

частная физиология сенсорных систем 2008.ppt
- Количество слайдов: 127
 Частная физиология сенсорных систем
Частная физиология сенсорных систем
 Сенсорные системы мозга это совокупность сенсорных рецепторов, вспомогательных аппаратов, нейронов мозга, участвующих в обработке информации о сигналах внешнего или внутреннего мира
Сенсорные системы мозга это совокупность сенсорных рецепторов, вспомогательных аппаратов, нейронов мозга, участвующих в обработке информации о сигналах внешнего или внутреннего мира


, проприоцептивная, вкусовая,") сенсорные системы: • • зрительная, слуховая, вестибулярная, соматическая (тактильная, температурная и болевая), проприоцептивная, вкусовая, обонятельная, висцеральная (интероцептивная)
сенсорные системы: • • зрительная, слуховая, вестибулярная, соматическая (тактильная, температурная и болевая), проприоцептивная, вкусовая, обонятельная, висцеральная (интероцептивная)
: • Периферический • Проводниковый • Центральный") Основные отделы (по И. П. Павлову): • Периферический • Проводниковый • Центральный
Основные отделы (по И. П. Павлову): • Периферический • Проводниковый • Центральный
 План лекции: § § § Физиология зрительного анализатора Физиология слухового анализатора Физиология вестибулярного анализатора
План лекции: § § § Физиология зрительного анализатора Физиология слухового анализатора Физиология вестибулярного анализатора
 Физиология зрительной сенсорной системы
Физиология зрительной сенсорной системы
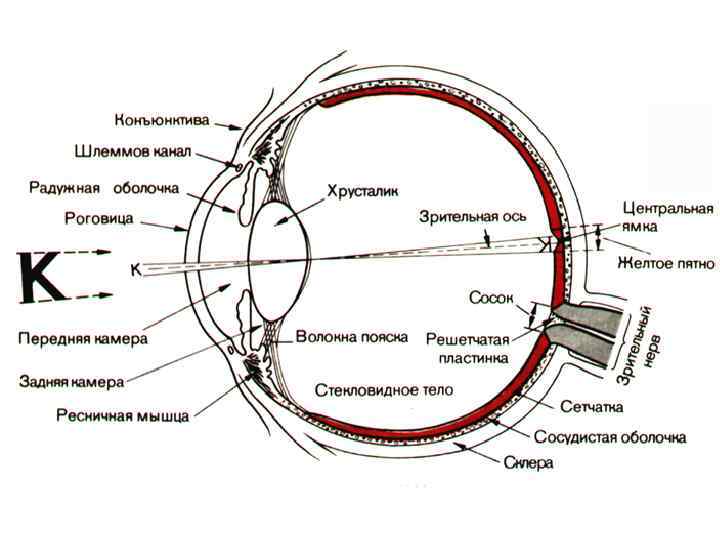
 Орган зрения - состоит из глазного яблока, зрительного нерва и вспомогательных аппаратов (веки и слёзные железы и глазодвигательные мышцы) Глаз • • • Оптическая часть (обеспечивает светопреломляющую функцию): роговица передняя камера глаза задняя камера глаза зрачок хрусталик стекловидное тело Фоторецепторная часть обеспечивает световосприятие фоторецепторы
Орган зрения - состоит из глазного яблока, зрительного нерва и вспомогательных аппаратов (веки и слёзные железы и глазодвигательные мышцы) Глаз • • • Оптическая часть (обеспечивает светопреломляющую функцию): роговица передняя камера глаза задняя камера глаза зрачок хрусталик стекловидное тело Фоторецепторная часть обеспечивает световосприятие фоторецепторы
 1. Периферический отдел зрительного анализатора
1. Периферический отдел зрительного анализатора
 функции рецептора • Обнаружение и различение сигнала • Кодирование информации переход энергии раздражителя в активность нервной клетки
функции рецептора • Обнаружение и различение сигнала • Кодирование информации переход энергии раздражителя в активность нервной клетки
 Фоторецепторы
Фоторецепторы
 • Клеточный состав сетчатки глаза
• Клеточный состав сетчатки глаза
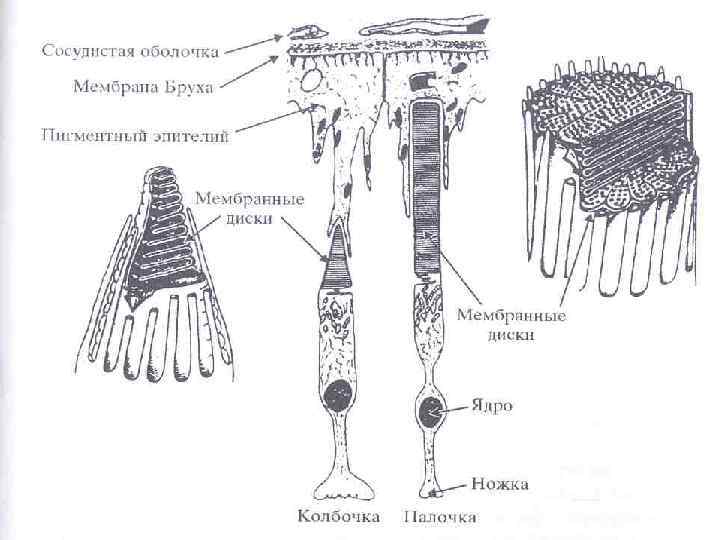
 Структура сетчатки • пигментный слой меланинсодержащих эпителиальных клеток • слой фоторецепторов • слой горизонатальных клеток • слой биполярных клеток • слой амакриновых клеток • слой ганглиозных клеток
Структура сетчатки • пигментный слой меланинсодержащих эпителиальных клеток • слой фоторецепторов • слой горизонатальных клеток • слой биполярных клеток • слой амакриновых клеток • слой ганглиозных клеток
 • Восприятие света осуществляется с участием фоторецепторов вторичночувствующих рецепторов (они представляют собой специализированные клетки)
• Восприятие света осуществляется с участием фоторецепторов вторичночувствующих рецепторов (они представляют собой специализированные клетки)
 Принцип работы фоторецепторов • Минимальное число квантов света для возникновения ощущения света - от 8 до 47. • Но одна палочка может быть возбуждена всего 1 квантом света.
Принцип работы фоторецепторов • Минимальное число квантов света для возникновения ощущения света - от 8 до 47. • Но одна палочка может быть возбуждена всего 1 квантом света.
 взаимодействие раздражителя с рецептором; • 2) изменение") Механизмы возбуждения в первичночувствующих рецепторах • 1) взаимодействие раздражителя с рецептором; • 2) изменение ионной проницаемости мембраны рецептора; • 3) генерация рецепторного потенциала • 4) генерация ПД в теле афферентного нейрона • 5) распространение ПД по аксону до второго афферентного нейрона
Механизмы возбуждения в первичночувствующих рецепторах • 1) взаимодействие раздражителя с рецептором; • 2) изменение ионной проницаемости мембраны рецептора; • 3) генерация рецепторного потенциала • 4) генерация ПД в теле афферентного нейрона • 5) распространение ПД по аксону до второго афферентного нейрона
 механизмы возбуждения во вторичночувствующих рецепторах • первые три этапа идут по такой же схеме; • 4 а) выделение квантов медиатора (например, ацетилхолина) в синапсе рецепторной клетки; • 5 а) ответ дендрита афферентного нейрона на выделение медиатора генерацией ВПСП или генераторного потенциала. • 4 и 5 идут так же, как и в первичночувствующих рецепторах.
механизмы возбуждения во вторичночувствующих рецепторах • первые три этапа идут по такой же схеме; • 4 а) выделение квантов медиатора (например, ацетилхолина) в синапсе рецепторной клетки; • 5 а) ответ дендрита афферентного нейрона на выделение медиатора генерацией ВПСП или генераторного потенциала. • 4 и 5 идут так же, как и в первичночувствующих рецепторах.
 Принцип фоторецепции. • В условиях темноты фоторецептор выделяет медиатор глутамат, который гиперполяризует биполярную клетку, соединенную с ганглиозной клеткой. • Поэтому в темноте биполярная клетка не возбуждает ганглиозную клетку и потенциалы действия не идут по зрительному нерву. • При попадании кванта света происходит «возбуждение» фоторецептора: внутренний сегмент фоторецепторной клетки гиперполяризуется и тем самым снижает выделение глутамата, происходит снятие тормозного влияния на биполярную клетку.
Принцип фоторецепции. • В условиях темноты фоторецептор выделяет медиатор глутамат, который гиперполяризует биполярную клетку, соединенную с ганглиозной клеткой. • Поэтому в темноте биполярная клетка не возбуждает ганглиозную клетку и потенциалы действия не идут по зрительному нерву. • При попадании кванта света происходит «возбуждение» фоторецептора: внутренний сегмент фоторецепторной клетки гиперполяризуется и тем самым снижает выделение глутамата, происходит снятие тормозного влияния на биполярную клетку.
 2 вида фоторецепторов
2 вида фоторецепторов
 Зависимость остроты зрения от положения стимула на сетчатке
Зависимость остроты зрения от положения стимула на сетчатке
 Потенциалы фоторецептора
Потенциалы фоторецептора
 Превращения зрительного пигмента в палочках • Фотон света • Родопсин • Метародопсин 1 • Возбуждение Метародопсин 2 Опсин • Метародопсин 3 • Трансретиналь 11 -цис-ретиналь • Трансретинол 11 -цис-ретинол • Изомеризация
Превращения зрительного пигмента в палочках • Фотон света • Родопсин • Метародопсин 1 • Возбуждение Метародопсин 2 Опсин • Метародопсин 3 • Трансретиналь 11 -цис-ретиналь • Трансретинол 11 -цис-ретинол • Изомеризация
 Механизм реакции палочек на фотон света МРН-2 - метародопсин-2; Т - трансдуцин ФДЭ - фосфодиэстераза
Механизм реакции палочек на фотон света МРН-2 - метародопсин-2; Т - трансдуцин ФДЭ - фосфодиэстераза
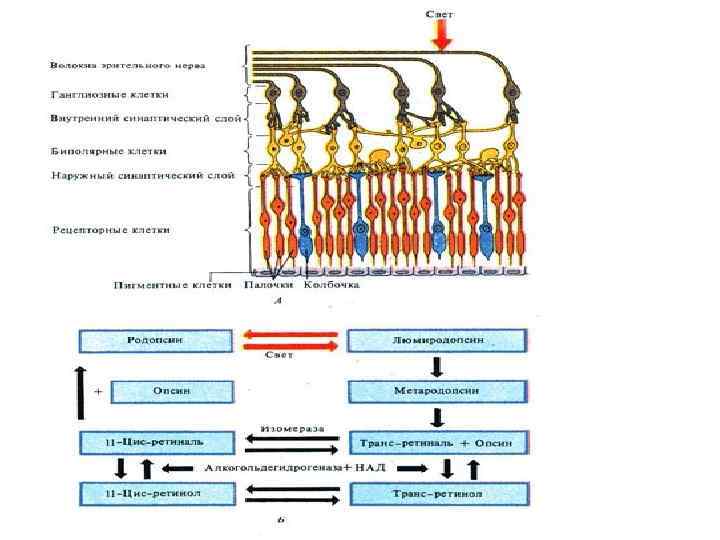
,") Механизм реакции на фотон света • образование метародпсина II, • активация трансдуцина (или G-белка), • активация фосфодиэстеразы, • снижение содержания ц. ГМФ, • закрытие натриевых каналов и гиперполяризация • снижение синтеза глутамата в фоторецепторе • деполяризация биполярной клетки (выделение ацетилхолина) • генерация ПД ганглиозной клеткой
Механизм реакции на фотон света • образование метародпсина II, • активация трансдуцина (или G-белка), • активация фосфодиэстеразы, • снижение содержания ц. ГМФ, • закрытие натриевых каналов и гиперполяризация • снижение синтеза глутамата в фоторецепторе • деполяризация биполярной клетки (выделение ацетилхолина) • генерация ПД ганглиозной клеткой
 Потенциалы клеток сетчатки
Потенциалы клеток сетчатки
 • Фоторецепторы объединяются в рецептивное поле – • это группа рецепторов, которые выходят на одну ганглиозную клетку • Все рецептивные поля сетчатки имеют концентрический вид • Диаметр рецептивного поля ганглиозной клетки, связанной с палочками - 1 мм.
• Фоторецепторы объединяются в рецептивное поле – • это группа рецепторов, которые выходят на одну ганглиозную клетку • Все рецептивные поля сетчатки имеют концентрический вид • Диаметр рецептивного поля ганглиозной клетки, связанной с палочками - 1 мм.
 возбуждается, если освещается круглый центр этого поля, а кольцевая") • Ганглиозная клетка (оn-нейрон) возбуждается, если освещается круглый центр этого поля, а кольцевая периферия не освещается. Если сразу осветить и центр, и периферию поля, то нейрон не возбуждается. • Ганглиозная клетка (оff-нейрон) будет возбуждаться только если освещается периферия.
• Ганглиозная клетка (оn-нейрон) возбуждается, если освещается круглый центр этого поля, а кольцевая периферия не освещается. Если сразу осветить и центр, и периферию поля, то нейрон не возбуждается. • Ганглиозная клетка (оff-нейрон) будет возбуждаться только если освещается периферия.
 Рецептивные поля ганглиозных клеток при воздействии цвета
Рецептивные поля ганглиозных клеток при воздействии цвета
 непрерывно генерируют ПД") Разновидности оn- и оff-нейронов: • Х-нейроны (например, на постоянное освещение центра) непрерывно генерируют ПД – это медленно адаптирующие нейроны. • У-нейроны возбуждаются только в момент включения или выключения светового сигнала – это быстроадаптирующие нейроны, • W-нейроны возбуждаются только в случае, если стимул «движется» , т. е. перемещается по сетчатке.
Разновидности оn- и оff-нейронов: • Х-нейроны (например, на постоянное освещение центра) непрерывно генерируют ПД – это медленно адаптирующие нейроны. • У-нейроны возбуждаются только в момент включения или выключения светового сигнала – это быстроадаптирующие нейроны, • W-нейроны возбуждаются только в случае, если стимул «движется» , т. е. перемещается по сетчатке.
 • Благодаря такой организации нейронов сетчатки уже на этом уровне происходит определение таких качеств как: • освещенность, • цвет, • форма, • движение сигнала.
• Благодаря такой организации нейронов сетчатки уже на этом уровне происходит определение таких качеств как: • освещенность, • цвет, • форма, • движение сигнала.


 Проводниковый и корковый отдел зрительного анализатора
Проводниковый и корковый отдел зрительного анализатора
 ЗРИТЕЛЬНЫЕ ПУТИ
ЗРИТЕЛЬНЫЕ ПУТИ
. Латеральное коленчатое тело (в") Зрительные нервы (после зрительного перекреста зрительные нервы называют зрительными трактами). Латеральное коленчатое тело (в составе зрительной лучистости, или пучка Грациоле) первичная проекционная область зрительной зоны коры (стриарная кора, или 17 -е поле по Бродману) вторичная проекционная зона (поле 18 и 19, престриарная кора) ассоциативные зоны коры
Зрительные нервы (после зрительного перекреста зрительные нервы называют зрительными трактами). Латеральное коленчатое тело (в составе зрительной лучистости, или пучка Грациоле) первичная проекционная область зрительной зоны коры (стриарная кора, или 17 -е поле по Бродману) вторичная проекционная зона (поле 18 и 19, престриарная кора) ассоциативные зоны коры
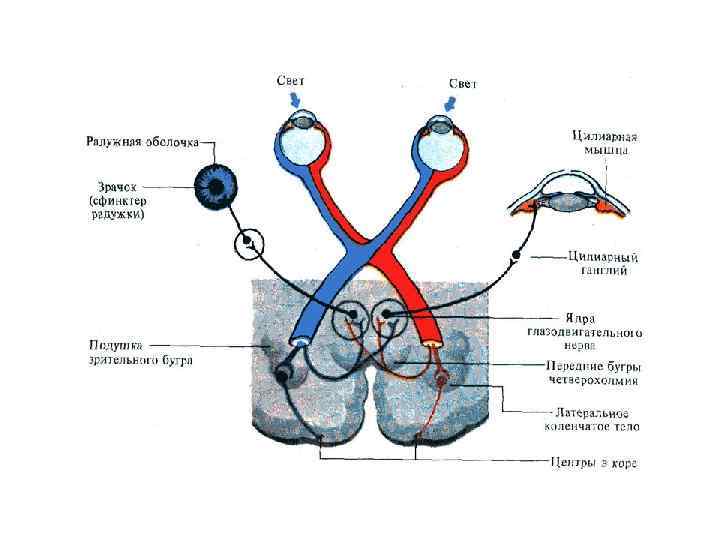
 • Возбуждение ганглиозных клеток сетчатки по их аксонам, составляющих зрительный нерв, поступает в мозг. • Ганглиозная клетка является первым нейроном зрительного анализатора. Волокна зрительного нерва образуют перекрест, • сетчатка одного глаза имеет контра- и ипсилатеральную проекцию. Большая часть волокон поступает в наружные коленчатые тела. Их аксоны идут в затылочную область коры, где расположена первичная проекционная зона зрительного анализатора. Часть волокон направляется в передние бугры четверохолмия и в таламус, от которого возбуждение поступает в кору.
• Возбуждение ганглиозных клеток сетчатки по их аксонам, составляющих зрительный нерв, поступает в мозг. • Ганглиозная клетка является первым нейроном зрительного анализатора. Волокна зрительного нерва образуют перекрест, • сетчатка одного глаза имеет контра- и ипсилатеральную проекцию. Большая часть волокон поступает в наружные коленчатые тела. Их аксоны идут в затылочную область коры, где расположена первичная проекционная зона зрительного анализатора. Часть волокон направляется в передние бугры четверохолмия и в таламус, от которого возбуждение поступает в кору.
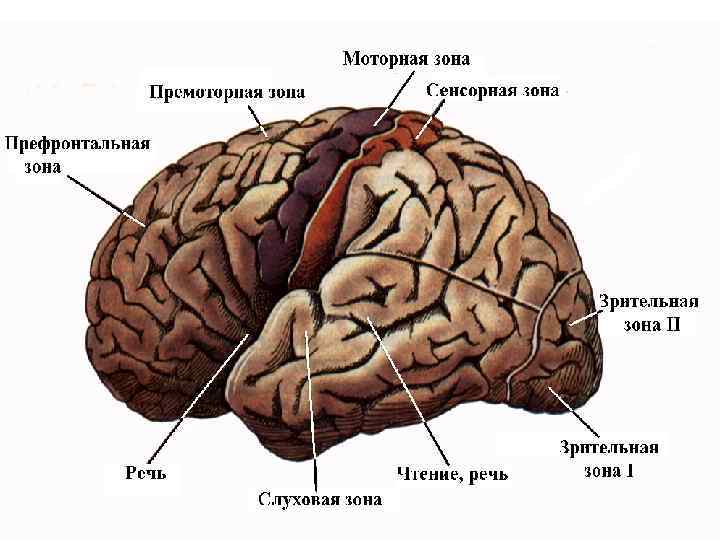
 Корковый отдел • поле 17 - точная ретинооптическая карта • поля 18 и 19 - нетопографические ассоциативные функции
Корковый отдел • поле 17 - точная ретинооптическая карта • поля 18 и 19 - нетопографические ассоциативные функции
 • Афферентные волокна из наружного коленчатого тела оканчиваются в слое IV и в глубине слоя III 17 -го поля, которое является центральным полем зрительной коры, а 18 -е и 19 -е поля – периферическими. Ассоциативные волокна из поля 17 направляются в 18 -е и 19 -е поля. • Между сетчаткой и полем 17 существует упорядоченное топографическое соответствие. Центральная ямка, где острота зрения максимальна, занимает большую часть коркового представительства. • Поля 18 и 19 выполняют нетопографические ассоциативные функции.
• Афферентные волокна из наружного коленчатого тела оканчиваются в слое IV и в глубине слоя III 17 -го поля, которое является центральным полем зрительной коры, а 18 -е и 19 -е поля – периферическими. Ассоциативные волокна из поля 17 направляются в 18 -е и 19 -е поля. • Между сетчаткой и полем 17 существует упорядоченное топографическое соответствие. Центральная ямка, где острота зрения максимальна, занимает большую часть коркового представительства. • Поля 18 и 19 выполняют нетопографические ассоциативные функции.
 • Вся информация от нейронов первичного и вторичных проекционных полей коры передается в передние и задние ассоциативные зоны коры, где с участием более «обученных» (гностических) нейронов происходит окончательное формирование образа. • В ассоциативных полях содержатся нейроны, обученные узнавать все буквы алфавита, слова, лица и т. д.
• Вся информация от нейронов первичного и вторичных проекционных полей коры передается в передние и задние ассоциативные зоны коры, где с участием более «обученных» (гностических) нейронов происходит окончательное формирование образа. • В ассоциативных полях содержатся нейроны, обученные узнавать все буквы алфавита, слова, лица и т. д.
 Роль ассоциативной коры В коре расположены: нейроны с простыми, сложными и «сверхсложными» рецептивными полями, ансамбли нейронов, выделяемые «под образы» , а также гностические ( «бабушкины» ) нейроны, узнающие определенный предмет внешнего мира
Роль ассоциативной коры В коре расположены: нейроны с простыми, сложными и «сверхсложными» рецептивными полями, ансамбли нейронов, выделяемые «под образы» , а также гностические ( «бабушкины» ) нейроны, узнающие определенный предмет внешнего мира
 нарушение") повреждение ассоциативных зон коры • агнозии- нарушение узнавания (слуховая, цветовая, тактильная агнозии) нарушение речи: • моторная афазия при нарушении передней ассоциативной зоны коры • сенсорная афазия при повреждении задней ассоциативной зоны коры.
повреждение ассоциативных зон коры • агнозии- нарушение узнавания (слуховая, цветовая, тактильная агнозии) нарушение речи: • моторная афазия при нарушении передней ассоциативной зоны коры • сенсорная афазия при повреждении задней ассоциативной зоны коры.
 Патология периферического отдела
Патология периферического отдела
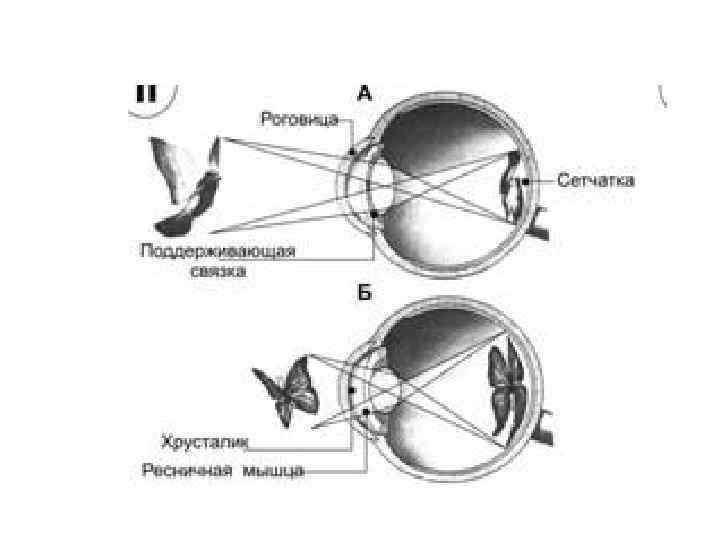
 • Аккомодация - — приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии • Основная роль в процессе аккомодации принадлежит хрусталику, способному изменять свою кривизну. • Рефракция - способность оптической системы глаза фокусировать изображение предмета на сетчатке глаза
• Аккомодация - — приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии • Основная роль в процессе аккомодации принадлежит хрусталику, способному изменять свою кривизну. • Рефракция - способность оптической системы глаза фокусировать изображение предмета на сетчатке глаза
 • Гиперметропия") Нарушения рефракции глаза • Миопия (преломляющая сила высокая изображение попадает перед сетчаткой) • Гиперметропия (преломляющая сила низкая - изображение попадает за сетчатку) • Астигматизм – неодинаковая диоптическая сила роговицы в разных плоскостях
Нарушения рефракции глаза • Миопия (преломляющая сила высокая изображение попадает перед сетчаткой) • Гиперметропия (преломляющая сила низкая - изображение попадает за сетчатку) • Астигматизм – неодинаковая диоптическая сила роговицы в разных плоскостях
 Миопия
Миопия
 Гиперметропия
Гиперметропия
 • максимальная способность глаза различать отдельные детали объектов • Острота зрения") острота зрения (visus) • максимальная способность глаза различать отдельные детали объектов • Острота зрения 1. 0 означает, что человек способен рассмотреть две отдельные точки, видимые под углом в 1 минуту.
острота зрения (visus) • максимальная способность глаза различать отдельные детали объектов • Острота зрения 1. 0 означает, что человек способен рассмотреть две отдельные точки, видимые под углом в 1 минуту.

 Таблицы повышенной точности Рожковой Г. И. , Токаревой В. С. , 2001
Таблицы повышенной точности Рожковой Г. И. , Токаревой В. С. , 2001

 § Психофизиологические особенности зрения
§ Психофизиологические особенности зрения
 Теории цветового зрения 1. Трехкомпонентная теория цветового зрения (М. В. Ломоносов, Т. Юнг, Г. Гельмгольц), цветовое восприятие обеспечивается тремя типами колбочек с различной спектральной (цветовой) чувствительностью.
Теории цветового зрения 1. Трехкомпонентная теория цветового зрения (М. В. Ломоносов, Т. Юнг, Г. Гельмгольц), цветовое восприятие обеспечивается тремя типами колбочек с различной спектральной (цветовой) чувствительностью.
 центр воспринимает все цвета (т.") Теории цветового зрения 2. Теория оппонентных цветов (Э. Геринг) центр воспринимает все цвета (т. е. колбочки чувствительны ко всем цветам – это дает ощущение белого цвета), а периферия – не чувствительна к цвету
Теории цветового зрения 2. Теория оппонентных цветов (Э. Геринг) центр воспринимает все цвета (т. е. колбочки чувствительны ко всем цветам – это дает ощущение белого цвета), а периферия – не чувствительна к цвету
 РЕЦЕПТИВНЫЕ ПОЛЯ off - центр on - центр
РЕЦЕПТИВНЫЕ ПОЛЯ off - центр on - центр
 Различия ганглиозных клеток по рецептивным полям Рецептивные поля больших ганглиозных клеток Рецептивные поля малых ганглиозных клеток
Различия ганглиозных клеток по рецептивным полям Рецептивные поля больших ганглиозных клеток Рецептивные поля малых ганглиозных клеток
 Цветовая слепота, или дальтонизм - дефект цветового восприятия дальтонизм у мужчин составляет 8% общей популяции лица, не различающие один цвет называются: Протанопы «краснослепые» Дейтеранопы «зеленослепые» Тританопы «синеслепые» Ахромазия– полная цветовая слепота (черно-белое восприятие) Диагностика - полихроматические таблицы Е. Рабкина
Цветовая слепота, или дальтонизм - дефект цветового восприятия дальтонизм у мужчин составляет 8% общей популяции лица, не различающие один цвет называются: Протанопы «краснослепые» Дейтеранопы «зеленослепые» Тританопы «синеслепые» Ахромазия– полная цветовая слепота (черно-белое восприятие) Диагностика - полихроматические таблицы Е. Рабкина


 Глаз человека воспринимает электромагнитное излучение с длиной волны 400 - 750 нм. Преломляющую силу выражают в диоптриях (D) Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила роговицы - 48, 8 D плоского хрусталика – 19, 1 D выпуклого хрусталика – 33, 0 D
Глаз человека воспринимает электромагнитное излучение с длиной волны 400 - 750 нм. Преломляющую силу выражают в диоптриях (D) Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила роговицы - 48, 8 D плоского хрусталика – 19, 1 D выпуклого хрусталика – 33, 0 D
 Возбудимость зрительных рецепторов
Возбудимость зрительных рецепторов
 БИНОКУЛЯРНОЕ ЗРЕНИЕ
БИНОКУЛЯРНОЕ ЗРЕНИЕ
 Стереоскопические рисунки
Стереоскопические рисунки
 Стереоскопические рисунки
Стереоскопические рисунки
 Бинокулярное зрение • – это одновременное видение предметов двумя глазами. Бинокулярное зрение обусловлено симметричным расположением глаз. В этом случае изображение предмета возникает на сетчатках обоих глаз, изображение попадает на соответствующие (корреспондирующие), или идентичные точки. • Фоторецепторы, расположенные на идентичных участках сетчатки, тесно связаны между собой функционально и находятся в одинаковом состоянии возбуждения. • Поэтому возникает один образ предмета. Доказано, что бинокулярное слитие или объединение сигналов от двух сетчаток в единый нервный образ происходит в первичной зрительной коре.
Бинокулярное зрение • – это одновременное видение предметов двумя глазами. Бинокулярное зрение обусловлено симметричным расположением глаз. В этом случае изображение предмета возникает на сетчатках обоих глаз, изображение попадает на соответствующие (корреспондирующие), или идентичные точки. • Фоторецепторы, расположенные на идентичных участках сетчатки, тесно связаны между собой функционально и находятся в одинаковом состоянии возбуждения. • Поэтому возникает один образ предмета. Доказано, что бинокулярное слитие или объединение сигналов от двух сетчаток в единый нервный образ происходит в первичной зрительной коре.
, точки сетчатки, то предмет") Бинокулярное зрение • Если же изображение падает на неидентичные, (диспарантные), точки сетчатки, то предмет начинает двоиться (если надавить слегка на один глаз сбоку, то начнется двоение в глазах, потому что нарушается соответствие сетчаток).
Бинокулярное зрение • Если же изображение падает на неидентичные, (диспарантные), точки сетчатки, то предмет начинает двоиться (если надавить слегка на один глаз сбоку, то начнется двоение в глазах, потому что нарушается соответствие сетчаток).
 и оценка расстояния до предмета. • В этих процессах важную") Восприятие глубины рельефа (пространства) и оценка расстояния до предмета. • В этих процессах важную роль играет бинокулярное зрение. • Каждый глаз видит предмет несколько сбоку, поэтому изображение на сетчатках получается не одинаковым. • Чем ближе находится предмет, тем больше будет разница в изображении, и в мозге, получающем соответствующие сигналы, создается представление о том или ином расстоянии до предмета.
Восприятие глубины рельефа (пространства) и оценка расстояния до предмета. • В этих процессах важную роль играет бинокулярное зрение. • Каждый глаз видит предмет несколько сбоку, поэтому изображение на сетчатках получается не одинаковым. • Чем ближе находится предмет, тем больше будет разница в изображении, и в мозге, получающем соответствующие сигналы, создается представление о том или ином расстоянии до предмета.
 и оценка расстояния до предмета. • Если смотреть на близкий") Восприятие глубины рельефа (пространства) и оценка расстояния до предмета. • Если смотреть на близкий предмет, конвергируя глаза, то изображение какой -либо более отдаленной точки попадает на неидентичные (диспаратные) точки двух сетчаток. • Таким образом, диспарация играет большую роль в оценке расстояния и в видении глубины рельефа.
Восприятие глубины рельефа (пространства) и оценка расстояния до предмета. • Если смотреть на близкий предмет, конвергируя глаза, то изображение какой -либо более отдаленной точки попадает на неидентичные (диспаратные) точки двух сетчаток. • Таким образом, диспарация играет большую роль в оценке расстояния и в видении глубины рельефа.
 Поле зрения левого глаза
Поле зрения левого глаза
 Физиология слухового анализатора
Физиология слухового анализатора
 • проводниковый отдел (акустическая") Слуховой анализатор • периферический отдел (наружное, среднее и внутреннее ухо) • проводниковый отдел (акустическая часть VIII пары черепно-мозгового нерва, кохлеарный комплекс, оливарный комплекс, трапециевидное тело, латеральный лемниск, нижние бугры четверохолмия, медиальное коленчатое тело) • корковый - (височная слуховая кора)
Слуховой анализатор • периферический отдел (наружное, среднее и внутреннее ухо) • проводниковый отдел (акустическая часть VIII пары черепно-мозгового нерва, кохлеарный комплекс, оливарный комплекс, трапециевидное тело, латеральный лемниск, нижние бугры четверохолмия, медиальное коленчатое тело) • корковый - (височная слуховая кора)
 Звукоулавливающий и звукопроводящий аппарат • наружное ухо: - ушная раковина и наружный слуховой проход • - среднее ухо: барабанная перепонка, косточки, мышцы среднего уха, евстахиева труба • - внутреннее ухо содержит рецепторный аппарат двух анализаторов: вестибулярного (преддверие и полукружные каналы) и слухового- улитка с кортиевым органом
Звукоулавливающий и звукопроводящий аппарат • наружное ухо: - ушная раковина и наружный слуховой проход • - среднее ухо: барабанная перепонка, косточки, мышцы среднего уха, евстахиева труба • - внутреннее ухо содержит рецепторный аппарат двух анализаторов: вестибулярного (преддверие и полукружные каналы) и слухового- улитка с кортиевым органом
 Схема строения уха
Схема строения уха
 Орган слуха и равновесия Среднее ухо Наружное ухо Внутреннее ухо
Орган слуха и равновесия Среднее ухо Наружное ухо Внутреннее ухо
 Слуховая система
Слуховая система
 Периферический отдел слухового анализатора Ухо человека чувствительно к тонам в диапазоне частот от 16 до 20000 Гц Единицей громкости звука является бел (Б). Бел - десятичный логарифм отношения действующей интенсивности звука I к пороговой его интенсивности Iо, т. е. L= lg I/ Iо. Звуковое давление выше 100 д. Б может повредить периферический слуховой аппарат
Периферический отдел слухового анализатора Ухо человека чувствительно к тонам в диапазоне частот от 16 до 20000 Гц Единицей громкости звука является бел (Б). Бел - десятичный логарифм отношения действующей интенсивности звука I к пороговой его интенсивности Iо, т. е. L= lg I/ Iо. Звуковое давление выше 100 д. Б может повредить периферический слуховой аппарат
 Психофизиологические характеристики слуха
Психофизиологические характеристики слуха
 Кортиев орган - базилярная мембрана с с наружными и") Периферический отдел слухового анализатора 1) Кортиев орган - базилярная мембрана с с наружными и внутренними рецепторными клетками 2) Текториальная мембрана – коллагеновые волокна и желатинозная масса, с нижней поверхностью которой связаны волоски рецепторных клеток
Периферический отдел слухового анализатора 1) Кортиев орган - базилярная мембрана с с наружными и внутренними рецепторными клетками 2) Текториальная мембрана – коллагеновые волокна и желатинозная масса, с нижней поверхностью которой связаны волоски рецепторных клеток
, трансформирующие механические колебания в электрические потенциалы. • Фонорецепторы") • Фонорецепторы вторичночувствующие рецепторы (механорецепторы), трансформирующие механические колебания в электрические потенциалы. • Фонорецепторы подразделяются на внутренние и наружные.
• Фонорецепторы вторичночувствующие рецепторы (механорецепторы), трансформирующие механические колебания в электрические потенциалы. • Фонорецепторы подразделяются на внутренние и наружные.
 • При действии звуков основная мембрана колеблется, волоски рецепторных клеток касаются покровной пластинки и деформируются, генерируют рецепторный потенциала и возбуждают слуховой нерв. Передача возбуждения с волосковой клетки на нервное волокно происходит через синапсы как электрическим, так и химическим путем.
• При действии звуков основная мембрана колеблется, волоски рецепторных клеток касаются покровной пластинки и деформируются, генерируют рецепторный потенциала и возбуждают слуховой нерв. Передача возбуждения с волосковой клетки на нервное волокно происходит через синапсы как электрическим, так и химическим путем.
 Разрез улиткового хода
Разрез улиткового хода
 Орган Корти
Орган Корти
 Орган Корти
Орган Корти
 Потенциал улитки
Потенциал улитки
 Глутамат
Глутамат
 Резонансная теория Г. Гельмгольца (1863): базилярная мембрана натянута в поперечном") Кодирование звука • 1) Резонансная теория Г. Гельмгольца (1863): базилярная мембрана натянута в поперечном направлении короткие волокна (ближе к овальному окну), резонируют на высокочастотные тоны, • длинные, (ближе к геликотреме), резонируют в ответ на низкочастотные тоны
Кодирование звука • 1) Резонансная теория Г. Гельмгольца (1863): базилярная мембрана натянута в поперечном направлении короткие волокна (ближе к овальному окну), резонируют на высокочастотные тоны, • длинные, (ближе к геликотреме), резонируют в ответ на низкочастотные тоны
 Теория «бегущей волны» Г. Бекеши (1960): под действием звука") Кодирование звуковой волны • 2) Теория «бегущей волны» Г. Бекеши (1960): под действием звука возникают колебания базилярной мембраны кортиева органа в виде бегущей волны, максимальная амплитуда которой зависит от частоты звука
Кодирование звуковой волны • 2) Теория «бегущей волны» Г. Бекеши (1960): под действием звука возникают колебания базилярной мембраны кортиева органа в виде бегущей волны, максимальная амплитуда которой зависит от частоты звука
 Пути слуховой сенсорной системы • Волосковые клетки органа Корти • Спиральный ганглий Первый перекрест • Кохлеарные ядра • Верхняя олива • Ядро латерального лемниска Второй перекрест • Нижнее двухолмие • Медиальные коленчатые тела • Слуховая кора
Пути слуховой сенсорной системы • Волосковые клетки органа Корти • Спиральный ганглий Первый перекрест • Кохлеарные ядра • Верхняя олива • Ядро латерального лемниска Второй перекрест • Нижнее двухолмие • Медиальные коленчатые тела • Слуховая кора
 Пути слуховой сенсорной системы
Пути слуховой сенсорной системы
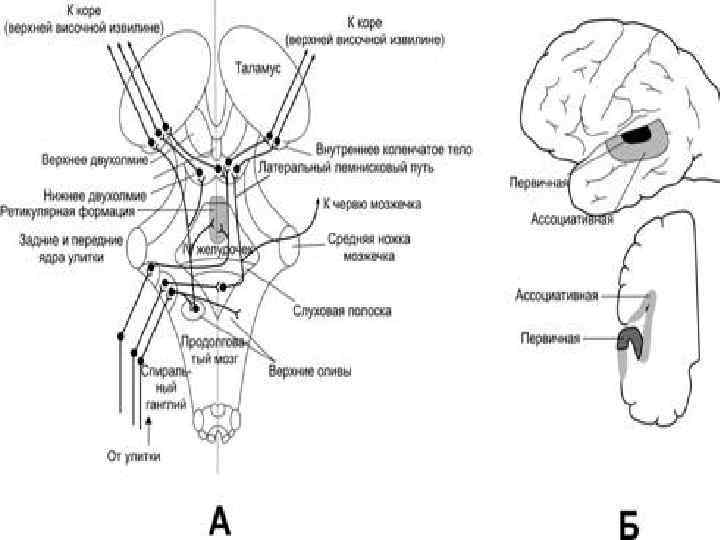
 Первичная слуховая кора: 41 поле извилины Гешля височной") Корковый отдел слухового анализатора • 1) Первичная слуховая кора: 41 поле извилины Гешля височной доли в глубине сильвиевой борозды и 42 поле верхней височной извилины – ощущение тонов, шумов, звуков • 2) Вторичная слуховая кора: 22 поле верхней височной извилины – понимание последовательности тонов (мелодии), последовательности звуков (слов), последовательности слов (фраз)
Корковый отдел слухового анализатора • 1) Первичная слуховая кора: 41 поле извилины Гешля височной доли в глубине сильвиевой борозды и 42 поле верхней височной извилины – ощущение тонов, шумов, звуков • 2) Вторичная слуховая кора: 22 поле верхней височной извилины – понимание последовательности тонов (мелодии), последовательности звуков (слов), последовательности слов (фраз)
 • В зонах слуховой коры происходит сложнейший анализ звукового сигнала, представление о последовательности слов или мелодии и определение локализации звука в пространстве
• В зонах слуховой коры происходит сложнейший анализ звукового сигнала, представление о последовательности слов или мелодии и определение локализации звука в пространстве
 Вестибулярная сенсорная система
Вестибулярная сенсорная система
") Периферический отдел представлен вестибулярным аппаратом (орган равновесия)
Периферический отдел представлен вестибулярным аппаратом (орган равновесия)
 орган равновесия • Костный лабиринт - система полукружных каналов и сообщающаяся с ними полость — преддверие. • Перепончатый лабиринт- система тонкостенных трубок и мешочков, расположенная внутри костного лабиринта.
орган равновесия • Костный лабиринт - система полукружных каналов и сообщающаяся с ними полость — преддверие. • Перепончатый лабиринт- система тонкостенных трубок и мешочков, расположенная внутри костного лабиринта.
") Рецепторные клетки • вторичночувствующие механорецепторы имеют на апикальной части один длинный подвижный волосок (киноцилия) и 60 -80 склеенных неподвижных (стереоцилия).
Рецепторные клетки • вторичночувствующие механорецепторы имеют на апикальной части один длинный подвижный волосок (киноцилия) и 60 -80 склеенных неподвижных (стереоцилия).
. • Мешочек,") • В мешочках преддверия находится отолитовый аппарат-скопление рецепторных клеток (вторичночувствующих механорецепторов). • Мешочек, расположенный ближе к улитке (саккулюс) • Мешочек, расположенный ближе к полукружным каналам (утрикулюс).
• В мешочках преддверия находится отолитовый аппарат-скопление рецепторных клеток (вторичночувствующих механорецепторов). • Мешочек, расположенный ближе к улитке (саккулюс) • Мешочек, расположенный ближе к полукружным каналам (утрикулюс).
 • Участки мешочков, содержащие чувствительные к гравитации и к линейным ускорениям клетки, называются пятнами, или макулами. • Участки ампул полукружных каналов, содержащие клетки, чувствительные к угловым ускорениям или замедлениям, называются гребешками, или кристами.
• Участки мешочков, содержащие чувствительные к гравитации и к линейным ускорениям клетки, называются пятнами, или макулами. • Участки ампул полукружных каналов, содержащие клетки, чувствительные к угловым ускорениям или замедлениям, называются гребешками, или кристами.
 • Киноцилии клеток пятен пронизывают толстую отолитовую мембрану содержащую кристаллики из углекислого кальция (Са. СОз); эти кристаллики называют отолитами.
• Киноцилии клеток пятен пронизывают толстую отолитовую мембрану содержащую кристаллики из углекислого кальция (Са. СОз); эти кристаллики называют отолитами.

 и пятна (Б, В). ОМ") Рецепторная область органа равновесия. Вертикальные срезы через гребешок (А) и пятна (Б, В). ОМ — отолитовая мембрана, О — отолиты, ПК — поддерживающая клетка, РК — рецепторная клетка.
Рецепторная область органа равновесия. Вертикальные срезы через гребешок (А) и пятна (Б, В). ОМ — отолитовая мембрана, О — отолиты, ПК — поддерживающая клетка, РК — рецепторная клетка.
 воспринимаю щие структуры гребешка
воспринимаю щие структуры гребешка
 • Проводниковый и корковый отдел
• Проводниковый и корковый отдел
 • Первыми нейронами, воспринимающими возбуждение волосковых клеток вестибулярного аппарата, являются биполярные нейроны вестибулярного ганглия, который залегает на дне внутреннего слухового прохода. • Их дендриты, контактирующие с волосковыми клетками, в ответ на возбуждение клетокрецепторов генерируют потенциалы действия, которые передаются в ЦНС по аксонам. Аксоны биполярных клеток образуют вестибулярную часть преддверно-улиткового нерва (VIII пара черепно-мозговых нервов).
• Первыми нейронами, воспринимающими возбуждение волосковых клеток вестибулярного аппарата, являются биполярные нейроны вестибулярного ганглия, который залегает на дне внутреннего слухового прохода. • Их дендриты, контактирующие с волосковыми клетками, в ответ на возбуждение клетокрецепторов генерируют потенциалы действия, которые передаются в ЦНС по аксонам. Аксоны биполярных клеток образуют вестибулярную часть преддверно-улиткового нерва (VIII пара черепно-мозговых нервов).
 • Волокна вестибулярной части нерва входят в области мостомозжечкового угла в продолговатый мозг и оканчиваются на нейронах бульбарного вестибулярного комплекса: • верхнее (ядро Бехтерева), • латеральное (ядро Дейтерса), • медиальное (ядро Швальбе) • нижнее (ядро Роллера).
• Волокна вестибулярной части нерва входят в области мостомозжечкового угла в продолговатый мозг и оканчиваются на нейронах бульбарного вестибулярного комплекса: • верхнее (ядро Бехтерева), • латеральное (ядро Дейтерса), • медиальное (ядро Швальбе) • нижнее (ядро Роллера).
 • Угловое ускорение→ смещение купола и сгибания ресничек волосковых клеток гребешков → деполяризация и возбуждение → рецепторный потенциал→ выброс ацетилхолина → афферентные окончания вестибулярного нерва → продолговатый мозг → вестибулярные ядра продолговатого мозга.
• Угловое ускорение→ смещение купола и сгибания ресничек волосковых клеток гребешков → деполяризация и возбуждение → рецепторный потенциал→ выброс ацетилхолина → афферентные окончания вестибулярного нерва → продолговатый мозг → вестибулярные ядра продолговатого мозга.
 Линейное ускорение → смещение эндолимфы и сгибания ресничек волосковых клеток пятен → деполяризация и возбуждение → рецепторный потенциал→ выброс ацетилхолина → афферентные окончания вестибулярного нерва → вестибулярные ядра продолговатого мозга.
Линейное ускорение → смещение эндолимфы и сгибания ресничек волосковых клеток пятен → деполяризация и возбуждение → рецепторный потенциал→ выброс ацетилхолина → афферентные окончания вестибулярного нерва → вестибулярные ядра продолговатого мозга.
 воспринимающие структуры гребешка
воспринимающие структуры гребешка
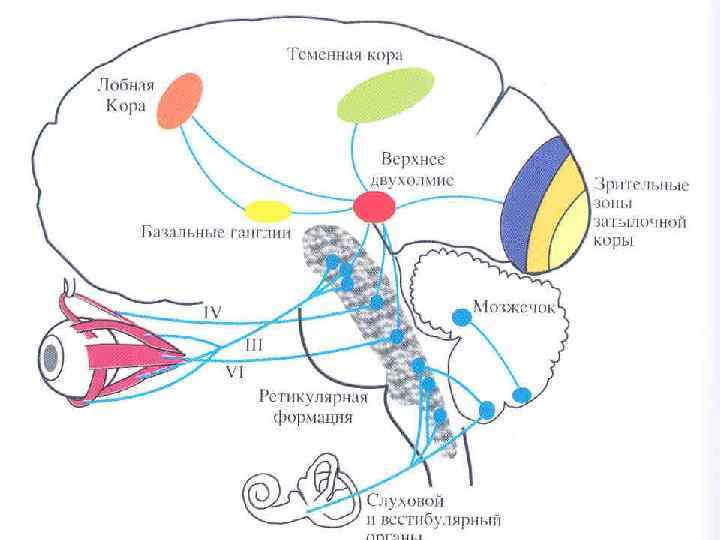
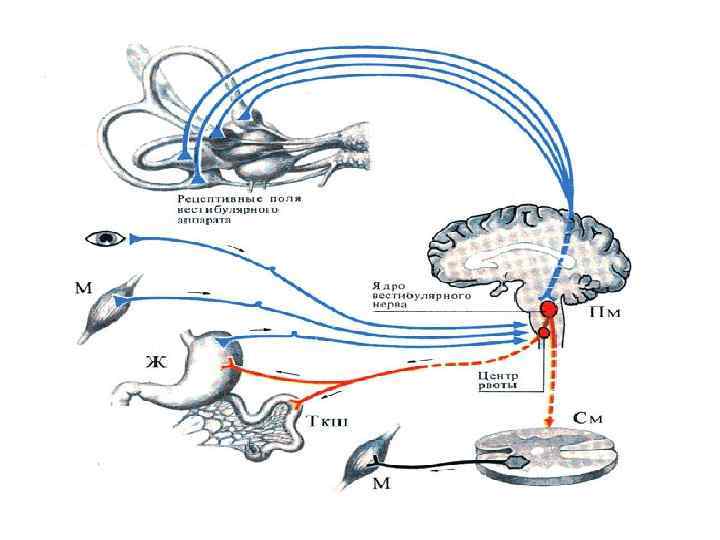
 • Вестибулярные ядра продолговатого мозга связаны со всеми отделами ЦНС: спинным мозгом, мозжечком, ретикулярной формацией ствола мозга, глазодвигательными ядрами, корой головного мозга, вегетативной нервной системой.
• Вестибулярные ядра продолговатого мозга связаны со всеми отделами ЦНС: спинным мозгом, мозжечком, ретикулярной формацией ствола мозга, глазодвигательными ядрами, корой головного мозга, вегетативной нервной системой.
 Пять главных проекционных систем. • 1. Вестибулоспинальный путь – это аксоны нейронов вестибулярных ядер, идущие к альфа -мотонейронам спинного мозга в виде двух продольных пучков – медиального и латерального. Благодаря вестибулоспинальному тракту вестибулярный аппарат участвует в регуляции мышечной активности (за счет реализации статических и статокинетических рефлексов) и тем самым способствует сохранению равновесия в условиях покоя и при движении.
Пять главных проекционных систем. • 1. Вестибулоспинальный путь – это аксоны нейронов вестибулярных ядер, идущие к альфа -мотонейронам спинного мозга в виде двух продольных пучков – медиального и латерального. Благодаря вестибулоспинальному тракту вестибулярный аппарат участвует в регуляции мышечной активности (за счет реализации статических и статокинетических рефлексов) и тем самым способствует сохранению равновесия в условиях покоя и при движении.
 • 2. Вестибуломозжечковая система, или вестибуломозжечковый путь, связывает вестибулярные ядра (через нижние ножки мозжечка) с ядрами шатра мозжечка, т. е. с архиоцеребеллумом, доставляя к нему информацию о положении тела в пространстве. Нарушение связи вестибулярного аппарата и мозжечка может приводить к развитию вестибулярной атаксии, т. е. к нарушению координации движений, что, например, наблюдается при болезни Миньера.
• 2. Вестибуломозжечковая система, или вестибуломозжечковый путь, связывает вестибулярные ядра (через нижние ножки мозжечка) с ядрами шатра мозжечка, т. е. с архиоцеребеллумом, доставляя к нему информацию о положении тела в пространстве. Нарушение связи вестибулярного аппарата и мозжечка может приводить к развитию вестибулярной атаксии, т. е. к нарушению координации движений, что, например, наблюдается при болезни Миньера.
 3. Вестибулоокулярный путь связывает вестибулярные ядра продолговатого мозга с альфа-мотонейронам глазодвигательных нервов (III, IV и VI пары черепно-мозговых нервов). Этот путь идет в составе медиального продольного пучка, который соединяет между собой все ядра глазодвигательных нервов. Эти связи играют важную роль в регуляции движений глаз, что необходимо для сохранения стабильного изображения на сетчатке во время движения. При возбуждении вестибулярных ядер возникают вестибуло-глазодвигательные рефлексы (нистагм глаз, нистагм головы).
3. Вестибулоокулярный путь связывает вестибулярные ядра продолговатого мозга с альфа-мотонейронам глазодвигательных нервов (III, IV и VI пары черепно-мозговых нервов). Этот путь идет в составе медиального продольного пучка, который соединяет между собой все ядра глазодвигательных нервов. Эти связи играют важную роль в регуляции движений глаз, что необходимо для сохранения стабильного изображения на сетчатке во время движения. При возбуждении вестибулярных ядер возникают вестибуло-глазодвигательные рефлексы (нистагм глаз, нистагм головы).
 нейронами гипоталамуса. При возбуждении вестибулярных ядер") вестибуло-гипоталамический путь связывает вестибулярные ядра с парасимпатическими (трофотропными) нейронами гипоталамуса. При возбуждении вестибулярных ядер повышается тонус парасимпатической нервной системы - тонус блуждающего и языкоглоточного нервов, что вызывает при выраженном раздражении вестибулярного аппарата вестибуло-вегетативные реакции – тошноту, рвоту, падение артериального давления, а также кинетоз, т. е. укачивание. 4.
вестибуло-гипоталамический путь связывает вестибулярные ядра с парасимпатическими (трофотропными) нейронами гипоталамуса. При возбуждении вестибулярных ядер повышается тонус парасимпатической нервной системы - тонус блуждающего и языкоглоточного нервов, что вызывает при выраженном раздражении вестибулярного аппарата вестибуло-вегетативные реакции – тошноту, рвоту, падение артериального давления, а также кинетоз, т. е. укачивание. 4.
 5. Лемнисковый путь – это путь, по которому информация от вестибулярных ядер поступает (после перекреста на уровне трапециевидного тела) через медиальную петлю (лемниск) к специфическим ядрам таламуса.
5. Лемнисковый путь – это путь, по которому информация от вестибулярных ядер поступает (после перекреста на уровне трапециевидного тела) через медиальную петлю (лемниск) к специфическим ядрам таламуса.
 Корковый отдел • первичная вестибулярная зона локализована в нижней области постцентральной извилины, в верхней и средней височных извилинах • вторичная зона в лобных долях, где в моторной зоне (спереди от нижней части центральной борозды) обнаружена вторая вестибулярная зона. • 21 - 22 поля по Бродману
Корковый отдел • первичная вестибулярная зона локализована в нижней области постцентральной извилины, в верхней и средней височных извилинах • вторичная зона в лобных долях, где в моторной зоне (спереди от нижней части центральной борозды) обнаружена вторая вестибулярная зона. • 21 - 22 поля по Бродману
 Связи вестибулярных ядер
Связи вестибулярных ядер
 Реакции организма, вызванные стимуляцией полукружных каналов • Головокружение • Нистагм
Реакции организма, вызванные стимуляцией полукружных каналов • Головокружение • Нистагм
 Благодарю за внимание!
Благодарю за внимание!