возрастом.
: • Кривые") Динамика популяции, обладающей возрастной структурой Три основных типа кривых выживания (Pearl, 1928): • Кривые выживания для реальных видов бывают очень различны и могут лишь приближаться к тому или иному типу. • Нередко они являются комбинацией разных типов кривых, особенно в тех случаях, когда характер смертности особей на разных стадиях жизненного цикла очень различен. Кривая выживания, типичная для высших позвоночных: Повышенная смертность детенышей
Динамика популяции, обладающей возрастной структурой Три основных типа кривых выживания (Pearl, 1928): • Кривые выживания для реальных видов бывают очень различны и могут лишь приближаться к тому или иному типу. • Нередко они являются комбинацией разных типов кривых, особенно в тех случаях, когда характер смертности особей на разных стадиях жизненного цикла очень различен. Кривая выживания, типичная для высших позвоночных: Повышенная смертность детенышей
: • Кривые") Динамика популяции, обладающей возрастной структурой Три основных типа кривых выживания (Pearl, 1928): • Кривые выживания для реальных видов бывают очень различны и могут лишь приближаться к тому или иному типу. • Нередко они являются комбинацией разных типов кривых, особенно в тех случаях, когда характер смертности особей на разных стадиях жизненного цикла очень различен. Кривая выживания, типичная для насекомых: Выживание яиц и личинок Выживание имаго
Динамика популяции, обладающей возрастной структурой Три основных типа кривых выживания (Pearl, 1928): • Кривые выживания для реальных видов бывают очень различны и могут лишь приближаться к тому или иному типу. • Нередко они являются комбинацией разных типов кривых, особенно в тех случаях, когда характер смертности особей на разных стадиях жизненного цикла очень различен. Кривая выживания, типичная для насекомых: Выживание яиц и личинок Выживание имаго
 Примеры кривых выживания Phlox drummondii • В конце стадии семени кривая выживания вогнута, а в конце периода существования поколения - выпукла • Построение кривых выживания – это наиболее употребительный способ изображения динамики смертности
Примеры кривых выживания Phlox drummondii • В конце стадии семени кривая выживания вогнута, а в конце периода существования поколения - выпукла • Построение кривых выживания – это наиболее употребительный способ изображения динамики смертности
 Примеры кривых выживания
Примеры кривых выживания
 Динамика популяции, обладающей возрастной структурой Роль "смертности до рождения" в динамике популяций растений и животных • У человека: – • до рождения доживает лишь около 31% зигот. У растений: – доля “выкидышей” составляет около 15% у однолетних и до 50% у многолетних видов.
Динамика популяции, обладающей возрастной структурой Роль "смертности до рождения" в динамике популяций растений и животных • У человека: – • до рождения доживает лишь около 31% зигот. У растений: – доля “выкидышей” составляет около 15% у однолетних и до 50% у многолетних видов.
 Динамика популяции, обладающей возрастной структурой Точное и приближенное вычисление удельной мгновенной скорости роста популяции А. Точное вычисление – по таблице размножения путем решения уравнения Эйлера (Lotka, 1907): – – – Это так называемое «неявное уравнение» Его можно решить только путем последовательных итераций Недостаток уравнения в том, что оно лишено сколько-нибудь ясного биологического смысла
Динамика популяции, обладающей возрастной структурой Точное и приближенное вычисление удельной мгновенной скорости роста популяции А. Точное вычисление – по таблице размножения путем решения уравнения Эйлера (Lotka, 1907): – – – Это так называемое «неявное уравнение» Его можно решить только путем последовательных итераций Недостаток уравнения в том, что оно лишено сколько-нибудь ясного биологического смысла
 Динамика популяции, обладающей возрастной структурой Точное и приближенное вычисление удельной мгновенной скорости роста популяции Б. Приближенное вычисление Пусть t = T – длительность одного поколения Тогда: Отсюда: Но и если условия стабильны, то Следовательно: После логарифмирования: Или иначе:
Динамика популяции, обладающей возрастной структурой Точное и приближенное вычисление удельной мгновенной скорости роста популяции Б. Приближенное вычисление Пусть t = T – длительность одного поколения Тогда: Отсюда: Но и если условия стабильны, то Следовательно: После логарифмирования: Или иначе:
 Динамика популяции, обладающей возрастной структурой Точное и приближенное вычисление удельной мгновенной скорости роста популяции Б. Приближенное вычисление Более правильно: – когортное время генерации (=продолжительность поколения) Эта формула справедлива: – – – в условиях, близких к равновесию когда длительность жизни особей и продолжительность поколения постоянны когда популяция стабильна, либо изменяет свою численность достаточно медленно
Динамика популяции, обладающей возрастной структурой Точное и приближенное вычисление удельной мгновенной скорости роста популяции Б. Приближенное вычисление Более правильно: – когортное время генерации (=продолжительность поколения) Эта формула справедлива: – – – в условиях, близких к равновесию когда длительность жизни особей и продолжительность поколения постоянны когда популяция стабильна, либо изменяет свою численность достаточно медленно
 Динамика популяции, обладающей возрастной структурой Пример вычисления параметров выживания и размножения для популяции
Динамика популяции, обладающей возрастной структурой Пример вычисления параметров выживания и размножения для популяции
 Динамика популяции, обладающей возрастной структурой Когортное время генерации Это средне-взвешенный возраст особи, в котором она производит потомство: Более общая формула учитывает динамику вымирания и размножения всей когорты:
Динамика популяции, обладающей возрастной структурой Когортное время генерации Это средне-взвешенный возраст особи, в котором она производит потомство: Более общая формула учитывает динамику вымирания и размножения всей когорты:
 Динамика популяции, обладающей возрастной структурой Когортное время генерации
Динамика популяции, обладающей возрастной структурой Когортное время генерации
 Динамика популяции, обладающей возрастной структурой Когортное время генерации Зависимость времени генерации от размера организмов
Динамика популяции, обладающей возрастной структурой Когортное время генерации Зависимость времени генерации от размера организмов
 Ограниченный рост популяции Действие факторов, ограничивающих рост популяции, обычно зависит от численности популяции таким образом, что: – – при увеличении численности смертность увеличивается, а рождаемость падает; это ведет к уменьшению скорости роста популяции вплоть до полного его прекращения.
Ограниченный рост популяции Действие факторов, ограничивающих рост популяции, обычно зависит от численности популяции таким образом, что: – – при увеличении численности смертность увеличивается, а рождаемость падает; это ведет к уменьшению скорости роста популяции вплоть до полного его прекращения.
 Ограниченный рост популяции Логистическое уравнение роста популяции • • Пусть удельная рождаемость ( b ) и смертность ( d ) зависят от численности линейно. При некоторой численности популяции b и d становятся равными другу. Обозначим эту численность буквой K. Тогда мгновенная удельная скорость роста популяции ( r ) также будет уменьшаться линейно. Удельная скорость роста • r=b-d d b K N
Ограниченный рост популяции Логистическое уравнение роста популяции • • Пусть удельная рождаемость ( b ) и смертность ( d ) зависят от численности линейно. При некоторой численности популяции b и d становятся равными другу. Обозначим эту численность буквой K. Тогда мгновенная удельная скорость роста популяции ( r ) также будет уменьшаться линейно. Удельная скорость роста • r=b-d d b K N
 Ограниченный рост популяции Логистическое уравнение роста популяции • • Пусть удельная рождаемость ( b ) и смертность ( d ) зависят от численности линейно. При некоторой численности популяции b и d становятся равными другу. Обозначим эту численность буквой K. Тогда мгновенная удельная скорость роста популяции ( r ) также будет уменьшаться линейно. Удельная скорость роста • r=b-d При N = K, r = 0 r K N
Ограниченный рост популяции Логистическое уравнение роста популяции • • Пусть удельная рождаемость ( b ) и смертность ( d ) зависят от численности линейно. При некоторой численности популяции b и d становятся равными другу. Обозначим эту численность буквой K. Тогда мгновенная удельная скорость роста популяции ( r ) также будет уменьшаться линейно. Удельная скорость роста • r=b-d При N = K, r = 0 r K N
 Ограниченный рост популяции Логистическое уравнение роста популяции • Это простое предположение приводит к классическому уравнению Ферхюльста-Пёрла: – Verhulst, 1838 – Pearl & Read, 1920 Pierre François Verhulst (1804 – 1849), бельгийский математик Raymond Pearl (1879 – 1940) American biologist
Ограниченный рост популяции Логистическое уравнение роста популяции • Это простое предположение приводит к классическому уравнению Ферхюльста-Пёрла: – Verhulst, 1838 – Pearl & Read, 1920 Pierre François Verhulst (1804 – 1849), бельгийский математик Raymond Pearl (1879 – 1940) American biologist
 Ограниченный рост популяции Логистическое уравнение роста популяции • Это простое предположение приводит к классическому уравнению Ферхюльста-Пёрла: – Verhulst, 1838 – Pearl & Read, 1920 Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
Ограниченный рост популяции Логистическое уравнение роста популяции • Это простое предположение приводит к классическому уравнению Ферхюльста-Пёрла: – Verhulst, 1838 – Pearl & Read, 1920 Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
 Ограниченный рост популяции Логистическое уравнение роста популяции • Это простое предположение приводит к классическому уравнению Ферхюльста-Пёрла: – Verhulst, 1838 – Pearl & Read, 1920 Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
Ограниченный рост популяции Логистическое уравнение роста популяции • Это простое предположение приводит к классическому уравнению Ферхюльста-Пёрла: – Verhulst, 1838 – Pearl & Read, 1920 Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
 Ограниченный рост популяции Логистическое уравнение роста популяции • Уравнение Ферхюльста-Пёрла описывает S-образный, или сигмоидный, или иначе логистический рост популяции. Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
Ограниченный рост популяции Логистическое уравнение роста популяции • Уравнение Ферхюльста-Пёрла описывает S-образный, или сигмоидный, или иначе логистический рост популяции. Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
 Ограниченный рост популяции Логистическое уравнение роста популяции • Разные формы логистического уравнения: 1 или 2 3 4 где – экспоненциальное уравнение – “сопротивление среды”
Ограниченный рост популяции Логистическое уравнение роста популяции • Разные формы логистического уравнения: 1 или 2 3 4 где – экспоненциальное уравнение – “сопротивление среды”
 Ограниченный рост популяции Логистическое уравнение роста популяции • Логистическое уравнение после интегрирования: где K – Предельная плотность популяции, потенциальная емкость среды обитания
Ограниченный рост популяции Логистическое уравнение роста популяции • Логистическое уравнение после интегрирования: где K – Предельная плотность популяции, потенциальная емкость среды обитания
 Ограниченный рост популяции Логистическое уравнение роста популяции • Логистическое уравнение после интегрирования Сопротивление среды
Ограниченный рост популяции Логистическое уравнение роста популяции • Логистическое уравнение после интегрирования Сопротивление среды
 Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – – – когда t = 0, Nt = K / (1+b) = N 0 когда Nt = ½K – скорость роста d. N/dt максимальна когда t стремится к бесконечности, Nt стремится к K Это перегиб кривой Скорость роста увеличивается Скорость роста уменьшается
Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – – – когда t = 0, Nt = K / (1+b) = N 0 когда Nt = ½K – скорость роста d. N/dt максимальна когда t стремится к бесконечности, Nt стремится к K Это перегиб кривой Скорость роста увеличивается Скорость роста уменьшается
 Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – Изменение абсолютной скорости роста популяции Скорость роста увеличивается Скорость роста уменьшается N
Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – Изменение абсолютной скорости роста популяции Скорость роста увеличивается Скорость роста уменьшается N
 Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – При увеличении значения r кривая выходит на плато быстрее
Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – При увеличении значения r кривая выходит на плато быстрее
 Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – Если исходная численность N 0 > K, то b < 0 и r < 0, и популяция будет уменьшаться
Ограниченный рост популяции Логистическое уравнение роста популяции • Анализ уравнения – Если исходная численность N 0 > K, то b < 0 и r < 0, и популяция будет уменьшаться
 Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения – – – простота; удобство использования в других более сложных моделях; получило широчайшее распространение в популяционной экологии.
Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения – – – простота; удобство использования в других более сложных моделях; получило широчайшее распространение в популяционной экологии.
 Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения • Недостатки логистического уравнения – – – чрезмерно упрощает биологические явления; все особи в популяции эквивалентны; добавление одной особи при любой плотности уменьшает скорость роста на одну и ту же величину – 1/K; реакция популяции на возрастание или уменьшение плотности является мгновенной; линейная зависимость удельной мгновенной скорости роста r от численности – нереалистична; значение K и максимальная величина r считаются константами;
Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения • Недостатки логистического уравнения – – – чрезмерно упрощает биологические явления; все особи в популяции эквивалентны; добавление одной особи при любой плотности уменьшает скорость роста на одну и ту же величину – 1/K; реакция популяции на возрастание или уменьшение плотности является мгновенной; линейная зависимость удельной мгновенной скорости роста r от численности – нереалистична; значение K и максимальная величина r считаются константами;
 Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения • Недостатки логистического уравнения • Несоответствие динамики роста природных популяций логистической модели
Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения • Недостатки логистического уравнения • Несоответствие динамики роста природных популяций логистической модели
 Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
 Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
 Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
 Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте
 Примеры S-образного роста популяций в природе и эксперименте
Примеры S-образного роста популяций в природе и эксперименте
 Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте Реальный рост населения США в XX столетии Теоретическая кривая роста, рассчитанная в 1920 г. по логистическому уравнению Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
Ограниченный рост популяции Примеры S-образного роста популяций в природе и эксперименте Реальный рост населения США в XX столетии Теоретическая кривая роста, рассчитанная в 1920 г. по логистическому уравнению Рост населения США в XIX и начале XX столетия (по Pearl & Read, 1920)
 Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения • Недостатки логистического уравнения • Несоответствие динамики роста природных популяций логистической модели • Другие модели ограниченного роста популяции
Ограниченный рост популяции Логистическое уравнение роста популяции Ферхюльста-Пёрла • Популярность логистического уравнения • Недостатки логистического уравнения • Несоответствие динамики роста природных популяций логистической модели • Другие модели ограниченного роста популяции