

Центральная рег сомат функц 2.ppt
- Количество слайдов: 63
 Центральная регуляция соматических функций
Центральная регуляция соматических функций
 Как сенсорный вход влияет на моторный выход?
Как сенсорный вход влияет на моторный выход?
 Всё бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению – мышечному движению. Смеётся ли ребёнок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к родине, дрожит ли девушка при первой мысли о любви, создаёт ли ньютон мировые законы и пишет их на бумаге – везде окончательным фактом является мышечное движение. И. М. Сеченов. «Рефлексы головного мозга» 1861 г.
Всё бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению – мышечному движению. Смеётся ли ребёнок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к родине, дрожит ли девушка при первой мысли о любви, создаёт ли ньютон мировые законы и пишет их на бумаге – везде окончательным фактом является мышечное движение. И. М. Сеченов. «Рефлексы головного мозга» 1861 г.






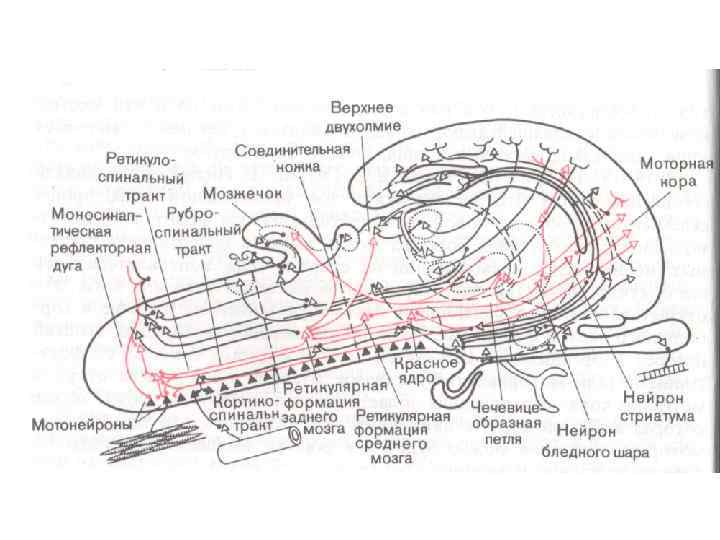











 Нисходящие длинноаксонные проприоспинальные связи спинного мозга Шейное Утолщение C 4 -Th 1 Поясничное Утолщение L 1 -S 3 - проходят на границе белого и серого вещества (красным) (Molenaar et al. , 1974, Schoenen, 2004) - связывают шейное и поясничное утолщения спинного мозга, обеспечивая координацию верхних и нижних конечностей (Skinner et al. 1979, Schomburg 1990)
Нисходящие длинноаксонные проприоспинальные связи спинного мозга Шейное Утолщение C 4 -Th 1 Поясничное Утолщение L 1 -S 3 - проходят на границе белого и серого вещества (красным) (Molenaar et al. , 1974, Schoenen, 2004) - связывают шейное и поясничное утолщения спинного мозга, обеспечивая координацию верхних и нижних конечностей (Skinner et al. 1979, Schomburg 1990)
 Стандартная методика регистрации Н-рефлекса нижней конечности человека В А Н-рефлекс вызывается при электрической стимуляции смешанного нерва (А), активируя чувствительные волокна и моносинаптически возбуждая альфа-мотонейроны, что приводит к синхронизированному эфферентному залпу и сокращению мышцы. При супрамаксимальной стимуляции происходит амплитудное подавление Н-рефлекса (В). - Позволяет оценивать нисходящие проприоспинальные связи при кондиционирующей стимуляции n. ulnaris (Коц 1975) и ритмических движениях рук (Shapkova 1998, Zehr 2007).
Стандартная методика регистрации Н-рефлекса нижней конечности человека В А Н-рефлекс вызывается при электрической стимуляции смешанного нерва (А), активируя чувствительные волокна и моносинаптически возбуждая альфа-мотонейроны, что приводит к синхронизированному эфферентному залпу и сокращению мышцы. При супрамаксимальной стимуляции происходит амплитудное подавление Н-рефлекса (В). - Позволяет оценивать нисходящие проприоспинальные связи при кондиционирующей стимуляции n. ulnaris (Коц 1975) и ритмических движениях рук (Shapkova 1998, Zehr 2007).
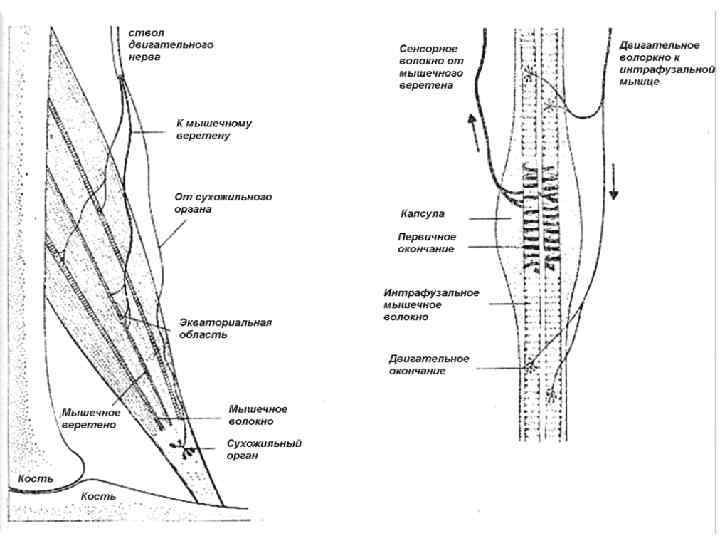
 Сенсорное обеспечение движения M. Latash, Neurophysiological Basis of Movement 2008 Рецептор Что измеряет Дополнительные факторы Iа веретёна II веретёна Длина, скорость Длина Чувствительность модулируется гамма-МН, мышечная длина отличается от длины «мышца+сухожилие» Ib Сухожильный орган Гольджи Сила измеряется в точке соединения мышцы и сухожилия. Сила зависит от длины и скорости. Суставные рецепторы Углы в суставах Чувствительнось максимальна около физиологических границ суставов и минимальна в среднем диапазоне. Суставные рецепторы чувствительны к давлению суставной капсулы. Кожные рецепторы Давление на кожу, смещение кожи Давление на кожу и смещение кожи может зависеть от многих факторов
Сенсорное обеспечение движения M. Latash, Neurophysiological Basis of Movement 2008 Рецептор Что измеряет Дополнительные факторы Iа веретёна II веретёна Длина, скорость Длина Чувствительность модулируется гамма-МН, мышечная длина отличается от длины «мышца+сухожилие» Ib Сухожильный орган Гольджи Сила измеряется в точке соединения мышцы и сухожилия. Сила зависит от длины и скорости. Суставные рецепторы Углы в суставах Чувствительнось максимальна около физиологических границ суставов и минимальна в среднем диапазоне. Суставные рецепторы чувствительны к давлению суставной капсулы. Кожные рецепторы Давление на кожу, смещение кожи Давление на кожу и смещение кожи может зависеть от многих факторов
") Нервные пути, участвующие в сгибательном и перекрестном разгибательном рефлексах. Болевое раздражение кожи конечности (А) активирует болевые афференты, синаптически возбуждающие интернейрон. Этот интернейрон возбуждает ипсилатералъные сгибателъные (С) мотонейроны (что приводит к сокращению мышцы-сгибателя и сгибанию колена) и тормозные интернейроны (в сером веществе), иннервирующие ипсилатеральные разгибательные (Р) мотонейроны. Одновременно активация и торможение соответствующих разгибателей и сгибателей другой ноги (Б) вызывают ее разгибание.
Нервные пути, участвующие в сгибательном и перекрестном разгибательном рефлексах. Болевое раздражение кожи конечности (А) активирует болевые афференты, синаптически возбуждающие интернейрон. Этот интернейрон возбуждает ипсилатералъные сгибателъные (С) мотонейроны (что приводит к сокращению мышцы-сгибателя и сгибанию колена) и тормозные интернейроны (в сером веществе), иннервирующие ипсилатеральные разгибательные (Р) мотонейроны. Одновременно активация и торможение соответствующих разгибателей и сгибателей другой ноги (Б) вызывают ее разгибание.

 Схемы взаимодействия лабиринтных и шейных рефлексов в случаях управления конечностями при различных положениях тела. (Roberts, in: Wilson, Melvill-Jones, 1979. ) А. Шейные и лабиринтные рефлексы противодействуют другу Б. Д. З. Действуют одни шейные рефлексы В. Шейные и лабиринтные рефлексы усиливают друга Г. Д. Е. Действуют одни лабиринтные позиционные рефлексы Ж. Шейные и лабиринтные рефлексы усиливают друга И. Шейные и лабиринтные рефлексы противодействуют другу
Схемы взаимодействия лабиринтных и шейных рефлексов в случаях управления конечностями при различных положениях тела. (Roberts, in: Wilson, Melvill-Jones, 1979. ) А. Шейные и лабиринтные рефлексы противодействуют другу Б. Д. З. Действуют одни шейные рефлексы В. Шейные и лабиринтные рефлексы усиливают друга Г. Д. Е. Действуют одни лабиринтные позиционные рефлексы Ж. Шейные и лабиринтные рефлексы усиливают друга И. Шейные и лабиринтные рефлексы противодействуют другу
 Функции мозжечка Мозжечок как надсегментарный орган, входящий в систему регуляции движений, выполняет следующие важные функции: 1) регуляция позы и мышечного тонуса; 2) сенсомоторная координация позных и целенаправленных движений; 3) координация быстрых целенаправленных движений, осуществляемых по команде из больших полушарий. Эти функции мозжечка наиболее удобно разбирать в соответствии с топической классификацией его отделов, основанной на характере эфферентных связей.
Функции мозжечка Мозжечок как надсегментарный орган, входящий в систему регуляции движений, выполняет следующие важные функции: 1) регуляция позы и мышечного тонуса; 2) сенсомоторная координация позных и целенаправленных движений; 3) координация быстрых целенаправленных движений, осуществляемых по команде из больших полушарий. Эти функции мозжечка наиболее удобно разбирать в соответствии с топической классификацией его отделов, основанной на характере эфферентных связей.



") Клетки-зёрна, параллельные волокна и клетки Пуркинье с развитой системой дендритов ( «деревом дендритов» )
Клетки-зёрна, параллельные волокна и клетки Пуркинье с развитой системой дендритов ( «деревом дендритов» )
 Нейрональные взаимосвязи коры мозжечка: MF — моховидные волокна; DCN — ядра мозжечка; IO — нижнее ядро оливы; CF — лазящие волокна; GC — клетки-зёрна; PF — параллельные волокна; Gg. C — клетка Гольджи; SC — звёздчатая клетка; BC — корзинчатая клетка
Нейрональные взаимосвязи коры мозжечка: MF — моховидные волокна; DCN — ядра мозжечка; IO — нижнее ядро оливы; CF — лазящие волокна; GC — клетки-зёрна; PF — параллельные волокна; Gg. C — клетка Гольджи; SC — звёздчатая клетка; BC — корзинчатая клетка
![Синаптические связи нейронов мозжечка [Экклс Дж. , 1969]. Схематически показаны возбуждающие синапсы, образуемые на](https://present5.com/presentation/12901826_437453235/image-36.jpg "Синаптические связи нейронов мозжечка [Экклс Дж. , 1969]. Схематически показаны возбуждающие синапсы, образуемые на") Синаптические связи нейронов мозжечка [Экклс Дж. , 1969]. Схематически показаны возбуждающие синапсы, образуемые на грушевидных нейронах (клетки Пуркинье) лазающими волокнами (ЛВ), аксонами клеток-зерен (КЗ), которые в свою очередь активируются мшистыми волокнами (MB), и тормозные синапсы образованы аксонами звездчатых (Зв. К) и корзинчатых клеток; Т — торможение: В — возбуждение.
Синаптические связи нейронов мозжечка [Экклс Дж. , 1969]. Схематически показаны возбуждающие синапсы, образуемые на грушевидных нейронах (клетки Пуркинье) лазающими волокнами (ЛВ), аксонами клеток-зерен (КЗ), которые в свою очередь активируются мшистыми волокнами (MB), и тормозные синапсы образованы аксонами звездчатых (Зв. К) и корзинчатых клеток; Т — торможение: В — возбуждение.
 Связи между мозжечком и красным ядром, таламусом, корой больших полушарий спинным мозгом, благодаря которым осуществляется мозжечковый контроль движений (по Гайтону).
Связи между мозжечком и красным ядром, таламусом, корой больших полушарий спинным мозгом, благодаря которым осуществляется мозжечковый контроль движений (по Гайтону).
 – синаптическое") Морфофункциональная организация таламуса По функции ядра таламуса можно разделить на: специфические (проекционные) – синаптическое переключение сенсорной информации с аксонов восходящих афферентных путей на следующие, конечные нейроны, отростки которых идут в соответствующие сенсорные проекционные области коры больших полушарий; ассоциативные – в отличие от специфических ядер они не могут быть отнесены к какой-либо одной сенсорной системе и получают афферентные импульсы от специфических проекционных ядер; неспецифические – большая группа ядер таламуса, которые функционально связаны с ретикулярной формацией ствола. К числу этих ядер относится медианная и внтурипластинчатая группы из ретикулярной формации и, кроме того, имеет двусторонние связи со специфическими ядрами таламуса. В отличие от специфических ядер с локальными проекциями в коре филогенетически более древние неспецифические ядра обнаруживают диффузные проекции во все области коры. Этой структурной особенностью обусловлены их название и функция, которая состоит в регуляции возбудимости и электрической активности корковых нейронов.
Морфофункциональная организация таламуса По функции ядра таламуса можно разделить на: специфические (проекционные) – синаптическое переключение сенсорной информации с аксонов восходящих афферентных путей на следующие, конечные нейроны, отростки которых идут в соответствующие сенсорные проекционные области коры больших полушарий; ассоциативные – в отличие от специфических ядер они не могут быть отнесены к какой-либо одной сенсорной системе и получают афферентные импульсы от специфических проекционных ядер; неспецифические – большая группа ядер таламуса, которые функционально связаны с ретикулярной формацией ствола. К числу этих ядер относится медианная и внтурипластинчатая группы из ретикулярной формации и, кроме того, имеет двусторонние связи со специфическими ядрами таламуса. В отличие от специфических ядер с локальными проекциями в коре филогенетически более древние неспецифические ядра обнаруживают диффузные проекции во все области коры. Этой структурной особенностью обусловлены их название и функция, которая состоит в регуляции возбудимости и электрической активности корковых нейронов.
 Схематическое изображение основных ядер гипоталамуса. I — передняя комиссура; 2 — преоптическое ядро; 3 — супраоптическое ядро; 4 — ножка гипофиза; 5 Схематическое изображение вентромедиальное гипоталамическос ядро; специфических и неспецифических 6 - дорсомедиальное гипоталамическое таламокортикальных проекций. ядро; 7 — медиальное ядро мамиллярного НП — неспецифические таламические пути; тела; 8 — мамиллярно-таламический пучок; СП — специфические таламические пути. 9 — заднее гипоталамическое ядро; 10 паравентрикулярное ядро.
Схематическое изображение основных ядер гипоталамуса. I — передняя комиссура; 2 — преоптическое ядро; 3 — супраоптическое ядро; 4 — ножка гипофиза; 5 Схематическое изображение вентромедиальное гипоталамическос ядро; специфических и неспецифических 6 - дорсомедиальное гипоталамическое таламокортикальных проекций. ядро; 7 — медиальное ядро мамиллярного НП — неспецифические таламические пути; тела; 8 — мамиллярно-таламический пучок; СП — специфические таламические пути. 9 — заднее гипоталамическое ядро; 10 паравентрикулярное ядро.
 и неспецифических (б) ядер таламуса в") Расположение основных ядерных групп гипоталамуса Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших полушарий 1 — сенсорные афферентные пути, 2 — ретикулярно-таламические пути, 3 — проекционная зона коры, 4 — ассоциативная зона коры. 1 — паравентрикулярное ядро, 2 — спайка, 3 — свод, 4 — мозолистое тело, 5 — таламус, 6 — шишковидное тело, 7 — водопровод, 8 — средний мозг, 9 — заднее ядро, 10 — сосцевидное тело, 11 — дорсомедиальное ядро, 12 — латеральное ядро, 13 — вентромедиальное ядро, 14 — нейрогипофиз, 15 — аденогипофиз, 16 — зрительный перекрест, 17 — супраоптическое ядро.
Расположение основных ядерных групп гипоталамуса Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших полушарий 1 — сенсорные афферентные пути, 2 — ретикулярно-таламические пути, 3 — проекционная зона коры, 4 — ассоциативная зона коры. 1 — паравентрикулярное ядро, 2 — спайка, 3 — свод, 4 — мозолистое тело, 5 — таламус, 6 — шишковидное тело, 7 — водопровод, 8 — средний мозг, 9 — заднее ядро, 10 — сосцевидное тело, 11 — дорсомедиальное ядро, 12 — латеральное ядро, 13 — вентромедиальное ядро, 14 — нейрогипофиз, 15 — аденогипофиз, 16 — зрительный перекрест, 17 — супраоптическое ядро.
, составляющее вместе с медиальным") Специфическим ядром зрительной сенсорной системы является латеральное коленчатое тело (ЛКТ), составляющее вместе с медиальным коленчатым телом метаталамус и имеющее прямые связи с затылочными (зрительными) проекционными областями коры больших полушарий. ЛКТ имеет слоистую структуру и организовано также по топическому принципу. Аксоны, идущие в ЛКТ из зрительного тракта, распределяются в нем с поразительной четкостью: три слоя ЛКТ связаны с ипсилатеральным (расположенном на той же стороне) глазом, а три остальные – с контрлатеральным. Рецептивное поле нейрона ЛКТ представляет собой концентрически организованную совокупность рецепторов сетчатки, имеющую либо возбуждающий (при включении света) центр и периферическую тормозную сеть, либо, напротив, тормозной центр и возбуждающую данный нейрон ЛКТ периферическую область.
Специфическим ядром зрительной сенсорной системы является латеральное коленчатое тело (ЛКТ), составляющее вместе с медиальным коленчатым телом метаталамус и имеющее прямые связи с затылочными (зрительными) проекционными областями коры больших полушарий. ЛКТ имеет слоистую структуру и организовано также по топическому принципу. Аксоны, идущие в ЛКТ из зрительного тракта, распределяются в нем с поразительной четкостью: три слоя ЛКТ связаны с ипсилатеральным (расположенном на той же стороне) глазом, а три остальные – с контрлатеральным. Рецептивное поле нейрона ЛКТ представляет собой концентрически организованную совокупность рецепторов сетчатки, имеющую либо возбуждающий (при включении света) центр и периферическую тормозную сеть, либо, напротив, тормозной центр и возбуждающую данный нейрон ЛКТ периферическую область.
 к бледному шару черной субстанции") Схематическое изображение топографический организации проекций нейронов полосатого тела (С) к бледному шару черной субстанции (ЧС). а — дорсовентральная и медиовентральная организация; б — вентродорсальная и медиолатеральная организация; ХЯ — хвостатое ядро; Схематическое изображение организации синаптических проекций полосатого тела
Схематическое изображение топографический организации проекций нейронов полосатого тела (С) к бледному шару черной субстанции (ЧС). а — дорсовентральная и медиовентральная организация; б — вентродорсальная и медиолатеральная организация; ХЯ — хвостатое ядро; Схематическое изображение организации синаптических проекций полосатого тела
 Афферентные и эфферентные связи базальных ядер 1 — хвостатое ядро, 2 — скорлупа, 3 — черное вещество, 4 —- таламус, 5, 5* — внутренний и внешний отделы бледного шара, 6 — субталамическое ядро, 7 — сенсомоторная кора, 8 — двигательная кора, 9 — пути к стволовой части мозга; стрелками обозначены афферентные пути, эфферентные пути, межъядерные взаимодействия.
Афферентные и эфферентные связи базальных ядер 1 — хвостатое ядро, 2 — скорлупа, 3 — черное вещество, 4 —- таламус, 5, 5* — внутренний и внешний отделы бледного шара, 6 — субталамическое ядро, 7 — сенсомоторная кора, 8 — двигательная кора, 9 — пути к стволовой части мозга; стрелками обозначены афферентные пути, эфферентные пути, межъядерные взаимодействия.
 Нарушение механизмов организации соматического рефлекса
Нарушение механизмов организации соматического рефлекса
, конвергенция (Б), окклюзия (В), временная (Г) и пространственная (Д) суммация") Общая организация ЦНС Дивергенция(А), конвергенция (Б), окклюзия (В), временная (Г) и пространственная (Д) суммация в локальных нейронных цепях. Реципрокное (А), пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга: 1 – мотонейрон, 2 – тормозный интернейрон, 3 – эфферентные терминали
Общая организация ЦНС Дивергенция(А), конвергенция (Б), окклюзия (В), временная (Г) и пространственная (Д) суммация в локальных нейронных цепях. Реципрокное (А), пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга: 1 – мотонейрон, 2 – тормозный интернейрон, 3 – эфферентные терминали

 и подпороговая «кайма» (пунктирная линия) популяции") А — кругами обозначены центральные зоны (сплошная линия) и подпороговая «кайма» (пунктирная линия) популяции нейронов. При изолированном раздражении волокон / и // возбуждаются нейроны только в центральных зонах. При одновременном раздражении волокон / и // в реакцию вовлекаются и нейроны подпороговой «каймы» , на которых конвергируют разветвления этих волокон (суммация); Б — кругами обозначены центральные зоны. При раздражении одного из волокон (/ или //) возбуждение возникает в 5 нейронах. При одновременной же стимуляции этих волокон потенциалы действия возникают не в 10, а в 8 нейронах (окклюзия).
А — кругами обозначены центральные зоны (сплошная линия) и подпороговая «кайма» (пунктирная линия) популяции нейронов. При изолированном раздражении волокон / и // возбуждаются нейроны только в центральных зонах. При одновременном раздражении волокон / и // в реакцию вовлекаются и нейроны подпороговой «каймы» , на которых конвергируют разветвления этих волокон (суммация); Б — кругами обозначены центральные зоны. При раздражении одного из волокон (/ или //) возбуждение возникает в 5 нейронах. При одновременной же стимуляции этих волокон потенциалы действия возникают не в 10, а в 8 нейронах (окклюзия).
 Пространственное облегчение – это превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Феномен объясняется суммацией ВПСП до КУД в группе мотонейронов, имеющих подпороговые ВПСП при раздельной активации входов. Окклюзия – это явление, противоположное пространственному облегчению. Здесь два сильных (т. е. реализуемых большим числом элементов) афферентных входа вместе возбуждают группу мотонейронов, меньшую той, которая образуется при арифметическом сложении ответов групп мотонейронов, возбуждаемых от этих входов, когда они действуют порознь. Посттетаническая потенциация – усиление рефлекторной реакции под влиянием предшествующих частых ритмических (так называемых тетанизирующих) раздражений.
Пространственное облегчение – это превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Феномен объясняется суммацией ВПСП до КУД в группе мотонейронов, имеющих подпороговые ВПСП при раздельной активации входов. Окклюзия – это явление, противоположное пространственному облегчению. Здесь два сильных (т. е. реализуемых большим числом элементов) афферентных входа вместе возбуждают группу мотонейронов, меньшую той, которая образуется при арифметическом сложении ответов групп мотонейронов, возбуждаемых от этих входов, когда они действуют порознь. Посттетаническая потенциация – усиление рефлекторной реакции под влиянием предшествующих частых ритмических (так называемых тетанизирующих) раздражений.
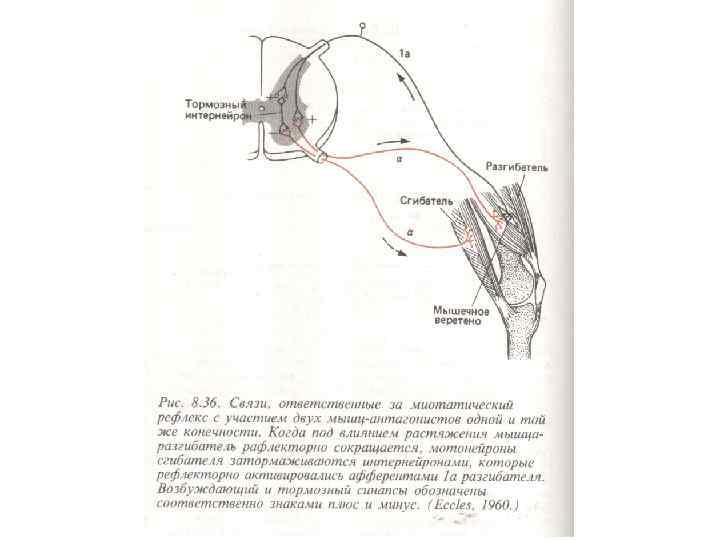
 торможение центров антагонистических рефлексов,") Торможение в нервных сетях: реципрокное торможение – это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов; возвратное торможение – это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам; латеральное торможение – это торможение элементов соседних нервных цепочек в конкурирующих сенсорных каналах связи; Латеральное торможение, видимо, также осуществляется с помощью тормозных вставочных клеток. Такое торможение четко показано у мечехвоста в сенсорных цепочках, идущих от фасеточного глаза к ЦНС. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах. прямое взаимное торможение – это тормозное взаимодействие двух (или большего числа) командных нейронов, осуществляющееся без специальных вставочных клеток;
Торможение в нервных сетях: реципрокное торможение – это взаимное (сопряженное) торможение центров антагонистических рефлексов, обеспечивающее координацию этих рефлексов; возвратное торможение – это торможение нейронов собственными импульсами, поступающими по возвратным коллатералям к тормозным клеткам; латеральное торможение – это торможение элементов соседних нервных цепочек в конкурирующих сенсорных каналах связи; Латеральное торможение, видимо, также осуществляется с помощью тормозных вставочных клеток. Такое торможение четко показано у мечехвоста в сенсорных цепочках, идущих от фасеточного глаза к ЦНС. Оно наблюдается у соседних элементов сетчатки позвоночных, а также в их зрительных, слуховых и других сенсорных центрах. прямое взаимное торможение – это тормозное взаимодействие двух (или большего числа) командных нейронов, осуществляющееся без специальных вставочных клеток;
 и в нервной системе ракообразных (Б): А") Реципрокное торможение в спинном мозгу позвоночных (Л) и в нервной системе ракообразных (Б): А — реципрокное торможение мотонейрона мышцы-разгибателя (осуществляется с помощью тормозного интернейрона); Б — реципрокное торможение мышцы-открывателя клешни рака при раздражении, возбуждающем закрыватель клешни (торможение направлено не на мотонейрон, а прямо на мышцу); 1 — мышца-разгибатель, 2 — мотонейрон разгибателя, 3 — тормозящий интернейрон, 4 — сенсорный нейрон, 5 — возбуждающий интернейрон, 6 — мотонейрон сгибателя, 7 — мышцасгибатель, 5 — тормоэный эффекторный нейрон, 9 — мышца-открыватель, 10 — мотонейрон; черным цветом обозначены тормозные клетки, красным — их нервные окончания
Реципрокное торможение в спинном мозгу позвоночных (Л) и в нервной системе ракообразных (Б): А — реципрокное торможение мотонейрона мышцы-разгибателя (осуществляется с помощью тормозного интернейрона); Б — реципрокное торможение мышцы-открывателя клешни рака при раздражении, возбуждающем закрыватель клешни (торможение направлено не на мотонейрон, а прямо на мышцу); 1 — мышца-разгибатель, 2 — мотонейрон разгибателя, 3 — тормозящий интернейрон, 4 — сенсорный нейрон, 5 — возбуждающий интернейрон, 6 — мотонейрон сгибателя, 7 — мышцасгибатель, 5 — тормоэный эффекторный нейрон, 9 — мышца-открыватель, 10 — мотонейрон; черным цветом обозначены тормозные клетки, красным — их нервные окончания
, латеральное (Б) и прямое взаимное торможение (В): А — в спинном мозгу") Возвратное (А), латеральное (Б) и прямое взаимное торможение (В): А — в спинном мозгу позвоночных; Б — в сенсорном пути; В — предполагаемое у командных клеток (моллюсков); в и т — возбуждающее и тормозное действие; стрелками показано направление нервного импульса Нервная цепочка, поддерживающая циркуляцию возбуждения (в) и постоянную выходную импульсацию (кольцевые связи нейронов в нервном центре по Лоренто де Но) Начало активности цепочки задается возбуждающим афферентным синапсом (1), конец — утомлением или внешним тормозным влиянием (т)
Возвратное (А), латеральное (Б) и прямое взаимное торможение (В): А — в спинном мозгу позвоночных; Б — в сенсорном пути; В — предполагаемое у командных клеток (моллюсков); в и т — возбуждающее и тормозное действие; стрелками показано направление нервного импульса Нервная цепочка, поддерживающая циркуляцию возбуждения (в) и постоянную выходную импульсацию (кольцевые связи нейронов в нервном центре по Лоренто де Но) Начало активности цепочки задается возбуждающим афферентным синапсом (1), конец — утомлением или внешним тормозным влиянием (т)


 НАЙТИ!") (дофаминэргические) НАЙТИ!
(дофаминэргические) НАЙТИ!


. На 6 -й") Так происходит образование амилоидной бляшки у генноинженерных мышей (показано белой стрелкой). На 6 -й день уже видна дистрофия нейрона (треугольная стрелка). Синий цвет — отложения амилоида, зеленый — нейроны; длина масштабной линейки 20 мкм; снимки сделаны с помощью мультифотонного микроскопа. Изображение из обсуждаемой статьи в Nature Так происходит образование амилоидной бляшки у генноинженерных мышей (показано белой стрелкой). На 6 -й день уже видна дистрофия нейрона (треугольная стрелка). Синий цвет — отложения амилоида, зеленый — нейроны; длина масштабной линейки 20 мкм; снимки сделаны с помощью мультифотонного микроскопа.
Так происходит образование амилоидной бляшки у генноинженерных мышей (показано белой стрелкой). На 6 -й день уже видна дистрофия нейрона (треугольная стрелка). Синий цвет — отложения амилоида, зеленый — нейроны; длина масштабной линейки 20 мкм; снимки сделаны с помощью мультифотонного микроскопа. Изображение из обсуждаемой статьи в Nature Так происходит образование амилоидной бляшки у генноинженерных мышей (показано белой стрелкой). На 6 -й день уже видна дистрофия нейрона (треугольная стрелка). Синий цвет — отложения амилоида, зеленый — нейроны; длина масштабной линейки 20 мкм; снимки сделаны с помощью мультифотонного микроскопа.
 Схема образования амилоидной бляшки у 5 -6 -месячных мышей. Day 1 — день, когда у мыши впервые замечено крохотное внеклеточное отложение амилоида — микробляшка. Затем отложение начинает стремительно расти, изменяются соседние нейроны и дендриты, к растущей бляшке стягиваются астроглия и микроглия. На 3 -й день уже заметно повреждение аксона. На 7 -й день бляшка достигает зрелости и ее структура стабилизируется.
Схема образования амилоидной бляшки у 5 -6 -месячных мышей. Day 1 — день, когда у мыши впервые замечено крохотное внеклеточное отложение амилоида — микробляшка. Затем отложение начинает стремительно расти, изменяются соседние нейроны и дендриты, к растущей бляшке стягиваются астроглия и микроглия. На 3 -й день уже заметно повреждение аксона. На 7 -й день бляшка достигает зрелости и ее структура стабилизируется.
 характеризуется: 1. Постепенным малозаметным началом в пресенильном или старческом возрасте. 2.") Болезнь Альцгеймера (БА) характеризуется: 1. Постепенным малозаметным началом в пресенильном или старческом возрасте. 2. Неуклонным прогрессированием расстройств памяти и высших корковых функций вплоть до тотального распада интеллекта и психической деятельности в целом. 3. Типичным набором нейропатологических признаков.
Болезнь Альцгеймера (БА) характеризуется: 1. Постепенным малозаметным началом в пресенильном или старческом возрасте. 2. Неуклонным прогрессированием расстройств памяти и высших корковых функций вплоть до тотального распада интеллекта и психической деятельности в целом. 3. Типичным набором нейропатологических признаков.
 Недавние исследования в области молекулярной генетики БА привели к идентификации трёх генов, ответственных за развитие наследственно-обусловленных форм заболевания: - На 21 -й хромосоме локализован ген амилоидного предшественника (АРР) - На 14 -й хромосоме – ген пресенилин-1 (PSN-1) - На 1 -й хромосоме – пресенилин-2 (PSN-2)
Недавние исследования в области молекулярной генетики БА привели к идентификации трёх генов, ответственных за развитие наследственно-обусловленных форм заболевания: - На 21 -й хромосоме локализован ген амилоидного предшественника (АРР) - На 14 -й хромосоме – ген пресенилин-1 (PSN-1) - На 1 -й хромосоме – пресенилин-2 (PSN-2)
, патогенетически более значимым процессом, вызывающим гибель") По мнению H. Braak и E. Braak (1996), патогенетически более значимым процессом, вызывающим гибель нейронов и развитие деменции, является не аномальный амилоидогенез, а накопление гиперфосфорилированного нерастворимого таупротеина, который составляет основу парноскрученных филаментов, образующих нейрофибрилярные клубки. Другим нейроморфологическим феном, который обнаруживает параллелизм с прогрессированием когнитивного снижения, является уменьшение числа синапсов в лобной и височной коре и в гиппокампе. Предположение, что развитие деменции при БА прямо связано с утратой синаптических контактов в специфических корковых и подкорковых областях мозга (1995).
По мнению H. Braak и E. Braak (1996), патогенетически более значимым процессом, вызывающим гибель нейронов и развитие деменции, является не аномальный амилоидогенез, а накопление гиперфосфорилированного нерастворимого таупротеина, который составляет основу парноскрученных филаментов, образующих нейрофибрилярные клубки. Другим нейроморфологическим феном, который обнаруживает параллелизм с прогрессированием когнитивного снижения, является уменьшение числа синапсов в лобной и височной коре и в гиппокампе. Предположение, что развитие деменции при БА прямо связано с утратой синаптических контактов в специфических корковых и подкорковых областях мозга (1995).
 К настоящему времени многочисленные нейрогистологические и нейрохимические исследования мозга больных с деменцией альцгеймеровского типа позволили установить несколько каскадов биологических событий: 1. Нарушение процессов фосфорилирования белков. 2. Изменения в метаболизме глюкозы. 3. Активация процессов перикисного окисления липидов. Полагают, что каждый из таких каскадов или их совокупность могут в конечном итоге приводить к описанным выше структурным изменениям, которые лежат в основе нейрональной дегенерации и сопровождаются развитием деменции.
К настоящему времени многочисленные нейрогистологические и нейрохимические исследования мозга больных с деменцией альцгеймеровского типа позволили установить несколько каскадов биологических событий: 1. Нарушение процессов фосфорилирования белков. 2. Изменения в метаболизме глюкозы. 3. Активация процессов перикисного окисления липидов. Полагают, что каждый из таких каскадов или их совокупность могут в конечном итоге приводить к описанным выше структурным изменениям, которые лежат в основе нейрональной дегенерации и сопровождаются развитием деменции.
 связаны с уменьшением размеров гиппокампа (жёлтый) и") Структурные изменения в мозге при шизофрении (сверху) связаны с уменьшением размеров гиппокампа (жёлтый) и увеличением объёма заполненных жидкостью желудочков (серый); для сравнения изображён мозг здорового человека (снизу). Получены с применением метода ЯМР.
Структурные изменения в мозге при шизофрении (сверху) связаны с уменьшением размеров гиппокампа (жёлтый) и увеличением объёма заполненных жидкостью желудочков (серый); для сравнения изображён мозг здорового человека (снизу). Получены с применением метода ЯМР.