Земноводные_пресмыкающиеся.pptx
- Количество слайдов: 26



Челюсти в классах костных и хрящевых рыб эволюционно образовались из третьей пары жаберных дуг (о чём свидетельствуют рудименты первых двух пар дуг у акул — так называемые губные хрящи). У костистых рыб челюсти несут основные группы зубов на переднечелюстной (premaxilla) и верхнечелюстной костях (maxilla) (верхняя челюсть), на dentale и articulare (нижняя челюсть). Несколько специализированных групп костей формируют дно ротовой полости и соединяют челюсти с другими элементами черепа. Наиболее рострально (впереди) расположена гиоидная дуга, которая играет важную роль при изменении объёма ротовой полости. За ней идут жаберные дуги, которые несут жаберные дыхательные структуры, и наиболее каудально расположены так называемые глоточные челюсти, которые также могут нести зубы. Во время питания мышцы, которые опускают комплекс нижней челюсти, смещают этот комплекс таким образом, что челюсти выдвигаются вперед. При этом в ротовой полости создаётся всасывающая сила за счёт опускания дна рта. Жаберные крышки при этом закрывают жабры. Такая комбинация движений приводит к всасыванию воды и втягиванию пищи в рот. Позвоночник рыб состоит из отдельных, не сращённых позвонков. Позвонки рыб амфицельные (то есть обе их торцевые поверхности вогнутые), между позвонками находится хрящевая прослойка; невральные дуги, расположенные сверху над телами позвонков, формируют позвоночный канал, защищающий спинной мозг. Позвоночник делится на два отдела — туловищный и хвостовой. От позвонков туловища в стороны отходят рёберные отростки, к которым крепятся рёбра. В хвостовом отделе позвоночника боковых отростков нет, зато кроме невральной дуги имеется сосудистая (гемальная) дуга, которая прикрепляется к позвонку снизу и защищает проходящий в ней большой кровеносный сосуд — брюшную аорту. От невральных и гемальных дуг вертикально вверх и вниз отходят заострённые остистые отростки.

Мурены обладают двумя парами челюстей. Вторая пара, называемая глоточной, или фарингеальной, находится позади черепа. Хотя внутренние челюсти не выходят за пределы внешних, они помогают более крепко ухватить добычу

Парные плавники, по сути представляют собой парные конечности, опирающиеся на соответствующие им внутренние скелетные элементы — пояса плавников. У подавляющего большинства рыб представлены двумя парами плавников: грудным плавником (лат. pinna pectoralis, обозначается латинской буквой P) и брюшным плавником (лат. pinna ventralis, обозначается заглавной латинской буквой V).

Глаза рыб по своему строению очень схожи с глазами других позвоночных. Важное отличие глаза рыб от глаза млекопитающих заключается в том, что для аккомодации рыбы не изменяют кривизну хрусталика, а приближают его к сетчатке или отдаляют от неё. Перемещение воды по поверхности рыбы ощущается структурами, которые называются нейромастами. Эти органы могут быть рассеяны поодиночке или собраны под чешуёй в совокупности каналов, которые называются боковой линией. Нейромасты включают полушарие гелевой консистенции (капулу) и сенсорные волосовидные клетки, а также синапсы нервных волокон, которые находятся на волосовидных клетках. Ток воды отгибает волосовидные клетки, вызывая нервные импульсы. Эти импульсы позволяют составить довольно подробную картину окружающей среды: некоторые виды рыб, лишённые глаз, целиком ориентируются и перемещаются, полагаясь только на органы боковой линии. Ощущение электрического поля — электрорецепция — присуще многим видам рыб — не только тем, которые могут сами генерировать электрические разряды. Электрические сигналы улавливаются с помощью специальных ямок на поверхности тела. Эти ямки заполнены гелеподобным веществом, которое проводит электрический ток и содержит в себе электрорецепторные клетки, которые образуют синапсы с нейронами.

Плечевой пояс лягушки: 1 - грудина; 2 - хрящевая задняя часть грудины; 3 - коракоид; 4 - ключица; 5 - предгрудина; 6 -- лопатка. Хрящи помечены точками.

Сердце лягушки
 Присутствуют характерные для амниот межрёберные мышцы, играющие важную роль при")
Скелет ящерицы (отр. Чешуйчатые) Присутствуют характерные для амниот межрёберные мышцы, играющие важную роль при дыхании. Дыхание при помощи грудной клетки (у кого она есть и подвижна!). Подкожная мускулатура позволяет изменять положение роговых чешуй.

Скелет болотной черепахи. А — карапакс; Б — пластрон: 1 — туловищный отдел позвоночного столба, 2 — реберные пластинки, 3 — краевые пластинки, 4 — коракоид, 5 — лопатка, 6 — подвздошная кость, 7 — лобковая кость, 8 — седалищная кость

В осевом скелете пресмыкающихся разделение на отделы более заметно, чем у земноводных. Хорошо различимы 5 отделов позвоночника: шейный, туловищный, поясничный, крестцовый и хвостовой. Типичным для пресмыкающихся является следующее строение осевого скелета. Общее количество позвонков различно у разных видов (50— 80, у змей возрастает до 140— 435). Из позвонков шейного отдела (от 7 до 10) два передних (атлант и эпистрофей) образуют сустав, позволяющий голове не только двигаться в вертикальной плоскости относительно первого шейного позвонка, но и поворачиваться. В туловищном отделе от 16 до 25 позвонков, каждый с парой рёбер. Первые несколько позвонков прикрепляются к грудине, образуя грудную клетку (отсутствует у змей). В крестцовом отделе всего 2 позвонка, к широким поперечным отросткам которых причленяется таз. Хвостовой отдел составляют несколько десятков (15— 40) постепенно уменьшающихся в размерах позвонков. Последние хвостовые позвонки представляют собой небольшие палочковидные косточки. В некоторых группах рептилий осевой скелет имеет отличия. У змей позвоночник отчетливо делится лишь на туловищный и хвостовой отделы, грудина отсутствует. У черепах позвонки туловищного отдела срастаются со спинным щитом панциря, вследствие чего неподвижны. Череп пресмыкающихся значительно более окостеневший, чем у земноводных. Лишь в обонятельной капсуле и слуховой области содержится небольшое количество хряща. Осевой и висцеральный отделы черепа эмбрионально формируются отдельно, но у взрослых особей срастаются в единое образование. В состав черепа входят как замещающие (первичные), так и многочисленные кожные (покровные, или вторичные) кости. Пояс передних конечностей сходен с поясом земноводных, отличаясь лишь более сильным развитием окостенения. Пара передних конечностей рептилий состоит из плеча, предплечья и кисти. Пара задних конечностей — из бедра, голени и стопы. На фалангах конечностей расположены когти.
, и")
Подвижность головы обеспечивается дифференцировкой первых двух шейных позвонков - атлáса, или атланта (atlas), и эпистрофéя (epistropheus). Атлáс имеет вид костного кольца, разделенного плотной связкой на верхнюю и нижнюю половины; через верхнее отверстие головной мозг соединяется со спинным; передняя поверхность нижней половины сочленяется с затылочным мыщелком черепа, а сзади в нижнее отверстие входит зубовидный отросток второго шейного позвонка - эпистрофея. Голова может поворачиваться в стороны на зубовидном отростке, а ее перемещение в вертикальной плоскости обеспечивается сочленением черепного мыщелка с атласом. Все это обеспечивает сложные движения головы, усиливаемые подвижностью всей шеи. Изучение эмбрионального развития показало, что зубовидный отросток образуется путем прирастания к эпистрофею тела атласа.

У позвоночных животных нервная трубка подразделяется на головной и спинной мозг. В эмбриональной стадии развития нервная трубка образует в передней части три вздутия - 3 мозговых пузыря, из которых развиваются отделы мозга: передний пузырь дает передний и промежуточный мозг , средний пузырь превращается в средний мозг , задний пузырь образует мозжечок и продолговатый мозг. Эти 5 отделов мозга характерны для всех позвоночных животных. Выделяют 5 отделов головного мозга пресмыкающихся. Передний мозг состоит из двух больших полушарий, от которых отходят обонятельные доли. Поверхность больших полушарий абсолютно гладкая. Большую часть крыши полушарий занимает древняя кора — архипаллиум. Впервые у рептилий появляется зачаток новой коры - неопаллиум. Дно переднего мозга в основном состоит из полосатых тел. Полушария переднего мозга наползают на другие отделы, вследствие чего образуется изгиб в области промежуточного мозга. Промежуточный мозг расположен между передним и средним мозгом. В верхней его части расположен эпифиз, а на нижней стороне — гипофиз. У большинства ящериц и гаттерии (а также многих вымерших форм) рядом с эпифизом развивается теменной глаз, а у крокодилов потеряны оба этих органа. Дно промежуточного мозга занято зрительными нервами и их перекрестом (хиазмой). Средний мозг представлен двумя крупными передними холмами — зрительными долями, а также небольшими задними холмами. Зрительная кора более развита, чем у амфибий.

Двигательная система пресмыкающихся усложняется, обеспечивая освоение всех наземных местообитаний, а у части вымерших и современных видов - и водоемов. Основное направление эволюции - усиление подвижности и маневренности движения, что расширяло возможности добывания пищи, спасения от врагов, поисков условий комфорта. Менее распространенное направление - выработка пассивных защитных приспособлений - костно-роговых щитов, шипов и других образований, ограничивающих подвижность (черепахи, многие вымершие рептилии). "Типичное" исходное строение скелета имеют пресмыкающиеся, передвигающиеся относительно медленно на четырех ногах. При "пресмыкании" с помощью расставленных по бокам тела коротких парных конечностей, примерно равных другу, туловище относительно подвижно, хвост умеренно длинен и шея коротка (гаттерия, большинство ящериц). Приспособлением к движению по сыпучему грунту служит появление на пальцах оторочки из роговых пластинок ("песчаные лыжи") и уплощение тела (ящерицы, круглоголовки и др. ). Ускорение движения по суше достигалось увеличением относительных размеров и изменением постановки конечностей: перенос конечностей под туловище позволяет приподнять его над грунтом и ускорить передвижение (многие пустынные ящерицы; крокодилы, изменяя постановку ног, могут бежать галопом со скоростью 12 км в час). Еще больше повысил скорость переход к двуногому передвижению, характерному для многих вымерших динозавров и некоторых пустынных ящериц (плащеносной; пустынной агамы, некоторых игуан). При этом увеличивались размеры хвоста, игравшего роль балансира, а передние конечности стали служить преимущественно для удерживания пищи; иногда они уменьшались до функционально почти бесполезного придатка. Сильное развитие задних конечностей даже позволяет молодняку шлемоносного василиска бегать по поверхности воды. При обитании в густых зарослях трав, в кронах деревьев, в лесной подстилке или на сыпучих субстратах оказался выгодным переход к ползанию, сопровождающийся утерей конечностей.
 изгибы тела в горизонтальной плоскости, при котором отдельные")
Змеевидные движения осуществляются тремя способами: а) изгибы тела в горизонтальной плоскости, при котором отдельные участки тела опираются на грунт и создают толкающую силу; след змеи при этом извитой, а скорость движения составляет 5 -8 км/ч; б) обитатели песков при так называемом "боковом ходе" забрасывают, не касаясь субстрата, заднюю часть тела вперед и в сторону, опираясь на покоящуюся часть тела, затем переносят впереднюю часть тела и операция повторяется. При таком движении на песке остаются параллельные полоски поперек направления хода; в) при "гусеничном передвижении" по телу пробегает волна сокращения, толкающая его вперед при опоре на расширенные брюшные щитки. Змеевидное тело эффективно и при передвижении в воде; совершенствование "водной специализации" выразилось в появлении морских змей с сжатым с боков хвостом. Уплощение тела при боковом сплющивании хвоста и приобретении плавательных перепонок между пальцами характерно для крокодилов и многих ископаемых архозавров. У черепах переход к водному образу жизни сопровождался уплощением, а затем и частичной редукцией панциря и превращением лап в ласты. Наконец, ихтиозавры приобретают сходство с рыбами и в форме тела, и в строении конечностей, напоминающих плавники. Летающие ящеры, видимо, жившие по опушкам леса и береговым обрывам, вероятно, приобрели способность к активному полету через планирование с помощью складки кожи, натянутой между боками тела и конечностями. Характерный для древнейших птерозавров - рамфоринхов хвост при усовершенствовании активного (машущего) полета укорачивался до почти полного исчезновения (птеранодон). По общему облику птерозавры удивительно напоминают современных летучих мышей (это один из ярких примеров конвергенции). Иным путем перешли к полету птицы.

Можно заключить, что в классе пресмыкающихся сложились все основные типы передвижений, присущие надклассу четвероногих в целом. Именно в этом основная причина победы пресмыкающихся над земноводными и поразительного по своей длительности и эффекту господства рептилий на протяжении всей мезозойской эры, проявившегося в освоении всех сред обитания на Земле. Их биологический успех выразился и в том, что пресмыкающиеся стали предками двух господствующих ныне в биосфере классов - птиц и млекопитающих. Оба эти класса, как будет показано далее, не выработали принципиально новых способов движения, и причины их успеха лежали не столько в совершенствовании двигательной системы, сколько в повышении общего уровня метаболизма, в усложнении нервной системы и поведения, позволившим использовать, уже сложившиеся "конструктивные схемы" с неизмеримо большей эффективностью. Наружный кожный покров пресмыкающихся в результате утолщения и синтеза кератина образует чешуйки или щитки. У ящериц роговые чешуйки перекрывают друга, напоминая черепицу. У черепах сросшиеся щитки формируют сплошной прочный панцирь. Смена рогового покрова происходит путём полной или частичной линьки, которая у многих видов происходит несколько раз в год. Плотная и сухая кожа содержит пахучие железы. Слизистые железы отсутствуют. В наружной части внутреннего слоя кожи часто находятся специальные клетки — хроматофоры. В этих клетках секретируются пигменты: меланины и каротиноиды. Также в хроматофорах встречается способный отражать свет гуанин. Благодаря хроматофорам, некоторые пресмыкающиеся способны изменять окраску своего тела за сравнительно короткое время. Хамелеоны — наиболее известные представители с подобным свойством.

Схема сердца и прилежащих сосудов крокодилы большинство пресмыкающихся

Пресмыкающиеся имеют 6 основных органов чувств: Глаза устроены сложнее, чем у амфибий: в склере присутствует кольцо из тонких костных пластинок; от задней стенки глазного яблока отходит вырост — гребешок, вдающийся в стекловидное тело; есть поперечно-полосатая мускулатура, которая позволяет не только перемещать хрусталик, но и изменять его форму, таким образом осуществляя наводку на резкость в процессе аккомодации. Органы зрения имеют приспособления к работе в воздушной среде. Слёзные железы предохраняют глаза от высыхания. Наружные веки и мигательная перепонка выполняют защитную функцию. У змей и некоторых ящериц веки срастаются, формируя прозрачную оболочку. Сетчатка глаза может содержать как палочки, так и колбочки. У ночных видов колбочки отсутствуют. У большинства дневных видов диапазон цветного зрения смещён в жёлто-оранжевую часть спектра. Зрение имеет решающее значение среди органов чувств пресмыкающихся. Теменной глаз, сохранившийся у некоторых видов, важен для ориентирования в пространстве и синхронизации суточных ритмов организма с циклом смены дня и ночи, хотя многое в его функциях ещё неясно. Орган обоняния представлен внутренними ноздрями — хоанами и вомероназальным органом. По сравнению со строением земноводных, хоаны расположены ближе к глотке, что даёт возможность свободно дышать в то время, как пища находится во рту. Обоняние развито лучше, чем у земноводных, позволяя многим ящерицам находить пищу, находящуюся под поверхностью песка на глубине до 6— 8 см. Орган вкуса — вкусовые луковицы, расположенные в основном в глотке. Орган тепловой чувствительности находится на лицевой ямке между глазом и носом с каждой стороны головы. Особенно развит у змей. У ямкоголовых змей термолокаторы позволяют определять даже направление источника теплового излучения. Орган слуха близок к органу слуха лягушек, он содержит внутреннее и среднее ухо, снабженное барабанной перепонкой, слуховой косточкой — стременем и евстахиевой трубой. Роль слуха в жизни пресмыкающихся сравнительно невелика, особенно слаб слух у змей, не имеющих барабанной перепонки и воспринимающих колебания, распространяющиеся по земле или в воде. Пресмыкающиеся воспринимают звуки в диапазоне 20— 6000 Гц, хотя большинство хорошо слышит лишь в диапазоне 60— 200 Гц (у крокодилов 100— 3000 Гц).
 Подкласс Анапсиды (Anapsida) Отряд † Captorhinida Отряд Черепахи (Testudines или Chelonia)")
Класс Пресмыкающиеся (Reptilia) Подкласс Анапсиды (Anapsida) Отряд † Captorhinida Отряд Черепахи (Testudines или Chelonia) Инфракласс Архозавроморфы Отряд † Котилозавры (Cotylosauria) Отряд † Rhynchosauria Отряд † Procolophonomorpha Отряд † Prolacertiformes Подотряд † Мезозавры (Mesosauria) = (Proganosauria) Отряд † Проторозавры (Protorosauria) Подкласс Диапсиды (Diapsida) Группа Архозавры (Archosauria) Отряд † Araeoscelidia Группа Круротарзы (Crurotarsi) Группа † Avicephala Надотряд Crocodylomorpha Группа Эозухии (Eosuchia) Отряд Крокодилы (Crocodylia или Crocodilia) — Группа † Acerosodontosauridae единственные выжившие современные представители Группа Neodiapsida Отряд † Sphenosuchia Отряд † Araeosceloidea Отряд † Rauisuchia Отряд † Ихтиозавры (Ichthyosauria) Отряд † Aetosauria Отряд † Younginiformes Отряд † Phytosauria Надсемейство † Younginoidea Группа † Avemetatarsalia Инфракласс Lepidosauromorpha Группа † Ornithodira Надотряд † Зауроптеригии (Sauropterygia) Группа † Dinosauriformes Отряд † Placodontia Семейство † Silesauridae Отряд † Nothosauroidea Отряд † Птерозавры или летающие ящеры (Pterosauria): Отряд † Плезиозавры (Plesiosauria) Надотряд † Динозавры (Dinosauria) (кроме птиц) Надотряд Лепидозавры или чешуйчатые ящеры (Lepidosauria) Отряд † Ящеротазовые динозавры (Saurichia) — вымерли, Отряд Клювоголовые или хоботноголовые (Rhynchocephalia) возможно, дали начало птицам Отряд Чешуйчатые (Squamata): ящерицы и змеи Отряд † Птицетазовые динозавры (Ornithischia)

Череп черепахи логгерхеда Caretta caretta

Типичное для синапсид строение черепа. Кости: j: скуловая; p: теменная; po: заглазничная; q: квадратная; qj: квадратно-скуловая; sq: чешуйчатая. Строение черепа анапсид Строение черепа диапсид
")
Отряд Черепахи (Testudines или Chelonia)
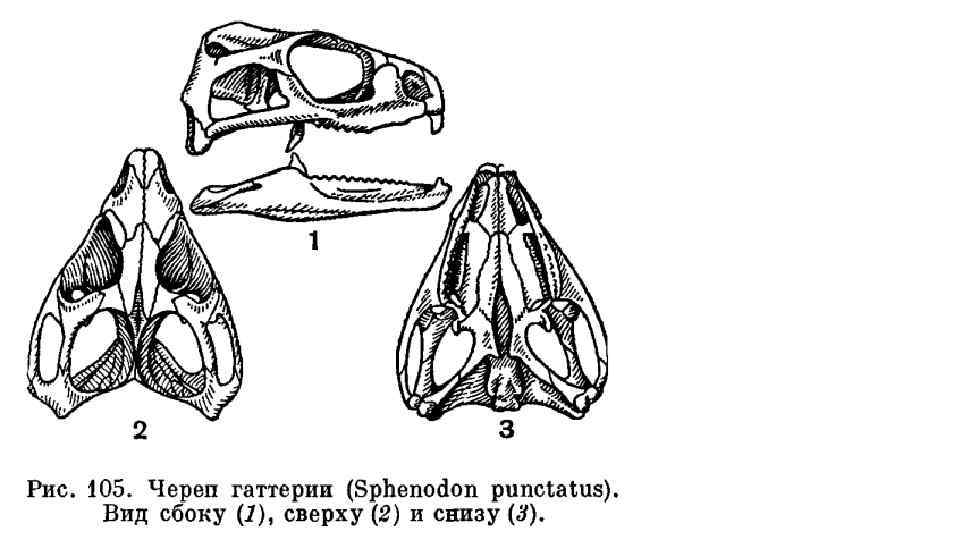
 Гатте рия, или туата ра (лат. Sphenodon punctatus) —")
Отряд Клювоголовые или хоботноголовые (Rhynchocephalia) Гатте рия, или туата ра (лат. Sphenodon punctatus) — вид пресмыкающихся, единственный современный представитель древнего отряда клювоголовых (иногда выделяют второй вид, Sphenodon guntheri). Обитает на нескольких небольших островах Новой Зеландии (на двух главных островах, Северном и Южном, вымерла). Длина тела достигает 76 см. Масса тела достигает 1, 3 кг. Средняя продолжительность жизни — 60 лет, однако могут жить и более 100 лет. Половой зрелости достигают в возрасте 15— 20 лет. Самки спариваются и откладывают яйца (от 8 до 15) раз в четыре года. От спаривания до вылупления проходит 12— 15 месяцев. Ведёт преимущественно ночной образ жизни. Внешним видом и образом жизни напоминает крупных игуан. В черепе: непосредственное сочленение сошников и крыловидных костей, утрата слезной и верхневисочной костей. Зубы у гаттерии простой клиновидной формы; они, как у большинства рептилий, прирастают к верхнему краю нижней и нижнему краю верхней челюстей (акродонтные). У взрослых животных зубы стираются настолько, что укус производится уже самими краями челюстей, покровы которых ороговевают. На нёбной кости расположен второй ряд зубов; зубы нижней челюсти входят между этими двумя зубными рядами. Позвонки сохраняют примитивное двояковогнутое (амфицельное) строение. Утраченный хвост регенерирует. Помимо обычных ребер, несущих направленные назад крючковидные отростки, имеется еще серия так называемых брюшных ребер, расположенных между грудиной и тазом под кожей. Среди современных пресмыкающихся и крючковидные отростки, и брюшные ребра, кроме гаттерии, сохранились только у крокодилов. В плечевом поясе, кроме лопатки и коракоида, есть ключицы и непарная межключица.
 • Инфраотряд Alethinophidia • Инфраотряд Scolecophidia • Подотряд амфисбены,")
• Подотряд змеи (Serpentes) • Инфраотряд Alethinophidia • Инфраотряд Scolecophidia • Подотряд амфисбены, или двуходки (Amphisbaenia) • Подотряд ящерицы (Lacertilia) • Инфраотряд варанообразные (Varanoidea) • Инфраотряд веретеницеобразные (Anguim orpha)) • Инфраотряд гекконообразные (Gekkota) • Инфраотряд игуанообразные (Iguania) • Инфраотряд сцинкообразные (Scincomorph a) Отряд Чешуйчатые (Squamata): ящерицы и змеи

Тело червеобразное, цилиндрически вытянутое, покрыто цельной роговой плёнкой, опоясано узкими поперечными кольцами, которые пересекаются более или менее явственными продольными бороздками. Каждое кольцо разбивается на правильные прямоугольники или квадраты, по внешнему виду несколько напоминающие чешую. Двуходок отличает сильное окостенение изменчивого по форме черепа, лишённого черепных дуг и характерных для ящериц столбчатых костей. Немногочисленные плевродонтные или, реже, акродонтные зубы располагаются лишь в передней части челюстей и варьируются по форме и величине. Эмбриональный яйцевой зуб не исчезает с возрастом, а, увеличиваясь в размерах, сохраняется на всю жизнь. В отличие от большинства змей и безногих ящериц, обладающих единственным правым лёгким, у двуходок развито одно левое.

Инфраотряд Alethinophidia Надсемейство Henophidia — Низшие змеи Семейство Aniliidae — Вальковатые змеи Семейство Anomochilidae Семейство Boidae — Ложноногие змеи Подсемейство Boinae Gray — Удавы Подсемейство Erycinae Bonaparte — Песчаные удавы Подсемейство Pythoninae — Питоны Семейство Bolyeriidae — Болиериды Семейство Cylindrophiidae — Цилиндрические змеи Семейство Loxocemidae — Двухцветные змеи Семейство Tropidophiidae — Земляные удавы Семейство Uropeltidae — Щитохвостые змеи Семейство Xenopeltidae — Лучистые змеи Надсемейство Colubroidea Семейство Acrochordidae — Бородавчатые змеи Семейство Atractaspididae — Шпильковые змеи Семейство Colubridae — Ужеобразные Подсемейство Homalopsinae Günther — Пресноводные змеи Подсемейство Natricinae Bonaparte — Ужовые Подсемейство Xenodontinae Cope — Гладкозубые змеи Семейство Elapidae — Аспиды Семейство Hydrophiidae — Морские змеи Семейство Viperidae — Гадюковые Подсемейство Azemiopinae Liem, Marx & Rabb Подсемейство Causinae Cope Подсемейство Crotalinae Oppel — Ямкоголовые Подсемейство Viperinae Oppel — Гадюковые

Инфраотряд Scolecophidia Надсемейство Typhlopoidea Семейство Anomalepididae — Американские червеобразные змеи, включает четыре рода распространённых в Центральной и Южной Америке; Семейство Gerrhopilidae, включает один род Gerrhopilus Семейство Leptotyphlopidae — Узкоротые змеи, включает два рода распространённых в Африке, Западной Азии и Америке; Семейство Typhlopidae — Слепозмейки, включает шесть родов распространённых в тропических и субтропических районах мира. Семейство Xenotyphlopidae, включает один род Xenotyphlops[2]
Земноводные_пресмыкающиеся.pptx