

лекция морфология.ppt
- Количество слайдов: 80
 11. Негативное контрастирование Streptococcus pyogenes при электронной микроскопии (28, 000 X). Ореол вокруг") Число(фигура) 11. Негативное контрастирование Streptococcus pyogenes при электронной микроскопии (28, 000 X). Ореол вокруг цепочки клеток - капсула из гиалуроновой кислоты, которая окружает бактерии с внешней стороны. Также может быть замечена септа между делящейся парой клеток.
Число(фигура) 11. Негативное контрастирование Streptococcus pyogenes при электронной микроскопии (28, 000 X). Ореол вокруг цепочки клеток - капсула из гиалуроновой кислоты, которая окружает бактерии с внешней стороны. Также может быть замечена септа между делящейся парой клеток.
 Колонии Bacillus anthracis. Рост слизистых или мукоидных бактериальных колоний - обычно свидетельствует о продукции капсул. В случае B. anthracis, капсула состоит из поли-D-глутамина. Капсула - существенный детерминант патогенности бактерий. На ранних стадиях колонизации и инфекции капсула защищает бактерии от антибактериальной активности иммунной и фагоцитарной систем.
Колонии Bacillus anthracis. Рост слизистых или мукоидных бактериальных колоний - обычно свидетельствует о продукции капсул. В случае B. anthracis, капсула состоит из поли-D-глутамина. Капсула - существенный детерминант патогенности бактерий. На ранних стадиях колонизации и инфекции капсула защищает бактерии от антибактериальной активности иммунной и фагоцитарной систем.
 Микроорганизм Природа капсулы Субъединицы капсульного полимера Acetobacter xylinum Целлюлоза Глюкоза Azotobacter vinelandii Полиуронид Глюкуроновая и маннуроновая кислоты Bac. antracis Полипептид D-Глутоминовая кислота Bac. licheniformis Отдельные виды из семейства Enterobacteriacceae Многие типы сложных полисахаридов, колановая кислота Галактоза, глюкуроновая кислота, ПВК, фукоза и др. Klebsiella pneumoniae Cложный полисахарид Leuconostoc mesenteroides Глюкан (декстран) Галактоза, галактуроновая кислота, фукоза Глюкоза Pseudomonas aerugenosa Полиуронид или другие полисахариды Гиалуроновая кислота Streptococcus haemoliticus Streptococcus pyogenes Sterptococcus pneumoniae Многие типы сложных полимеров, например: Тип I Тип II Sterptococcus salivarius Фруктан (леван) N. meningitidis Полисахарид H. influenzae Полисахарид Глюкуроновая. Маннуроновая кислота N-ацетилглюкозамин, глюкуроновая кислота 3 -Дезоксигалактоза, галактуроновая кислота, глюкоза, глюкуроновая кислота Фруктоза полимер N-ацетилманнозамина фосфата (группа А); полимер сиаловой кислоты (группа B и С) Полирибозфосфат
Микроорганизм Природа капсулы Субъединицы капсульного полимера Acetobacter xylinum Целлюлоза Глюкоза Azotobacter vinelandii Полиуронид Глюкуроновая и маннуроновая кислоты Bac. antracis Полипептид D-Глутоминовая кислота Bac. licheniformis Отдельные виды из семейства Enterobacteriacceae Многие типы сложных полисахаридов, колановая кислота Галактоза, глюкуроновая кислота, ПВК, фукоза и др. Klebsiella pneumoniae Cложный полисахарид Leuconostoc mesenteroides Глюкан (декстран) Галактоза, галактуроновая кислота, фукоза Глюкоза Pseudomonas aerugenosa Полиуронид или другие полисахариды Гиалуроновая кислота Streptococcus haemoliticus Streptococcus pyogenes Sterptococcus pneumoniae Многие типы сложных полимеров, например: Тип I Тип II Sterptococcus salivarius Фруктан (леван) N. meningitidis Полисахарид H. influenzae Полисахарид Глюкуроновая. Маннуроновая кислота N-ацетилглюкозамин, глюкуроновая кислота 3 -Дезоксигалактоза, галактуроновая кислота, глюкоза, глюкуроновая кислота Фруктоза полимер N-ацетилманнозамина фосфата (группа А); полимер сиаловой кислоты (группа B и С) Полирибозфосфат
 Бактерии Состав капсул Структурные компоненты Gram-positive Bacteria Bacillus anthracis Полипептид D-глутаминовая кислота Bacillus megaterium Полипептид и полисахарид Streptococcus mutans Полисахарид D-глутаминовая кислота, аминосахара, сахара (декстран) глюкоза Streptococcus pneumoniae Streptococcus pyogenes Gram-negative Bacteria Acetobacter xylinum Полисахарид сахара, аминосахара, уроновые кислоты полисахарид (гиалуроновая кислота) N-ацетил-глюкозамин и глюкуроновая кислота Полисахарид (целлюлоза) глюкоза Escherichia coli полисахарид кислота) Полисахарид Pseudomonas aeruginosa Azotobacter vinelandii Agrobacterium tumefaciens (колановая глюкоза, галактоза, глюкуроновая кислота Маннуроновая кислота Полисахарид глюкуроновая кислота полисахарид (глюкан) глюкоза фукоза,
Бактерии Состав капсул Структурные компоненты Gram-positive Bacteria Bacillus anthracis Полипептид D-глутаминовая кислота Bacillus megaterium Полипептид и полисахарид Streptococcus mutans Полисахарид D-глутаминовая кислота, аминосахара, сахара (декстран) глюкоза Streptococcus pneumoniae Streptococcus pyogenes Gram-negative Bacteria Acetobacter xylinum Полисахарид сахара, аминосахара, уроновые кислоты полисахарид (гиалуроновая кислота) N-ацетил-глюкозамин и глюкуроновая кислота Полисахарид (целлюлоза) глюкоза Escherichia coli полисахарид кислота) Полисахарид Pseudomonas aeruginosa Azotobacter vinelandii Agrobacterium tumefaciens (колановая глюкоза, галактоза, глюкуроновая кислота Маннуроновая кислота Полисахарид глюкуроновая кислота полисахарид (глюкан) глюкоза фукоза,
 Бактериальные капсулы, контрастированные китайской тушью, рассматриваемые в световой микроскоп. Это - истинная капсула, обособленный слой полисахаридов, вокруг клеток. Иногда бактериальные клетки окружены организованным более беспорядочно полисахаридным матриксом, называемым слизью или биопленкой. Полисахаридные образования, которые могут неизбежно присутствовать на поверхностях бактериальных клеток, но которые не могут быть обнаружены визуально, называются гликокаликс.
Бактериальные капсулы, контрастированные китайской тушью, рассматриваемые в световой микроскоп. Это - истинная капсула, обособленный слой полисахаридов, вокруг клеток. Иногда бактериальные клетки окружены организованным более беспорядочно полисахаридным матриксом, называемым слизью или биопленкой. Полисахаридные образования, которые могут неизбежно присутствовать на поверхностях бактериальных клеток, но которые не могут быть обнаружены визуально, называются гликокаликс.
 Рибитол тейхоевая кислота с") Figure 2 -9. Structures of cell wall teichoic acids. (A) Рибитол тейхоевая кислота с повторяющимися фрагментами связанными 1, 5 фосфодиэфирными связями D-рибитола и D-аланилового эфира в позиции 2 и гликозильные радикалы (R) в позиции 4. Гликозильными группами могут быть N-acetylglucosaminyl (a or b) как у S aureus или a-glucosyl как у B subtilis W 23. (B) Глицерол тейхоевая кислота с 1, 3 - фосфодиэфирными связями между повторяющимися глицерольными субъединицами (1, 2 -связи у некоторых видов). В качестве радикалов могут выступать (R - D-alanyl or glycosyl).
Figure 2 -9. Structures of cell wall teichoic acids. (A) Рибитол тейхоевая кислота с повторяющимися фрагментами связанными 1, 5 фосфодиэфирными связями D-рибитола и D-аланилового эфира в позиции 2 и гликозильные радикалы (R) в позиции 4. Гликозильными группами могут быть N-acetylglucosaminyl (a or b) как у S aureus или a-glucosyl как у B subtilis W 23. (B) Глицерол тейхоевая кислота с 1, 3 - фосфодиэфирными связями между повторяющимися глицерольными субъединицами (1, 2 -связи у некоторых видов). В качестве радикалов могут выступать (R - D-alanyl or glycosyl).
 Строение клеточной стенки Грам-отрицательных бактерий
Строение клеточной стенки Грам-отрицательных бактерий
 О-антиген, эндотоксин
О-антиген, эндотоксин
 О-антиген, эндотоксин •
О-антиген, эндотоксин •
 Общая структура ЛПС V. cholerae") Общая структура ЛПС V. cholerae O 1 • (a) Общая структура ЛПС V. cholerae O 1, серотипов Ogawa (R = CH 3) и Inaba (R = H). O-SP (О-специфическая полисахаридная цепь) содержит последовательность из 2– 18 моносахаридных повторяющихся единиц α (1, 2)-связанных d-перозамина (4 -амино-4, 6 -дидезокси-d-манноза) амино группа которых ацилирована 3 дезокси-l-глицеро-тетроновой кислотой. (b) Ogawa терминальный дисахаридный фрагмент © 2000 by National Academy of Sciences
Общая структура ЛПС V. cholerae O 1 • (a) Общая структура ЛПС V. cholerae O 1, серотипов Ogawa (R = CH 3) и Inaba (R = H). O-SP (О-специфическая полисахаридная цепь) содержит последовательность из 2– 18 моносахаридных повторяющихся единиц α (1, 2)-связанных d-перозамина (4 -амино-4, 6 -дидезокси-d-манноза) амино группа которых ацилирована 3 дезокси-l-глицеро-тетроновой кислотой. (b) Ogawa терминальный дисахаридный фрагмент © 2000 by National Academy of Sciences
 Figure 2 -10. Три главных, связанных ковалентно, региона формирующих типичные ЛПС. Липид А Кор О антиген Глюкозамин Β-гидроксимиристиновая кислота Жирные кислоты Кетодезоксиоктонат Фосфоэтаноламин Гептоза Глюкоза Галактоза N–ацитилглюкозамин Полисахаридные цепи: повторяющиеся субъединицы видоспецифичных моносахаридов, например у сальмонелл галактоза, рамноза и манноза и абеквоза в ЛПС у S. typhimurium
Figure 2 -10. Три главных, связанных ковалентно, региона формирующих типичные ЛПС. Липид А Кор О антиген Глюкозамин Β-гидроксимиристиновая кислота Жирные кислоты Кетодезоксиоктонат Фосфоэтаноламин Гептоза Глюкоза Галактоза N–ацитилглюкозамин Полисахаридные цепи: повторяющиеся субъединицы видоспецифичных моносахаридов, например у сальмонелл галактоза, рамноза и манноза и абеквоза в ЛПС у S. typhimurium
 Функция Барьер проницамости") Table 4. Функция компонентов наружной мембраны Escherichia coli. Компонент Липополисахарид (ЛПС) Функция Барьер проницамости Mg++ мостики Стабилизация ЛПС и важный проницаемость наружной мембраны компонент характеризующий Липопротеин Braun Якорь для наружной мембраны фиксирующий ее к пептидогликану Omp C и Omp F Протеины, формирующие поры или каналы через наружную мембрану порины для прохождения гидрофильных молекул Omp A протеин Рецептор для некоторых вирусов и бактериоцинов; стабилизирует стыковку клеток во время коньюгации
Table 4. Функция компонентов наружной мембраны Escherichia coli. Компонент Липополисахарид (ЛПС) Функция Барьер проницамости Mg++ мостики Стабилизация ЛПС и важный проницаемость наружной мембраны компонент характеризующий Липопротеин Braun Якорь для наружной мембраны фиксирующий ее к пептидогликану Omp C и Omp F Протеины, формирующие поры или каналы через наружную мембрану порины для прохождения гидрофильных молекул Omp A протеин Рецептор для некоторых вирусов и бактериоцинов; стабилизирует стыковку клеток во время коньюгации
 Для") Функции периплазматических белков у E. coli. Связывающие протеины Для аминокислот (например, гистидина, аргинина) Для сахаров (например, глюкозы, мальтозы) Для витаминов (например, тиамина, витамина B 12) Для ионов (например, фосфатов, сульфатов) Биосинтетические ферменты Для сборки муреина (трансгликозидазы, карбоксипептидазы, транспептидазы) Для секреции и сборки жгутиковых субъединиц (например, шапиронины) Деградативные ферменты Фосфатазы Протеазы Детоксифицирующие ферменты β-лактамазы (пенициллиназа) Аминогликозид-фосфорилирующие ферменты
Функции периплазматических белков у E. coli. Связывающие протеины Для аминокислот (например, гистидина, аргинина) Для сахаров (например, глюкозы, мальтозы) Для витаминов (например, тиамина, витамина B 12) Для ионов (например, фосфатов, сульфатов) Биосинтетические ферменты Для сборки муреина (трансгликозидазы, карбоксипептидазы, транспептидазы) Для секреции и сборки жгутиковых субъединиц (например, шапиронины) Деградативные ферменты Фосфатазы Протеазы Детоксифицирующие ферменты β-лактамазы (пенициллиназа) Аминогликозид-фосфорилирующие ферменты
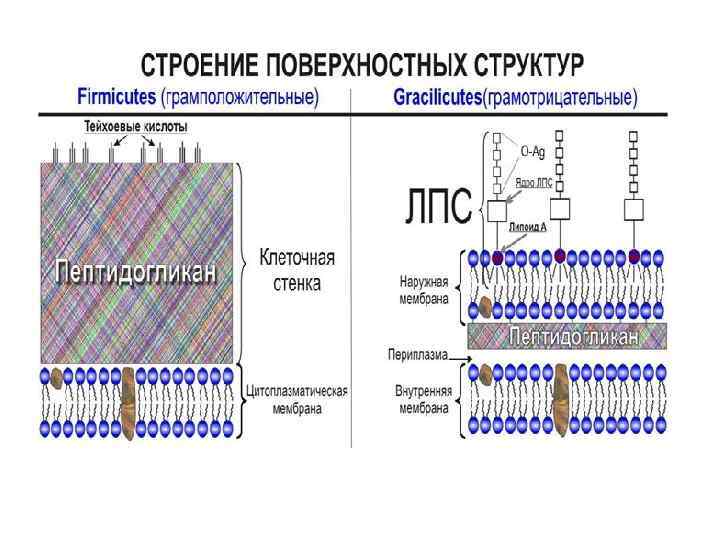
 Корреляция между реакцией окраски по Граму и свойствами клеточной стенки Свойства Gram-positive Gram-negative Толщина Толстая (20 -80 nm) Тонкая (10 nm) Количество слоев 1 2 Содержание пептидогликана (муреина) >50% 10 -20% Тейхоевые кислоты присутствуют Отсутствуют Содержание липидов и липопротеинов 0 -3% 58% Содержание протеинов 0 9% Содержание липополисахаридов 0 13% Чувствительность к Penicillin G Да Нет (1) Чувствительность к лизоциму да нет (2)
Корреляция между реакцией окраски по Граму и свойствами клеточной стенки Свойства Gram-positive Gram-negative Толщина Толстая (20 -80 nm) Тонкая (10 nm) Количество слоев 1 2 Содержание пептидогликана (муреина) >50% 10 -20% Тейхоевые кислоты присутствуют Отсутствуют Содержание липидов и липопротеинов 0 -3% 58% Содержание протеинов 0 9% Содержание липополисахаридов 0 13% Чувствительность к Penicillin G Да Нет (1) Чувствительность к лизоциму да нет (2)
 Микоплазмы
Микоплазмы
 Жидко-мозаичная модель биологической мембраны. В водной среде мембраные фосфолипиды спонтанно формируют жидкостный бислой. Мембранные белки, которые могут быть структурными или функциональными, могут постоянно или временно связываться с одной или другой стороной мембраны, или даже быть постоянно включены в бислой, в то время как другие белки проникают в бислой формируя транспортные каналы через мембрану.
Жидко-мозаичная модель биологической мембраны. В водной среде мембраные фосфолипиды спонтанно формируют жидкостный бислой. Мембранные белки, которые могут быть структурными или функциональными, могут постоянно или временно связываться с одной или другой стороной мембраны, или даже быть постоянно включены в бислой, в то время как другие белки проникают в бислой формируя транспортные каналы через мембрану.
 Жирная кислота (стеариновая) Обобщенная структура мембранных липидов. (Вврху). Фосфолипид в мембране") Жирная кислота (олеиновая) Жирная кислота (стеариновая) Обобщенная структура мембранных липидов. (Вврху). Фосфолипид в мембране Escherichia coli. R 1 и R 2 положения в глицероле замененные насыщеными или мононенасыщенными жирными кислотами. R 3 положение заменено фосфатидилэтаноламином, наиболее общий заместитель в этом положении у бактерий. (Внизу). Археальный мембранный липид. В отличие от бактериальных фософолипидов, которые являются, глицероловыми эфирами жирных кислот, липиды в мембранах Archaea - диэфиры глицерола и длинных разветвленных насыщенных углеводородных цепей, называемых изопреноиды или цепей составленых из повторяющихся C 5 субъединиц. Один из главных изопреноидов - C 20 молекула фитанол. R 3 положение глицерола может или может не быть замещено. Структура археальных мембранных липидов, как считают, является адаптацией к экстремальным условиям окружающей среды, например горячих и кислых условий, где Archaea преобладают в природе.
Жирная кислота (олеиновая) Жирная кислота (стеариновая) Обобщенная структура мембранных липидов. (Вврху). Фосфолипид в мембране Escherichia coli. R 1 и R 2 положения в глицероле замененные насыщеными или мононенасыщенными жирными кислотами. R 3 положение заменено фосфатидилэтаноламином, наиболее общий заместитель в этом положении у бактерий. (Внизу). Археальный мембранный липид. В отличие от бактериальных фософолипидов, которые являются, глицероловыми эфирами жирных кислот, липиды в мембранах Archaea - диэфиры глицерола и длинных разветвленных насыщенных углеводородных цепей, называемых изопреноиды или цепей составленых из повторяющихся C 5 субъединиц. Один из главных изопреноидов - C 20 молекула фитанол. R 3 положение глицерола может или может не быть замещено. Структура археальных мембранных липидов, как считают, является адаптацией к экстремальным условиям окружающей среды, например горячих и кислых условий, где Archaea преобладают в природе.
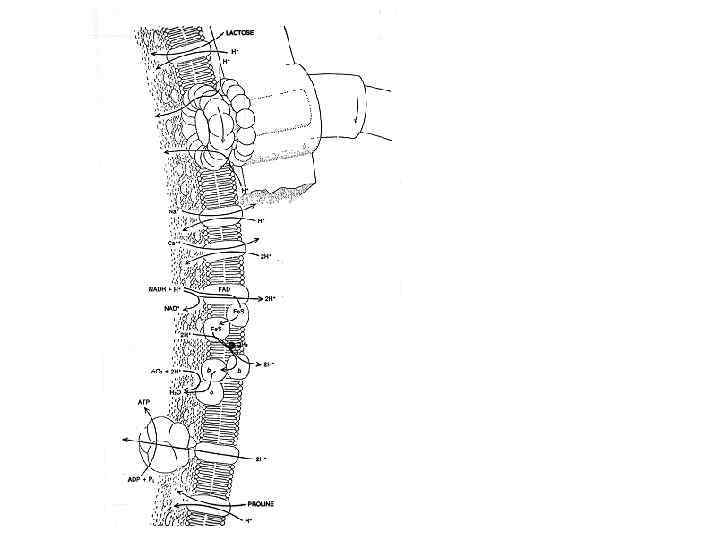
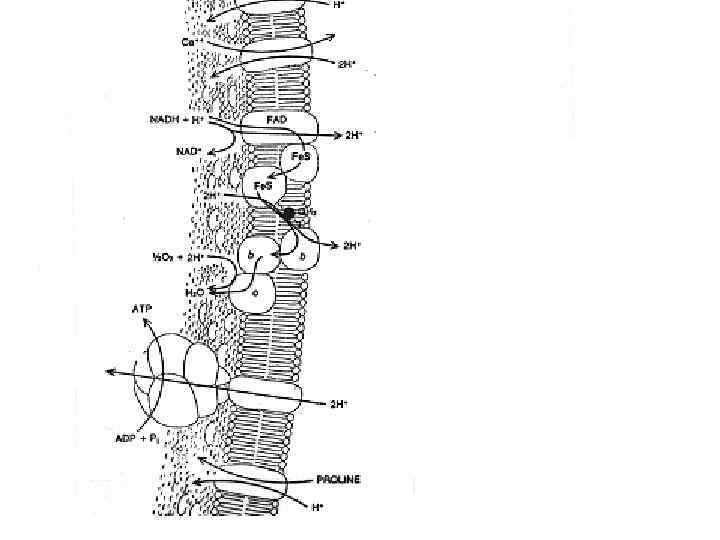
 Table 2. 3. Липидный состав клеточных мембран a Плазматическая мембрана Липид E. coli Эритроцит Шероховатый эндоплазматический ретикулум Другие митохондриальные мембраны Фосфатидилхолин 0 17 55 50 Фосфатидилсерин 0 6 3 2 80 16 16 23 Сфингомиелин 0 17 3 5 Гликолипиды 0 2 0 0 Холестерол 0 45 6 <5 Фосфатидилэтаноламин a Мембранный состав показан в моль% главных липидсоставляющих молекул.
Table 2. 3. Липидный состав клеточных мембран a Плазматическая мембрана Липид E. coli Эритроцит Шероховатый эндоплазматический ретикулум Другие митохондриальные мембраны Фосфатидилхолин 0 17 55 50 Фосфатидилсерин 0 6 3 2 80 16 16 23 Сфингомиелин 0 17 3 5 Гликолипиды 0 2 0 0 Холестерол 0 45 6 <5 Фосфатидилэтаноламин a Мембранный состав показан в моль% главных липидсоставляющих молекул.
 Таблица 6. Функции ЦПМ прокариот. 1. Осмотический барьер или барьер проницаемости 2. Локализация транспортных систем для специфических растворенных веществ (питательные вещества и ионы) 3. Генерирование энергии, с использованием дыхательных и фотосинтетических электронтранспортных систем, для создания протон движущей сылы, и трансмембранной, АТФ-синтезирующей АТФазы 4. Синтез мембранных липидов (включая ЛПС у грам-негативных бактерий) 5. Синтез муреина (пептидокликана клеточной стенки) 6. Сборка и секреция экстрацитоплазматических белков 7. Координация репликации ДНК и ее сегрегации с формированием септ и делением клетки 8. Хемотаксис (движение как таковое и сенсирующие функции) 9. Локализация специфических ферментативных систем
Таблица 6. Функции ЦПМ прокариот. 1. Осмотический барьер или барьер проницаемости 2. Локализация транспортных систем для специфических растворенных веществ (питательные вещества и ионы) 3. Генерирование энергии, с использованием дыхательных и фотосинтетических электронтранспортных систем, для создания протон движущей сылы, и трансмембранной, АТФ-синтезирующей АТФазы 4. Синтез мембранных липидов (включая ЛПС у грам-негативных бактерий) 5. Синтез муреина (пептидокликана клеточной стенки) 6. Сборка и секреция экстрацитоплазматических белков 7. Координация репликации ДНК и ее сегрегации с формированием септ и делением клетки 8. Хемотаксис (движение как таковое и сенсирующие функции) 9. Локализация специфических ферментативных систем
, малые полярные не") Figure 12. 15. Проницаемость бислойной фосфолипидной мембраны Газы, гидрофобные молекулы (бензол), малые полярные не заряженные молекулы могут диффундировать через бифосфолипидную мембрану. Большие полярные молекулы не могут.
Figure 12. 15. Проницаемость бислойной фосфолипидной мембраны Газы, гидрофобные молекулы (бензол), малые полярные не заряженные молекулы могут диффундировать через бифосфолипидную мембрану. Большие полярные молекулы не могут.
 Figure 15 -3. Диаграма схематически представляющая действие мембранных транспортных протеинов. Индикатором градиента явялется треугольник основание которого указывает на высокую концентрацию химических групп, электрического потенциала или их обоих. (a) Три основных типа транспортных протеинов. Насос использует энергию гидролиза АТФ для переноса специфических ионов или малых молекул против их электрохимического градиента. Каналы катализирующие перенос ионов (или воды) по их электрохимическому градиенту. Транспортеры, которые разделяются на три группы в зависимости от типа действия, специфических малых молекул или ионов. (b) Три группы транспортеров. Унипортеры, также продемострированы в части (a), транспортируют молекулы одного типа по концентрационному градиенту. Белки котранспортеры (симпортеры и антипортеры) катализируют движение одной молекулы против их концентрации, сопровождая движением одного или болшего количества ионов по их электрохимическому градиенту. Два типа котранспортеров различающихся по направлению движения транспортируемых молекул и котранспортируемых ионов.
Figure 15 -3. Диаграма схематически представляющая действие мембранных транспортных протеинов. Индикатором градиента явялется треугольник основание которого указывает на высокую концентрацию химических групп, электрического потенциала или их обоих. (a) Три основных типа транспортных протеинов. Насос использует энергию гидролиза АТФ для переноса специфических ионов или малых молекул против их электрохимического градиента. Каналы катализирующие перенос ионов (или воды) по их электрохимическому градиенту. Транспортеры, которые разделяются на три группы в зависимости от типа действия, специфических малых молекул или ионов. (b) Три группы транспортеров. Унипортеры, также продемострированы в части (a), транспортируют молекулы одного типа по концентрационному градиенту. Белки котранспортеры (симпортеры и антипортеры) катализируют движение одной молекулы против их концентрации, сопровождая движением одного или болшего количества ионов по их электрохимическому градиенту. Два типа котранспортеров различающихся по направлению движения транспортируемых молекул и котранспортируемых ионов.
 Figure 15 -15. Грам-негативные бактерии импортируют многие растворенные вещества при помощи ABC протеинов (АТФ-связывающие касетные пермеазы) которые используют белок связывающий растворенный субстрат, находящийся в периплазматическом пространстве. Здесь представлен импорт аминокислоты гистидина. После диффузии через порин наружной мембраны, гистидин связывается со специфическим гистидин-связывающим протеином, который при этом изменяет свою конформацию. Гистидин-белковый комплекс связывается с экзоплазматическим участком T субчастицы гистидиновой пермеазы расположенной в плазматической мембране. Гидролиз ATP A субчастицей протеина приводит к перемещению гистидина в цитозоль.
Figure 15 -15. Грам-негативные бактерии импортируют многие растворенные вещества при помощи ABC протеинов (АТФ-связывающие касетные пермеазы) которые используют белок связывающий растворенный субстрат, находящийся в периплазматическом пространстве. Здесь представлен импорт аминокислоты гистидина. После диффузии через порин наружной мембраны, гистидин связывается со специфическим гистидин-связывающим протеином, который при этом изменяет свою конформацию. Гистидин-белковый комплекс связывается с экзоплазматическим участком T субчастицы гистидиновой пермеазы расположенной в плазматической мембране. Гидролиз ATP A субчастицей протеина приводит к перемещению гистидина в цитозоль.
,") Мезосомы • Впячивания ЦПМ-мезосомы. Развиты у Г+. • 3 основных типа мезосом: ламеллярные (пластинчатые), везикулярные (имеющие форму пузырьков) и тубулярные (трубчатые). • По расположению в клетке различают мезосомы, образующиеся в зоне клеточного деления и формирования поперечной перегородки (септы), мезосомы, к которым прикреплен нуклеоид, и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ.
Мезосомы • Впячивания ЦПМ-мезосомы. Развиты у Г+. • 3 основных типа мезосом: ламеллярные (пластинчатые), везикулярные (имеющие форму пузырьков) и тубулярные (трубчатые). • По расположению в клетке различают мезосомы, образующиеся в зоне клеточного деления и формирования поперечной перегородки (септы), мезосомы, к которым прикреплен нуклеоид, и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ.
 Мезосомы • Мезосомы играют роль в репликации хромосомы и ее последующем расхождении по дочерним клеткам, участвуют в процессе инициации и формирования поперечной перегородки при клеточном делении. • разграничение внутриклеточного содержимого на обособленные отсеки, что создает более благоприятные условия для протекания определенных последовательностей ферментативных реакций.
Мезосомы • Мезосомы играют роль в репликации хромосомы и ее последующем расхождении по дочерним клеткам, участвуют в процессе инициации и формирования поперечной перегородки при клеточном делении. • разграничение внутриклеточного содержимого на обособленные отсеки, что создает более благоприятные условия для протекания определенных последовательностей ферментативных реакций.
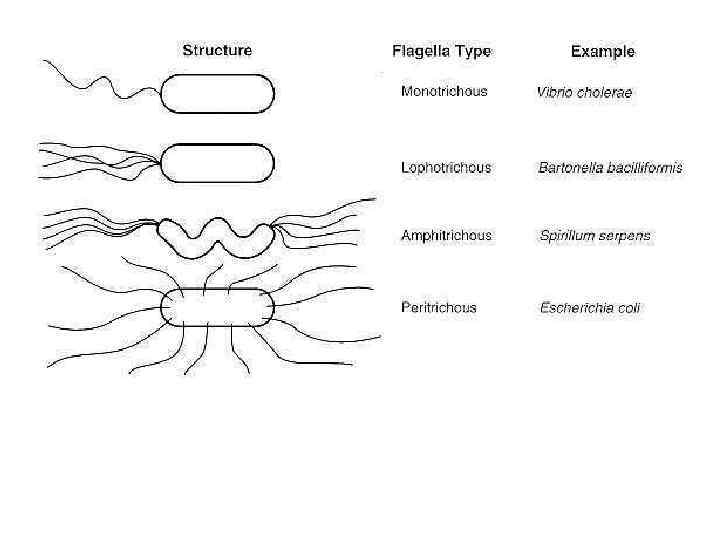
 и жгутики на поверхности бактериальных клеток. Слева: делящиеся Shigella с окружающими") Фимбрии (обычные pili) и жгутики на поверхности бактериальных клеток. Слева: делящиеся Shigella с окружающими их фимбриями. Структуры вероятно вовлечены в способность бактерий жестко фиксироваться на поверхности кишечника. Справа: деление пары клеток Salmonella, демонстрирующих их перитрихиально расположенные жгутики и их фимбрии. Фимбрии намного короче и слегка меньше в диаметре чем жгутики. И Shigella и Salmonella - энтеробактерии, которые вызывают различные типы кишечных диарей. Бактерии могут быть дифференцированы по обнаружению подвижности. Salmonella - подвижны; Shigella – не подвижны.
Фимбрии (обычные pili) и жгутики на поверхности бактериальных клеток. Слева: делящиеся Shigella с окружающими их фимбриями. Структуры вероятно вовлечены в способность бактерий жестко фиксироваться на поверхности кишечника. Справа: деление пары клеток Salmonella, демонстрирующих их перитрихиально расположенные жгутики и их фимбрии. Фимбрии намного короче и слегка меньше в диаметре чем жгутики. И Shigella и Salmonella - энтеробактерии, которые вызывают различные типы кишечных диарей. Бактерии могут быть дифференцированы по обнаружению подвижности. Salmonella - подвижны; Shigella – не подвижны.
 Figure 3. Salmonella enteritidis TEM about 10, 000 X. Salmonella is an enteric bacterium related to E. coli. The enterics are motile by means of peritriochous flagella.
Figure 3. Salmonella enteritidis TEM about 10, 000 X. Salmonella is an enteric bacterium related to E. coli. The enterics are motile by means of peritriochous flagella.
 Electron micrograph of negatively stained E coli showing wavy flagella") Figure 2 -3. (A) Electron micrograph of negatively stained E coli showing wavy flagella and numerous short, thinner, and more rigid hairlike structures, the pili. (B) The long sex pilus can be distinguished from the shorter common pili by mixing E coli cells with a male bacteriophage that binds specifically to sex pili.
Figure 2 -3. (A) Electron micrograph of negatively stained E coli showing wavy flagella and numerous short, thinner, and more rigid hairlike structures, the pili. (B) The long sex pilus can be distinguished from the shorter common pili by mixing E coli cells with a male bacteriophage that binds specifically to sex pili.
. Видны фибриллы") Электронная микрофотография ультратонкого среза делящейся пары стрептококков группы А (20, 000 X). Видны фибриллы на поверхности клеток, состоящие в основном из белка М. Также хорошо видна клеточная стенка к которой с одной стороны прикреплены фибриллы, а с другой стороны она контрастирована более электронно-плотной . цитоплазмой. Зона нуклеоида также отличается электроннопрозрачностью. Диаметр клеток составляет порядка одного микрона. Нуклеоид – область нуклеоплазмы которая не содержит рибосом, и в которой расположена бактериальная хромосома. Эта область не окружена мембраной.
Электронная микрофотография ультратонкого среза делящейся пары стрептококков группы А (20, 000 X). Видны фибриллы на поверхности клеток, состоящие в основном из белка М. Также хорошо видна клеточная стенка к которой с одной стороны прикреплены фибриллы, а с другой стороны она контрастирована более электронно-плотной . цитоплазмой. Зона нуклеоида также отличается электроннопрозрачностью. Диаметр клеток составляет порядка одного микрона. Нуклеоид – область нуклеоплазмы которая не содержит рибосом, и в которой расположена бактериальная хромосома. Эта область не окружена мембраной.
 Table 2. Некоторые свойства пилей. Распределение по поверхности Функции Escherichia coli (F или sex 1 -4 пили) перитрихиальное Коньюгация Escherichia coli (обычные 100 -200 пили фимбрии I типа) Перитрихиальное Прикрепление к поверхности клеток кишечного тракта Neisseria gonorrhoeae Перитрихиальное Прикрепление к поверхности клеток урогенитального тракта Бактериальные виды Streptococcus (фимбрии протеин) Типичное число на клетку 100 -200 pyogenes ? плюс M- Перитрихиальное Адгезия, резистентность к фагоцитозу, антигенная вариабельность Pseudomonas aeruginosa 10 -20 Полярное Поверхностная адгезия Sulfolobus (у архей) ? ? Прикрепление к частицам серы acidocaldarius
Table 2. Некоторые свойства пилей. Распределение по поверхности Функции Escherichia coli (F или sex 1 -4 пили) перитрихиальное Коньюгация Escherichia coli (обычные 100 -200 пили фимбрии I типа) Перитрихиальное Прикрепление к поверхности клеток кишечного тракта Neisseria gonorrhoeae Перитрихиальное Прикрепление к поверхности клеток урогенитального тракта Бактериальные виды Streptococcus (фимбрии протеин) Типичное число на клетку 100 -200 pyogenes ? плюс M- Перитрихиальное Адгезия, резистентность к фагоцитозу, антигенная вариабельность Pseudomonas aeruginosa 10 -20 Полярное Поверхностная адгезия Sulfolobus (у архей) ? ? Прикрепление к частицам серы acidocaldarius
 P-кольцо (ПГ) S-кольцо (ПГ или ППП) M-кольцо (ЦПМ) Figure 15 -61. Схема") L-кольцо (НМ) P-кольцо (ПГ) S-кольцо (ПГ или ППП) M-кольцо (ЦПМ) Figure 15 -61. Схема движения бактериального жгутикового мотора. Жгутик связан с подвижным крюком. Крюк соединен с серией белковых колец, которые пронизывают наружную и внутреннюю (ЦПМ) мембраны и вращаются со скоростью около 150 оборотов в секунду. Вращение, как считают, сообщается потоком протонов идущих через внешнее кольцо белков (the stator), которое кроме того содержит и белки отвечающие за изменение направления вращения. (Based on data from T. Kubori et al. , J. Mol. Biol. 226: 433 -446, 1992, and N. R. Francis et al. , Proc. Natl. Acad. Sci. USA 89: 6304 -6308, 1992. )
L-кольцо (НМ) P-кольцо (ПГ) S-кольцо (ПГ или ППП) M-кольцо (ЦПМ) Figure 15 -61. Схема движения бактериального жгутикового мотора. Жгутик связан с подвижным крюком. Крюк соединен с серией белковых колец, которые пронизывают наружную и внутреннюю (ЦПМ) мембраны и вращаются со скоростью около 150 оборотов в секунду. Вращение, как считают, сообщается потоком протонов идущих через внешнее кольцо белков (the stator), которое кроме того содержит и белки отвечающие за изменение направления вращения. (Based on data from T. Kubori et al. , J. Mol. Biol. 226: 433 -446, 1992, and N. R. Francis et al. , Proc. Natl. Acad. Sci. USA 89: 6304 -6308, 1992. )
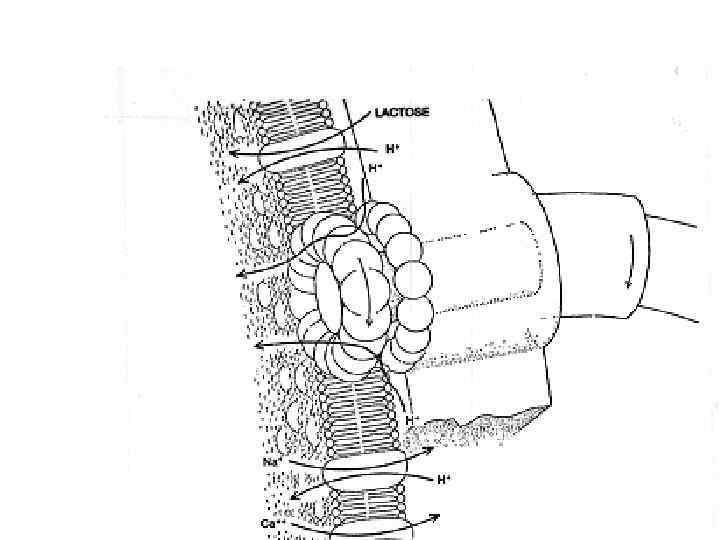
 Figure 15 -62. Расположение жгутиков у E. coli в процессе роения. Когда жгутики вращаются против часовой стрелки (A), они собираются и двигаются вместе на одном из полюсов клетки, что приводит к скольжению в одну сторону. Когда жгутики вращаются по часовой стрелке (B), они располагаются отдельно, что вызывает кувыркание клетки.
Figure 15 -62. Расположение жгутиков у E. coli в процессе роения. Когда жгутики вращаются против часовой стрелки (A), они собираются и двигаются вместе на одном из полюсов клетки, что приводит к скольжению в одну сторону. Когда жгутики вращаются по часовой стрелке (B), они располагаются отдельно, что вызывает кувыркание клетки.
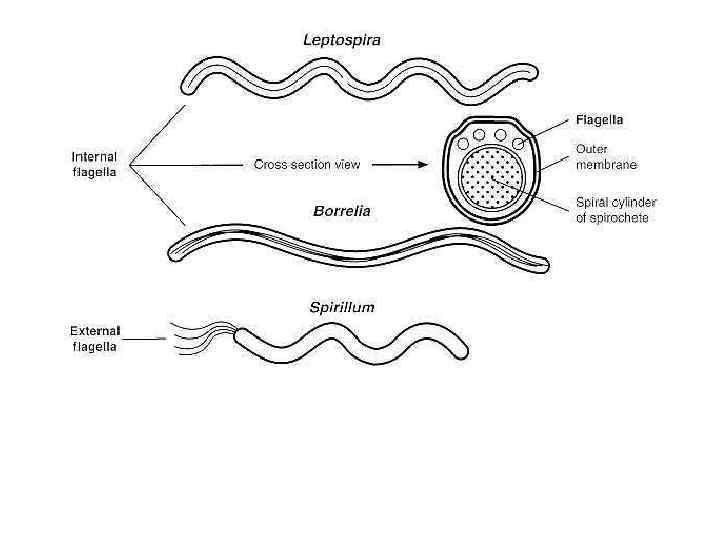
 Borrelia burgdorferi - спирохеты, которые размножаются в, клещах и распространяются среди мышей, и белохвостыми оленями через укусы инфицированных иксодовых клещей. Люди - случайные хозяева в этом цикле жизни, когда инфицируются клещами. Болезнь, которая возникает после инфекции, болезнь Lyme, названа по имени города в Штате Коннектикут, где болезнь была идентифицирована после того, как вспышка ювенильного ревматоидного артрита возникла у детей. Темнопольная микроскопия позволяет визуализацию, этих сильно извитых спиральных бактерии, которые не окрашиваются обычными бактериальными красителями.
Borrelia burgdorferi - спирохеты, которые размножаются в, клещах и распространяются среди мышей, и белохвостыми оленями через укусы инфицированных иксодовых клещей. Люди - случайные хозяева в этом цикле жизни, когда инфицируются клещами. Болезнь, которая возникает после инфекции, болезнь Lyme, названа по имени города в Штате Коннектикут, где болезнь была идентифицирована после того, как вспышка ювенильного ревматоидного артрита возникла у детей. Темнопольная микроскопия позволяет визуализацию, этих сильно извитых спиральных бактерии, которые не окрашиваются обычными бактериальными красителями.
 Figure 35 -3. Electron micrograph of Leptospira interrogans serovar icterohaemorrhagi ae. Bar equals 0. 5 mm.
Figure 35 -3. Electron micrograph of Leptospira interrogans serovar icterohaemorrhagi ae. Bar equals 0. 5 mm.
 А. Treponema denticola темнопольная микроскопия. Спирохеты ответственные за периодонтиты. Полоска 5 мкм. В. Деление клеток. Клетка удлинняется и делится на две дочерние. С. Обработка детергентом. После обработки периплазматические флагеллярные филаменты освобождаются и становятся видны цитоплазматичекие филаменты. Полоска 1 мкм.
А. Treponema denticola темнопольная микроскопия. Спирохеты ответственные за периодонтиты. Полоска 5 мкм. В. Деление клеток. Клетка удлинняется и делится на две дочерние. С. Обработка детергентом. После обработки периплазматические флагеллярные филаменты освобождаются и становятся видны цитоплазматичекие филаменты. Полоска 1 мкм.
. Видны фибриллы") Электронная микрофотография ультратонкого среза делящейся пары стрептококков группы А (20, 000 X). Видны фибриллы на поверхности клеток, состоящие в основном из белка М. Также хорошо видна клеточная стенка к которой с одной стороны прикреплены фибриллы, а с другой стороны она контрастирована более электронно-плотной . цитоплазмой. Зона нуклеоида также отличается электроннопрозрачностью. Диаметр клеток составляет порядка одного микрона. Нуклеоид – область нуклеоплазмы которая не содержит рибосом, и в которой расположена бактериальная хромосома. Эта область не окружена мембраной.
Электронная микрофотография ультратонкого среза делящейся пары стрептококков группы А (20, 000 X). Видны фибриллы на поверхности клеток, состоящие в основном из белка М. Также хорошо видна клеточная стенка к которой с одной стороны прикреплены фибриллы, а с другой стороны она контрастирована более электронно-плотной . цитоплазмой. Зона нуклеоида также отличается электроннопрозрачностью. Диаметр клеток составляет порядка одного микрона. Нуклеоид – область нуклеоплазмы которая не содержит рибосом, и в которой расположена бактериальная хромосома. Эта область не окружена мембраной.
, плотно распределенные рибосомальные частицы в") Figure 2 -2. Neisseria gonorrhoeae Электронно-прозрачный регион нуклеода (n), плотно распределенные рибосомальные частицы в цитоплазме, отсутствие внутриклеточных интрацелюлярных органелл.
Figure 2 -2. Neisseria gonorrhoeae Электронно-прозрачный регион нуклеода (n), плотно распределенные рибосомальные частицы в цитоплазме, отсутствие внутриклеточных интрацелюлярных органелл.
 Рибосомы • Рибосомы - место синтеза белка — рибонуклеопротеиновые частицы размером 15— 20 нм, количество непостоянно. • Константа седиментации 70 S (30 S и 50 S) • 30 S-частица содержит одну молекулу 16 S-р. РНК (косервативные элементы, основа геносистематики) и по одной молекуле белка 21 видa. • 50 S-субъединица состоит из двух молекул р. РНК (23 S и 5 S) и по одной копии 31 видов белка. • Рибосомальные белки-структурная функция. • Полисомы – содержат рибосомы, молекулы информационной РНК и т. РНК
Рибосомы • Рибосомы - место синтеза белка — рибонуклеопротеиновые частицы размером 15— 20 нм, количество непостоянно. • Константа седиментации 70 S (30 S и 50 S) • 30 S-частица содержит одну молекулу 16 S-р. РНК (косервативные элементы, основа геносистематики) и по одной молекуле белка 21 видa. • 50 S-субъединица состоит из двух молекул р. РНК (23 S и 5 S) и по одной копии 31 видов белка. • Рибосомальные белки-структурная функция. • Полисомы – содержат рибосомы, молекулы информационной РНК и т. РНК
 Бактериальный нуклеоид • Генетическая информация содержится в одной кольцевой двунитевой суперспирализованной молекуле ДНК (бактериальная хромосома). • ДНК прокариот построена так же, как и эукариот • Длина ДНК в 1000 раз превышает длину клетки. • Ядро бактерий не имеет ядерной оболочки, ядрышка, основных белков (гистонов) • Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. Нейтрализация зарядов осуществляется не гистонами, а полиаминами и ионами Mg 2+.
Бактериальный нуклеоид • Генетическая информация содержится в одной кольцевой двунитевой суперспирализованной молекуле ДНК (бактериальная хромосома). • ДНК прокариот построена так же, как и эукариот • Длина ДНК в 1000 раз превышает длину клетки. • Ядро бактерий не имеет ядерной оболочки, ядрышка, основных белков (гистонов) • Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. Нейтрализация зарядов осуществляется не гистонами, а полиаминами и ионами Mg 2+.
. Шкала 0, 1") А. Ультратонкий срез клеток Klebsiella pneumoniae. Видна фибриллярная ультраструктура нуклеоида (Н). Шкала 0, 1 мкм. В. Изолированный нуклеоид из E. coli, полученный при лизисе клеток путем осмотического шока. ДНК менее плотная и частично деспирализована из-за разрушения комплексов белок-нуклиновые кислоты. Видны петли ДНК. Шкала 1
А. Ультратонкий срез клеток Klebsiella pneumoniae. Видна фибриллярная ультраструктура нуклеоида (Н). Шкала 0, 1 мкм. В. Изолированный нуклеоид из E. coli, полученный при лизисе клеток путем осмотического шока. ДНК менее плотная и частично деспирализована из-за разрушения комплексов белок-нуклиновые кислоты. Видны петли ДНК. Шкала 1
 Контрастированный металлом препарат клеток E. coli, разрушенных при использовании мягких условий лизиса, т. е. не осмотического шока. Фрагменты нитей хромосомы выходят в среду вместе со связанными белками и РНК в виде нитей бус. ДНК-связывающие белки и РНК могут играть у прокариот важную роль в конденсации ДНК и в регуляции процессов с участием ДНК. При помощи ДНК-связывающих белков нить ДНК длинной не менее 1, 2 мм упаковывается в клетке в структуру длинной несколько микронов.
Контрастированный металлом препарат клеток E. coli, разрушенных при использовании мягких условий лизиса, т. е. не осмотического шока. Фрагменты нитей хромосомы выходят в среду вместе со связанными белками и РНК в виде нитей бус. ДНК-связывающие белки и РНК могут играть у прокариот важную роль в конденсации ДНК и в регуляции процессов с участием ДНК. При помощи ДНК-связывающих белков нить ДНК длинной не менее 1, 2 мм упаковывается в клетке в структуру длинной несколько микронов.
 Хромосома Плазмида Генетический материал бактерий представлен двунитевой молекулой ДНК замкнутой в кольцо. Кроме хромосом, генетическая информация содержится у прокариот в более мелких элементах – плазмидах. Плазмиды – это независимые элементы, содержащие дополнительные гены (необходимые лишь в специфических условиях). Подобно хромосомам, плазмиды реплицируются автономно. Размер хромосом у бактерий обычно составляет несколько миллионов пар нуклеотидов, редко несколько сотен тысяч пар нуклеотидов. Размер плазмид обычно составляет тысячи или десятки тысяч
Хромосома Плазмида Генетический материал бактерий представлен двунитевой молекулой ДНК замкнутой в кольцо. Кроме хромосом, генетическая информация содержится у прокариот в более мелких элементах – плазмидах. Плазмиды – это независимые элементы, содержащие дополнительные гены (необходимые лишь в специфических условиях). Подобно хромосомам, плазмиды реплицируются автономно. Размер хромосом у бактерий обычно составляет несколько миллионов пар нуклеотидов, редко несколько сотен тысяч пар нуклеотидов. Размер плазмид обычно составляет тысячи или десятки тысяч
 Двойная спираль В-формы ДНК. Две антипараллельные цепи ДНК закручены в правостороннюю двойную спираль, имеющую большую и малую бороздки, в которых ДНК-связывающие белки узнают последовательности нуклеотидов
Двойная спираль В-формы ДНК. Две антипараллельные цепи ДНК закручены в правостороннюю двойную спираль, имеющую большую и малую бороздки, в которых ДНК-связывающие белки узнают последовательности нуклеотидов
 ДНК состоит из сахарофосфатного остова, представляющего собой последовательность из чередующихся остатков сахара и фосфата и связанных при помощи N- гликозидных связей с дезоксирибозой
ДНК состоит из сахарофосфатного остова, представляющего собой последовательность из чередующихся остатков сахара и фосфата и связанных при помощи N- гликозидных связей с дезоксирибозой
 образует две водородные") Пары пуриновых и пиримидиновых оснований в составе ДНК. Пара аденинтимин (А-Т) образует две водородные связи, гуанин-цитозин (G-C) – три. Следует обратить внимание на нумерацию атомов в остатках оснований (1 -9)
Пары пуриновых и пиримидиновых оснований в составе ДНК. Пара аденинтимин (А-Т) образует две водородные связи, гуанин-цитозин (G-C) – три. Следует обратить внимание на нумерацию атомов в остатках оснований (1 -9)
 В полинуклеотидной цепи ДНК дезоксинуклеотиды соединены 3’-5’ фосфодиэфирными связями, образующимися между кольцами дезоксирибозы
В полинуклеотидной цепи ДНК дезоксинуклеотиды соединены 3’-5’ фосфодиэфирными связями, образующимися между кольцами дезоксирибозы
 В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке (слева направо) представлены A, B и Z формы
В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке (слева направо) представлены A, B и Z формы
 Длина геномов не зависит от сложности организма. Сравнительные размеры геномов в разных группах организмов Объект размер генома, пн Микоплазмы 104 -106 Эубактерии (E. coli) 105 -107 Грибы (2 -5)x 107 Водоросли (5 -7)x 10 Черви ~108 Моллюски 5 x 108 -5 x 109 Насекомые 108 -5 x 109 Ракообразные ~109 Иглокожие 2 x 108 -2 x 109 Рыбы 3 x 108 -1010 Амфибии 7 x 108 -7 x 1010 Рептилии (2 -3)109 Птицы 109 Млекопитающие 3 x 109 Цветковые растения 2 x 108 -1011 .
Длина геномов не зависит от сложности организма. Сравнительные размеры геномов в разных группах организмов Объект размер генома, пн Микоплазмы 104 -106 Эубактерии (E. coli) 105 -107 Грибы (2 -5)x 107 Водоросли (5 -7)x 10 Черви ~108 Моллюски 5 x 108 -5 x 109 Насекомые 108 -5 x 109 Ракообразные ~109 Иглокожие 2 x 108 -2 x 109 Рыбы 3 x 108 -1010 Амфибии 7 x 108 -7 x 1010 Рептилии (2 -3)109 Птицы 109 Млекопитающие 3 x 109 Цветковые растения 2 x 108 -1011 .
![Размеры геномов и количество генов [Alberts, 2002] Вид Число генов Длина генома, пн Eubacteria](https://present5.com/presentation/3/-42473852_285389902.pdf-img/-42473852_285389902.pdf-58.jpg "Размеры геномов и количество генов [Alberts, 2002] Вид Число генов Длина генома, пн Eubacteria") Размеры геномов и количество генов [Alberts, 2002] Вид Число генов Длина генома, пн Eubacteria Mycoplasma genitalium 477 580. 070 E. coli 4280 4. 639. 221 Helicobacter pylori 1590 1667 тыс Bacillus subtilis 4099 4214 тыс Micobacterium tuberculosis 4402 4447 тыс Treponema pallidum 1041 1138 тыс Rickettsia prowazekii 834 1111 тыс Eucaryotes Saccharomyces cerevisiae ~6300 12. 069 тыс Drosophila melanogaster ~14000 137. 000 тыс Homo sapiens ~30000 3. 200. 000 тыс
Размеры геномов и количество генов [Alberts, 2002] Вид Число генов Длина генома, пн Eubacteria Mycoplasma genitalium 477 580. 070 E. coli 4280 4. 639. 221 Helicobacter pylori 1590 1667 тыс Bacillus subtilis 4099 4214 тыс Micobacterium tuberculosis 4402 4447 тыс Treponema pallidum 1041 1138 тыс Rickettsia prowazekii 834 1111 тыс Eucaryotes Saccharomyces cerevisiae ~6300 12. 069 тыс Drosophila melanogaster ~14000 137. 000 тыс Homo sapiens ~30000 3. 200. 000 тыс
 ДНК. Слева направо: линейная, релаксированная, кольцевая сверхспирализованная и частично развернутая") Топоизомеры плазмидной (RFS 1010) ДНК. Слева направо: линейная, релаксированная, кольцевая сверхспирализованная и частично развернутая (показана стрелкой) сверхспирализованная ДНК
Топоизомеры плазмидной (RFS 1010) ДНК. Слева направо: линейная, релаксированная, кольцевая сверхспирализованная и частично развернутая (показана стрелкой) сверхспирализованная ДНК
 Для бактерий характерна оперонная модель экспрессии генов. Оперон представляет собой транскрипционную единицу в составе хромосомы, содержащую один (моноцистронная единица) или несколько (полицистронная единица) генов, старт-сигнал и стоп-сигнал для РНКполимеразы. В полицистронном опероне гены подвергаются сопряженной регуляции. Физическое сцепление этих генов обеспечивает транскрипцию их в виде одной молекулы РНК, количество копий которой изменяется под действием вне- или внутриоперонных регуляторных элементов.
Для бактерий характерна оперонная модель экспрессии генов. Оперон представляет собой транскрипционную единицу в составе хромосомы, содержащую один (моноцистронная единица) или несколько (полицистронная единица) генов, старт-сигнал и стоп-сигнал для РНКполимеразы. В полицистронном опероне гены подвергаются сопряженной регуляции. Физическое сцепление этих генов обеспечивает транскрипцию их в виде одной молекулы РНК, количество копий которой изменяется под действием вне- или внутриоперонных регуляторных элементов.
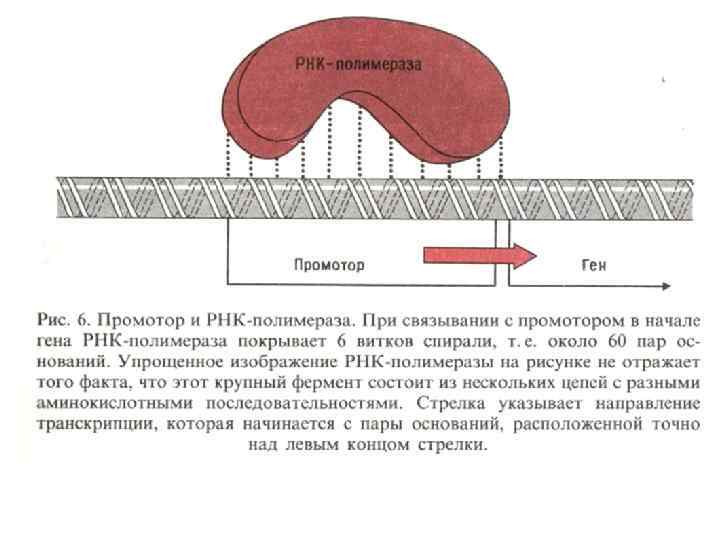
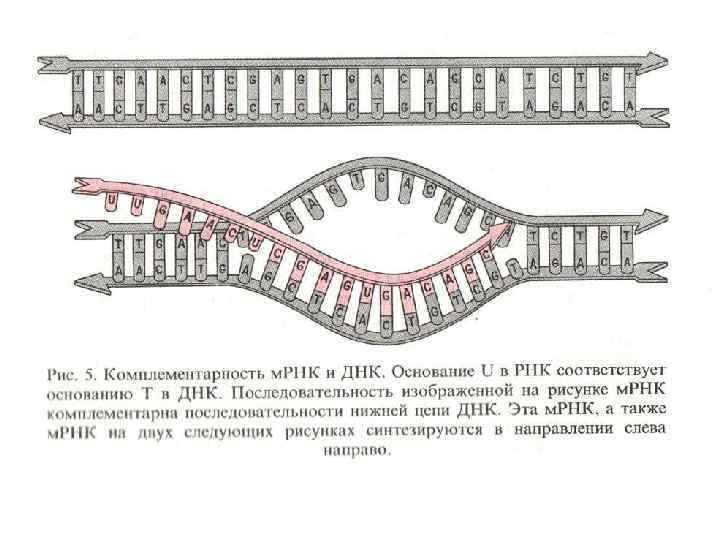
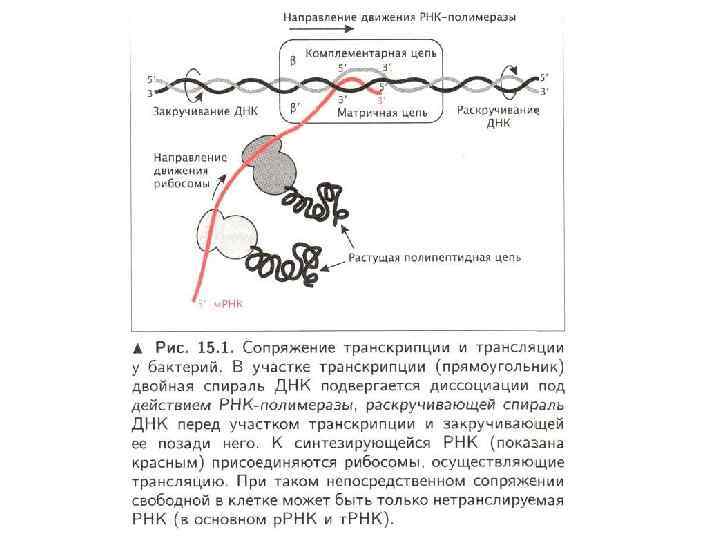
 Растущие белки и. РНК-полимераза Полирибосомальный комплекс Старт трансляции Связь транскрипции и трансляции у бактерий. Сразу после начала траскрипции рибосомы связываются с и. РНК на ее 5’ конце. Таким образом, трансляция стартует еще до завершения процесса траскрипции. Многочисленные рибосомы связываются с матричной РНК формируя полисомы
Растущие белки и. РНК-полимераза Полирибосомальный комплекс Старт трансляции Связь транскрипции и трансляции у бактерий. Сразу после начала траскрипции рибосомы связываются с и. РНК на ее 5’ конце. Таким образом, трансляция стартует еще до завершения процесса траскрипции. Многочисленные рибосомы связываются с матричной РНК формируя полисомы
 Активированный участок генома E. coli – оперон с предположительным сайтом транскрипции. Видны временно существующие комплексы, содержащие ДНК-зависимую РНК полимеразу и новообразованную м. РНК с полисомами, состаящими из рибосом, осуществляющими трасляцию.
Активированный участок генома E. coli – оперон с предположительным сайтом транскрипции. Видны временно существующие комплексы, содержащие ДНК-зависимую РНК полимеразу и новообразованную м. РНК с полисомами, состаящими из рибосом, осуществляющими трасляцию.
 Точка начала репликации Вилка Сайт завершения репликации Репликация ДНК. Цепи ДНК разделяются и каждая из них служит матрицей для синтеза комплементарной цепи ДНК. Каждая из дочерних молекул наполовину состоит из родительского материала. Этот механизм назван полуконсервативной репликацией.
Точка начала репликации Вилка Сайт завершения репликации Репликация ДНК. Цепи ДНК разделяются и каждая из них служит матрицей для синтеза комплементарной цепи ДНК. Каждая из дочерних молекул наполовину состоит из родительского материала. Этот механизм назван полуконсервативной репликацией.
 Ион Функции K+ Поддержание ионной силы, кофактор некоторых ферментов NH 4+ Основная форма неорганического N для ассимиляции Ca++ Кофактор некоторых ферментов Fe++ Присутствует в цитохромах и других металлоферментах Mg++ Кофактор многих ферментов; стабилизация наружной мембраны Грам-негативных бактерий Присутствует в некоторых металлоферментах Mn++ Co++ Cu++ Следы элемента в составе B 12 и его коферментных производных и найдены в некоторых металлоферментах Следы элемента присутствуют в некоторых металлоферментах Mo++ Следы элемента присутствуют в некоторых металлоферментах Ni++ Следы элемента присутствуют в некоторых металлоферментах Zn++ Следы элемента присутствуют в некоторых металлоферментах SO 4 -- Основная форма неорганической серы для ассимиляции PO 4 --- Основная форма P для ассимиляции и участи во многих метаболических реакциях
Ион Функции K+ Поддержание ионной силы, кофактор некоторых ферментов NH 4+ Основная форма неорганического N для ассимиляции Ca++ Кофактор некоторых ферментов Fe++ Присутствует в цитохромах и других металлоферментах Mg++ Кофактор многих ферментов; стабилизация наружной мембраны Грам-негативных бактерий Присутствует в некоторых металлоферментах Mn++ Co++ Cu++ Следы элемента в составе B 12 и его коферментных производных и найдены в некоторых металлоферментах Следы элемента присутствуют в некоторых металлоферментах Mo++ Следы элемента присутствуют в некоторых металлоферментах Ni++ Следы элемента присутствуют в некоторых металлоферментах Zn++ Следы элемента присутствуют в некоторых металлоферментах SO 4 -- Основная форма неорганической серы для ассимиляции PO 4 --- Основная форма P для ассимиляции и участи во многих метаболических реакциях
 Некоторые включения в бактериальные клетки. Включения Обнаружены Состав Гликоген Многие бактерии Полиглюкоза напр. , E. coli Запас углеводов и источник энергии Полигидрокси маслянная кислота Полифосфаты (зерна волютина) Многи бактерии Полимер ГМК напр. , Pseudomonas Запас углеводов и источник энергии Многи бактерии напр, , Corynebacterium Функции Линейные или циклические полимеры PO 4 Запас фосфатов; возможно выскоэнергетических связей Гранулы серы Фототрофные урпурные п и зеленые серные бактерии Пузырьки газа Водные бактерии в частности цианобактерии Белковые пузырьки с газом Околоспоровые кристаллы (род Bacillus) Белки Магнитосомы Некоторые бактерии Карбоксисомы Многие автотрофные автотрофной CO 2 Сайт CO 2 фиксации бактерии фиксации водные Элементарная сера Запас электронов (источник восстановителей) у фототрофов; запас источника энергии у литотрофов Плавучесть в вертикальном направлении Неизвестно, но токсичны для некоторых насекомых магнетит (оксид Ориентация и миграция железа) Fe 3 O 4 геомагнитных линий Ферменты запас для вдоль
Некоторые включения в бактериальные клетки. Включения Обнаружены Состав Гликоген Многие бактерии Полиглюкоза напр. , E. coli Запас углеводов и источник энергии Полигидрокси маслянная кислота Полифосфаты (зерна волютина) Многи бактерии Полимер ГМК напр. , Pseudomonas Запас углеводов и источник энергии Многи бактерии напр, , Corynebacterium Функции Линейные или циклические полимеры PO 4 Запас фосфатов; возможно выскоэнергетических связей Гранулы серы Фототрофные урпурные п и зеленые серные бактерии Пузырьки газа Водные бактерии в частности цианобактерии Белковые пузырьки с газом Околоспоровые кристаллы (род Bacillus) Белки Магнитосомы Некоторые бактерии Карбоксисомы Многие автотрофные автотрофной CO 2 Сайт CO 2 фиксации бактерии фиксации водные Элементарная сера Запас электронов (источник восстановителей) у фототрофов; запас источника энергии у литотрофов Плавучесть в вертикальном направлении Неизвестно, но токсичны для некоторых насекомых магнетит (оксид Ориентация и миграция железа) Fe 3 O 4 геомагнитных линий Ферменты запас для вдоль
, кортекс (OCL)") Figure 2 -11. Спора Bacillus megaterium spore показаны толстые споровые оболочки (SC), кортекс (OCL) клеточная стенка (GCW), ЦПМ (PM), нуклеоид (n)e. (Courtesy of John H Freer, University of Glasgow, Scotland. )
Figure 2 -11. Спора Bacillus megaterium spore показаны толстые споровые оболочки (SC), кортекс (OCL) клеточная стенка (GCW), ЦПМ (PM), нуклеоид (n)e. (Courtesy of John H Freer, University of Glasgow, Scotland. )
 Свойства Поверхностные оболочки Вегетативные клетки. Типичные для Гр+ бактерий Эндоспоры Толстая споровая оболочка, покровы, кортекс, пептидогликан Вид при микроскопии Не преломляет свет Преломляет свет Дипиколинат кальция Отсутствует Присутствует Цитоплазматическая активность воды Высокая Очень низкая Энзиматическая активность Присутствует Отсутствует Макромолекулярный синтез Присутствует Резистентность к to. C низкая Высокая Резистентность к кислотам и другим химическим агентам низкая Высокая Радиорезистентность низкая Высокая Чувствительность к лизоциму Чувствительны Резистентны Чувствительность к окраске Чувствительны Резистентны Отсутствует
Свойства Поверхностные оболочки Вегетативные клетки. Типичные для Гр+ бактерий Эндоспоры Толстая споровая оболочка, покровы, кортекс, пептидогликан Вид при микроскопии Не преломляет свет Преломляет свет Дипиколинат кальция Отсутствует Присутствует Цитоплазматическая активность воды Высокая Очень низкая Энзиматическая активность Присутствует Отсутствует Макромолекулярный синтез Присутствует Резистентность к to. C низкая Высокая Резистентность к кислотам и другим химическим агентам низкая Высокая Радиорезистентность низкая Высокая Чувствительность к лизоциму Чувствительны Резистентны Чувствительность к окраске Чувствительны Резистентны Отсутствует
 Электронная микрография бактериальной эндоспоры.
Электронная микрография бактериальной эндоспоры.

 Figure 14. 13 Cтадии споруляции B. subtilis. Левая панель показывает электронную микрофотографию стадии споруляции (EM), правая показывает, рисованную картинку, расположение генетического материала и время и сайт действия основных регуляторных протеинов которые управляют экспрессией генов споруляции. (A) Вегетативные клетки. ( с B до E) Спорангий входит в споруляцию (этап 0) (B), с полярным делением (также называемым полярное септирование) (этап II) (C), и поглощенение (этап III) (D), и формирование кортекса и оболочек (этапы V to VI) (E). (F) Свободная спора.
Figure 14. 13 Cтадии споруляции B. subtilis. Левая панель показывает электронную микрофотографию стадии споруляции (EM), правая показывает, рисованную картинку, расположение генетического материала и время и сайт действия основных регуляторных протеинов которые управляют экспрессией генов споруляции. (A) Вегетативные клетки. ( с B до E) Спорангий входит в споруляцию (этап 0) (B), с полярным делением (также называемым полярное септирование) (этап II) (C), и поглощенение (этап III) (D), и формирование кортекса и оболочек (этапы V to VI) (E). (F) Свободная спора.
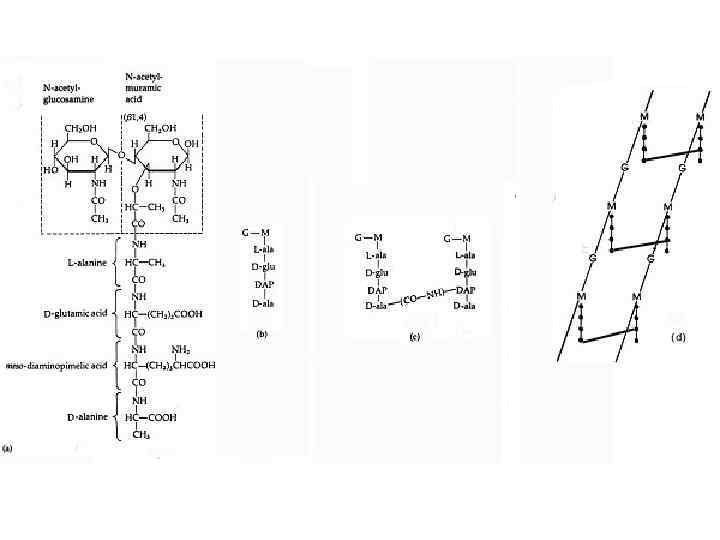
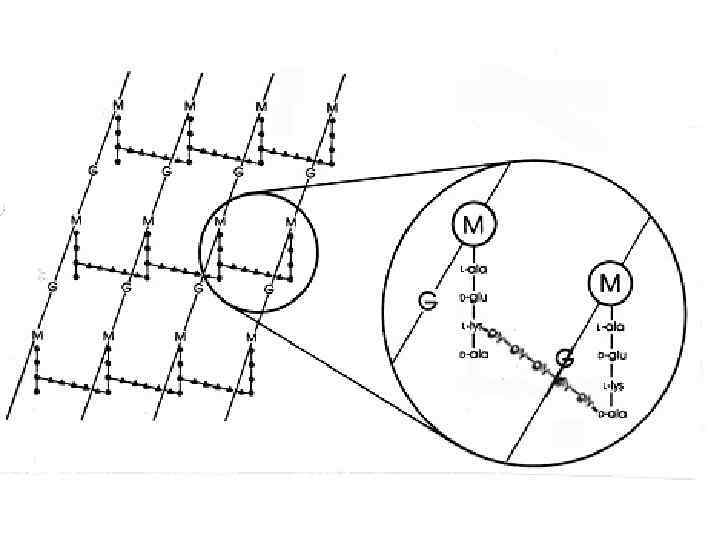
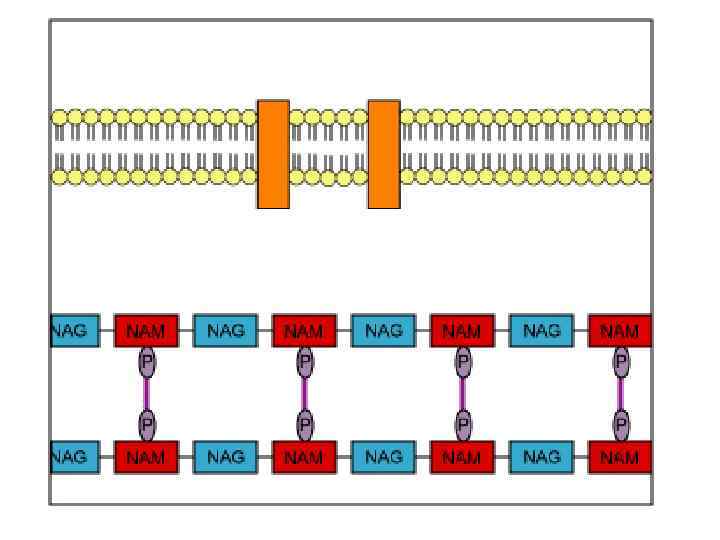
 FIG. 1. Cтруктура пептидогликана спор B. subtilis. Базовая структура пептидогликана спор была определена Warth и Strominger. Полисахаридная цепь состоит из чередующихся остатков N-acetylglucosamine (NAG) и N-acetyl-muramic acid (NAM). К NAM может быть присоединена боковая цепь состоящая из эфира L-Ala (в 25% случаях) или тетерапептида LAla-g-D-Glu-Dpm-D-Ala. Более 50% остатаков мурамовой кислоты не имеют присоединенной боковой пептидной цепи и могут быть конвертированы в форму лактама мурамовой кислоты (MAL). Полисахаридныеи цепи могут быть перекрестно соединены посредством связей между тетрапептидными цепочками или трипептидными цепочками. Только 11% диаминопимелиновых кислот вовлечены в образование перекрестных связей.
FIG. 1. Cтруктура пептидогликана спор B. subtilis. Базовая структура пептидогликана спор была определена Warth и Strominger. Полисахаридная цепь состоит из чередующихся остатков N-acetylglucosamine (NAG) и N-acetyl-muramic acid (NAM). К NAM может быть присоединена боковая цепь состоящая из эфира L-Ala (в 25% случаях) или тетерапептида LAla-g-D-Glu-Dpm-D-Ala. Более 50% остатаков мурамовой кислоты не имеют присоединенной боковой пептидной цепи и могут быть конвертированы в форму лактама мурамовой кислоты (MAL). Полисахаридныеи цепи могут быть перекрестно соединены посредством связей между тетрапептидными цепочками или трипептидными цепочками. Только 11% диаминопимелиновых кислот вовлечены в образование перекрестных связей.
 Все клеточные формы жизни на основании сравнительного анализа нуклеотидной последовательности их генов кодирующих 16 S р. РНК и других фундаментальных различий (аппараты трансляции и транскрипции, организация генетического материала и другое) разделены на 3 домена Вacteria Archaea Eucaria (Eubacteria - истинные бактерии) Группированы на 27 отделов, в т. ч. : Proteobacteria Firmicutes Actinobacteria Bacteroidetes Spirochaetes Chlamydiae Tenericutes Cyanobacteria (сине-зеленые водоросли)
Все клеточные формы жизни на основании сравнительного анализа нуклеотидной последовательности их генов кодирующих 16 S р. РНК и других фундаментальных различий (аппараты трансляции и транскрипции, организация генетического материала и другое) разделены на 3 домена Вacteria Archaea Eucaria (Eubacteria - истинные бактерии) Группированы на 27 отделов, в т. ч. : Proteobacteria Firmicutes Actinobacteria Bacteroidetes Spirochaetes Chlamydiae Tenericutes Cyanobacteria (сине-зеленые водоросли)
 28 классов Отделы группированы на , например: В отдел Firmicutes (Грам-позитивные бактерии с низким % содержанием G+C пар) входят, классы: Bacilli, подразделяющийся на порядки: Bacillales Lactobacillales Clostridia, подразделяющийся на порядки: Clostridiales Halanaerobiales Thermoanaerobacteriales Кроме того, к фирмикутам относятся рода Staphylococcus, Enterococcus и другие. В отдел Chlamydiae входит, класс: Chlamydiae В отдел Proteobacteria входят, классы: Alphaproteobacteria (порядок - Rickettsiales; семейство - Rickettsiaceae; триба - Rickettsieae; род - Rickettsia; группа тифов; Rickettsia typhi) Betaproteobacteria (порядок - Burkholderiales; семейство - Alcaligenaceae; род -Bordetella; вид - Bordetella pertussis) Gammaproteobacteria (порядок - Enterobacteriales; семейство - Enterobacteriaceae; род - Escherichia, вид Escherichia coli) Epsilonproteobacteria (порядок - Campylobacterales, семейство - Helicobacteraceae, род – Helicobacter, вид - Helicobacter pylori ) В отдел Actinobacteria (Грам-позитивные бактерии с высоким содержанием G+C пар) входят рода: Bifidobacterium, Mycobacterium, Corynebacterium В отдел Tenericutes (Бактерии без клеточной стенки) входит класс: Mollicutes, подразделяющийся на порядки: Acholeplasmatales Anaeroplasmatales Entomoplasmatales Mycoplasmatales
28 классов Отделы группированы на , например: В отдел Firmicutes (Грам-позитивные бактерии с низким % содержанием G+C пар) входят, классы: Bacilli, подразделяющийся на порядки: Bacillales Lactobacillales Clostridia, подразделяющийся на порядки: Clostridiales Halanaerobiales Thermoanaerobacteriales Кроме того, к фирмикутам относятся рода Staphylococcus, Enterococcus и другие. В отдел Chlamydiae входит, класс: Chlamydiae В отдел Proteobacteria входят, классы: Alphaproteobacteria (порядок - Rickettsiales; семейство - Rickettsiaceae; триба - Rickettsieae; род - Rickettsia; группа тифов; Rickettsia typhi) Betaproteobacteria (порядок - Burkholderiales; семейство - Alcaligenaceae; род -Bordetella; вид - Bordetella pertussis) Gammaproteobacteria (порядок - Enterobacteriales; семейство - Enterobacteriaceae; род - Escherichia, вид Escherichia coli) Epsilonproteobacteria (порядок - Campylobacterales, семейство - Helicobacteraceae, род – Helicobacter, вид - Helicobacter pylori ) В отдел Actinobacteria (Грам-позитивные бактерии с высоким содержанием G+C пар) входят рода: Bifidobacterium, Mycobacterium, Corynebacterium В отдел Tenericutes (Бактерии без клеточной стенки) входит класс: Mollicutes, подразделяющийся на порядки: Acholeplasmatales Anaeroplasmatales Entomoplasmatales Mycoplasmatales
 Высшие таксоны Домен Вacteria Отдел Proteobacteria Класс Gammaproteobacteria Малые таксоны Порядок Enterobacteriales Семейство Enterobacteriaceae Род Escherichia Вид Escherichia coli
Высшие таксоны Домен Вacteria Отдел Proteobacteria Класс Gammaproteobacteria Малые таксоны Порядок Enterobacteriales Семейство Enterobacteriaceae Род Escherichia Вид Escherichia coli