Консультация бакалавров.ppt
- Количество слайдов: 53



Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году и приведено в соответствие с накопившимися к тому времени данными в 1970 году. Переход генетической информации от ДНК к РНК и от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул.

Организация растительной и животной клетки Рис. 1. Схема ультрамикроскопического строения клеток животных организмов

Рис. 2. Схема строения молодой эвкариотической клетки. Обозначения: в — вакуоли, д — диктиосомы, з. к — зерно крахмала, мж — межклетник, м. п — межклеточная пластинка, мтх — митохондрии, плз — плазмодесмы, пп — пропластиды, р — рибосомы, с. к — стенка клетки, сф — сферосома, э. с — эндоплазматическая сеть, я — ядро, я. п — пора в ядерной оболочке, яш — ядрышко

1. Строение растительной и животной клеток. Признаки сходства в строении этих клеток: наличие ядра, цитоплазмы, клеточной мембраны, митохондрий, рибосом, комплекса Гольджи и др. Признаки сходства — доказательство родства растений и животных. Отличия: только растительные клетки имеют твердую оболочку из клетчатки, пластиды, вакуоли с клеточным соком. 2. Функции клеточных структур. Функции оболочки и клеточной мембраны: защита клетки, поступление в нее одних веществ из окружающей среды и выделение других. Выполнение оболочкой функции скелета (постоянная форма клетки). Расположение цитоплазмы между клеточной мембраной и ядром, а в цитоплазме всех органоидов клетки. Функции цитоплазмы: связь между ядром и органоидами клетки, осуществление всех процессов клеточного обмена веществ (кроме синтеза нуклеиновых кислот), расположение в ядре хромосом, в которых хранится наследственная информация о признаках организма, передача хромосом от родителей потомству в результате деления клеток. Роль ядра в управлении синтезом белка клетки и всеми физиологическими процессами. Окисление в митохондриях органических веществ кислородом с освобождением энергии. Синтез в рибосомах молекул белка. Наличие хлоропластов (пластид) в растительных клетках, образование в них органических веществ из неорганических с использованием солнечной энергии (фотосинтез). 3. Жизнедеятельность клетки. Питание, дыхание. Рост. Деление (размножение) клеток. Создание клеточной структуры в процессе питания из органических веществ. Сущность дыхания: окисление органических веществ клетки и освобождение энергии, которая используется в процессах жизнедеятельности. Рост молодых клеток и их старение. Размножение клетки путем деления.
 стенка из целлюлозы, у")
1. В растительной клетке присутствует прочная и толстая клеточная (жесткая) стенка из целлюлозы, у животной - она эластичная. 2. В растительной клетке развита сеть вакуолей, иногда одна большая вакуоль; в животной клетке она развита слабо. 3. Растительная клетка содержит особые органоиды – пластиды (а именно, хлоропласты, лейкопласты и хромопласты), а животная клетка их не содержит. 4. В растительной клетке отсутствие клеточного центра (клеточный центр есть только у низших растений). 5. У растительной клетки питание автотрофное, а у животной гетеротрофное.

Из цитологических особенностей важное значение имеет наличие у растений твердой углеводной оболочки — продукта жизнедеятельности протопласта. Между оболочками соседних клеток находится так называемая клеточная, или межклеточная пластинка, состоящая из цементирующих клетки пектиновых веществ. Клетки животных, как правило, не имеют оболочек, их поверхностные мембраны — плазмалеммы — непосредственно контактируют с другими клетками или с внешней средой. Однако различия между растениями и животными по этому признаку не абсолютны, так как некоторые клетки животных имеют образования, аналогичные клеточным оболочкам растений. Внутренний слой плазмалеммы состоит из белков, а наружный — практически у всех клеток животных представлен гликокаликсом — гидрофильным слоем, состоящим из полисахаридов, связанных с белками. Он играет роль соединительного слоя между плазмалеммами соседних клеток и аналогичен клеточной пластинке растений. Гликокаликс иногда развит настолько сильно, что образует вокруг плазмалеммы волокнистый чехол, имитирующий оболочку растительной клетки. Примером таких клеток могут служить клетки эпителия кишечника. С другой стороны, не все клетки растений имеют оболочки. Зооспоры некоторых водорослей в процессе развития их теряют.

Первым, кто обратил внимание на возникновение органов из зародышевых листков, или пластов, был К. Ф. Вольф. Изучая развитие цыпленка, он показал, что из «неорганизованной, бесструктурной» массы яйца возникают зародышевые листки , дающие затем начало отдельным органам. К. Ф. Вольф различал нервный и кишечный листки, из которых развиваются соответствующие органы. Впоследствии X. Пандер, последователь К. ФВольфа, также описал наличие у куриного эмбриона зародышевых листков. К. М. Бэр обнаружил наличие зародышевых листков и у других животных, в связи с чем распространил понятие о зародышевых листках на всех позвоночных. Так, К. М. Бэр различал первичные зародышевые листки, называя их анимальным и вегетативным, из которых впоследствии, в процессе эмбрионального развития, возникают вторичные зародышевые листки, дающие начало определенным органам. Описание зародышевых листков значительно облегчило изучение особенностей эмбрионального развития организмов и дало возможность установить филогенетические связи между животными, казалось весьма отдаленными в систематическом отношении. Это было блестяще продемонстрировано А. О. Ковалевским, который по праву считается основателем современной теории зародышевых листков. А. О. Ковалевский на основании широких сравнительно-эмбриологических сопоставлений показал, что двуслойную стадию развития проходят почти все многоклеточные организмы. Он доказал сходство зародышевых листков у различных животных не только по происхождению, но и по производным зародышевых листков.
 повторяет в")
Биогенетический закон Мюллера-Геккеля - каждое живое существо в своем индивидуальном развитии (онтогенез) повторяет в известной степени формы, пройденные его предками или его видом (филогенез). «Биогенетический закон» как следствие эволюционного развития организмов впервые был сформулирован (довольно нечётко) английским естествоиспытателем Чарльзом Дарвином в его книге «Происхождение видов» в 1859 г: «Интерес эмбриологии значительно повысится, если мы будем видеть в зародыше более или менее затененный образ общего прародителя, во взрослом или личиностном его состоянии, всех членов одного и того же большого класса» . За 2 года до формулировки Эрнстом Геккелем биогенетического закона сходную формулировку предложил на основе своих исследований развития ракообразных работавший в Бразилии немецкий зоолог Фриц Мюллер. В своей книге «За Дарвина» (Für Darwin), изданной в 1864 году, он выделяет курсивом мысль: «историческое развитие вида будет отражаться в истории его индивидуального развития» . Краткая афористичная формулировка этого закона была дана немецким естествоиспытателем Эрнстом Геккелем в 1866 г. Краткая формулировка закона звучит следующим образом: Онтогенез есть рекапитуляция филогенеза (во многих переводах — «Онтогенез есть быстрое и краткое повторение филогенеза» ).

Один из выдающихся советских морфологов.

Из эктодермы развиваются: Эпидермис кожи и его производные (волосы, ногти, перья, сальные, потовы и молочные железы), компоненты органов зрения (хрусталик и роговица), слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшими эктодермальными производными являются нервная трубка, нервный гребень и образующиеся из них все нервные клетки. Органы чувств, передающие нервной системе информацию о зрительных, звуковых, обонятельных и иных стимулах, также развиваются из эктодермальных закладок. Например, сетчатка глаза образуется как вырост мозга и, следовательно, является производным нервной трубки, тогда как обонятельные клетки дифференцируются прямо из эктодермального эпителия носовой полости. Производными энтодермы являются: Эпителий желудка и кишки, клетки печени, секреторные клетки поджелудочной железы, слюнных, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секреторные клетки передней и средней доли гипофиза, щитовидной и паращитовидной желез. Из мезодермы формируются: Cкелет, скелетная мускулатура, соединительно-тканная основа кожи (дерма), органы выделительной и половой систем, сердечно-сосудистая система, лимфатическая система, плевра, брюшина и перикард.
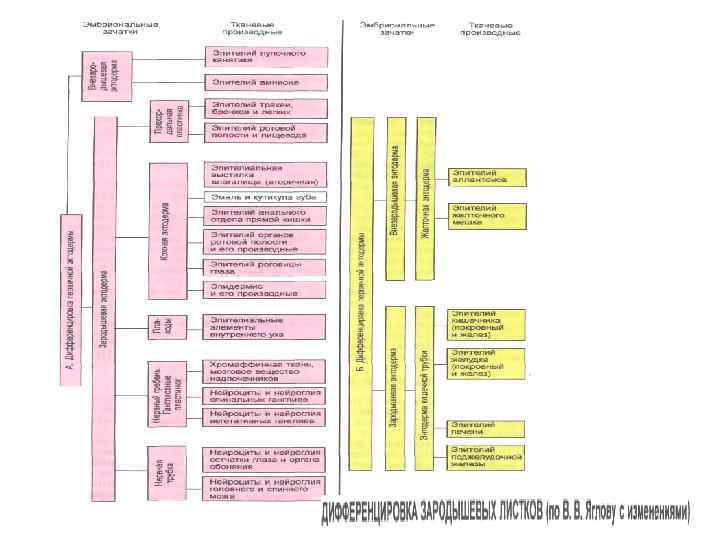
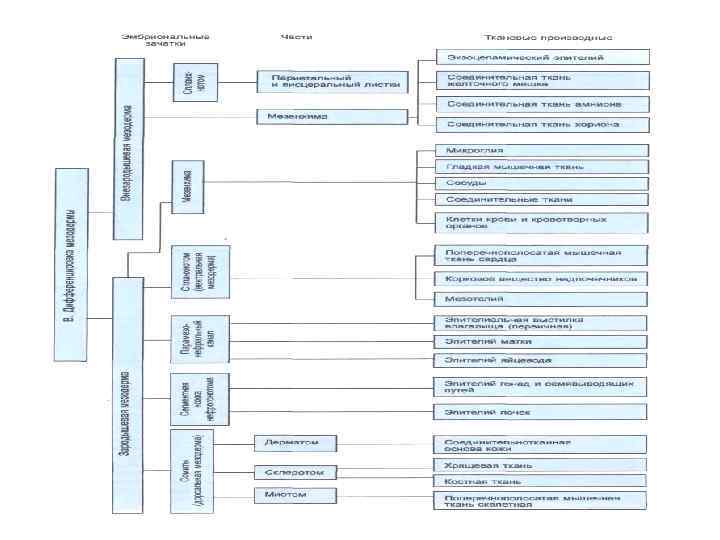


Рис. Схема строения клеточной мембраны
; 2 б")
2 – молекула липида: 2 а – гидрофобная часть (углеводные «хвосты» ); 2 б – гидрофильная часть Р- поверхность (от англ. рrotoplasmic) – протоплазматическая, ближайшая к цитоплазме; Е – поверхность (от англ. еxternal) - наружная



")
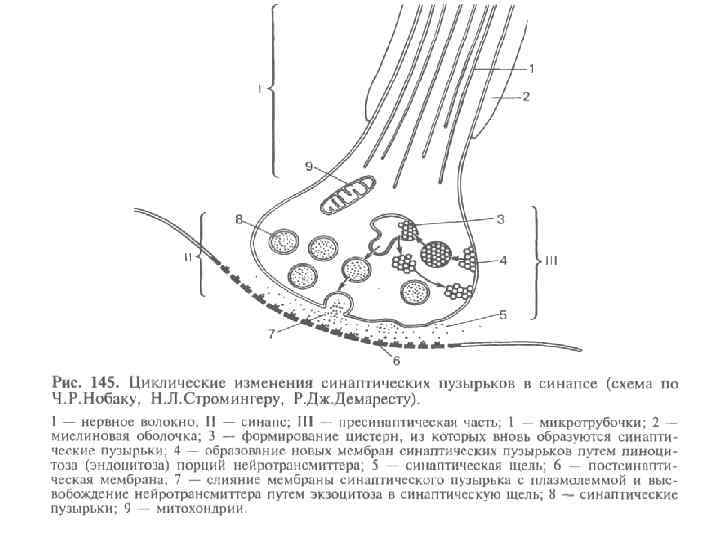
Рис. Строение синапса (схема)


ЖИЗНЕННЫЙ ЦИКЛ КЛЕТКИ Клеточный цикл - это совокупность явлений, или процессов между двумя последовательными делениями клетки, или между ее образованием и гибелью. Клеточный цикл состоит из митоза и интерфазы. Интерфаза значительно более длительна, чем митоз (обычно занимает не менее 90 % всего времени клеточного цикла) и подразделяется на три периода: • пресинтетический, или постмитотический - G 1; • синтетический - S; • постсинтетический, или премитотический G 2. ИНТЕРФАЗА = G 1 + S + G 2 КЛЕТОЧНЫЙ ЦИКЛ = М (митоз) + интерфаза 1. Пресинтетический, или постмитотический - G 1 период наступает сразу же после митотического деления клетки. Это самый продолжительный период, следует сразу за телофазой митоза. Продолжительность этого периода - от нескольких часов до нескольких дней. Длительность зависит от типа и расположения клеток, митотической активности. У быстро делящихся клеток (эмбриональные) этот период непродолжительный. Продолжительность периода G 1 у клеток кишечника - 4, 5 часа, у клеток роговицы 70 часов, клеток эпителия уха примерно 528 часов. В этот период происходит активный рост клетки (идет восстановление нормальных размеров клетки, восстановление клеточной массы, органелл), синтез белка и РНК. G 1 - один из основных периодов клеточного цикла. В этот период (или в эту фазу) синтезируются особые "запускающие" белки, или активаторы S-периода. Но не у всех клеток клеточный цикл одинаков. Условно в фазе G 1 выделяют точку R - точка рестрикции, или ограничения. Если в G 1 периоде синтезируется достаточно большое количество белков - активаторов, то клетка в своем развитии достигает точки R, при достижении этой точки клетка переходит в S -период.

Контроль, осуществляемый на уровне точки R ограничивает возможность нерегулируемого размножения клеток. Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G 0). У многих клеток это запланировано. Характерно для высокоспециализированных клеток (нервные клетки, эритроциты, кардиомиоциты). Клетка переходит в период для того, чтобы (в зависимости от причин остановки): • • • дифференцироваться и выполнять свои специфические функции; выжить в условиях недостаточности питательных веществ или факторов роста; осуществить репарацию поврежденной ДНК. Клетки одних тканей при соответствующей стимуляции вновь способны возвращаться из периода G 0 в клеточный цикл, либо может перейти в состояние D (смерть). 2. Синтетический - S - период. Из G 1 (при достижении точки R) клетка переходит в S период. Длительность этого периода в среднем составляет 8 - 12 часов. В этот период происходит репликация (удвоение) ДНК, удваивается число центриолей. Продолжается синтез белка, в частности, гистонов, которые поступают в ядро из цитоплазмы и обеспечивают нуклеосомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. 3. Когда заканчивается удвоение ДНК и центриолей наступает фаза, или период постсинтетический, или премитотический G 2, который длится 2 - 4 часа, вплоть до митоза (М). В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запас энергии (накапливается АТФ), синтезируются РНК и белки (в частности, тубулин), необходимые для процесса деления.

Для всех типов клеток суммарная длительность S + G 2 периодов имеет относительное постоянство. Далее наступает стадия митоза (М), который включает 4 основных фазы: профазу, метафазу, анафазу и телофазу. Митоз длится в среднем 1 - 3 часа (в зависимости от типа клетки) и обеспечивает равномерное распределение генетического материала в дочерние клетки. РЕГУЛЯЦИЯ КЛЕТОЧНОГО ЦИКЛА По интенсивности деления клеток (или по уровню обновления клеток) выделяют три типа клеточных популяций: Величина клеточного цикла подвержена регуляции, т. е. есть факторы, регулирующие клеточный цикл: усиливающие и тормозящие. стабильные клеточные популяции состоят из клеток, потерявших способность к делению (нейроны, кардиомиоциты). Величина клеточной популяции формируется в начале дифференцировки. По мере старения их число снижается вследствие невосполня емой естественной убыли клеток. растущие клеточные популяции специализированные клетки, которые обычно не делятся, но сохраняют эту способность. Это, как правило, долгоживущие клетки, но способные к делению под действием раздражителя (клетки почек, печени, поджелудочной и щитовидной желез). обновляющие клеточные популяции которые постоянно делятся, т. е. характеризуются постоянным обновлением клеток (эпителий кишки и эпидермис, клетки костного мозга, клетки крови и т. д. ).

УСИЛИВАЮЩИЕ: • Протоонкогены (группа генов - активаторов, контролирующих нормальное клеточное деление и дифференцировку. Продукты экспрессии этих генов (особые белки) оказывают "+" влияние на разные механизмы регуляции клеточного деления). К настоящему времени идентифицировано около 50 разновидностей протоонкогенов. Побочное действие - когда их много, то причина развития раковых заболеваний (установлено, что изменения структуры и усиление активности экспрессии протоонкогенов вызывает развитие опухолей - что определило их название). • Специфические белки, имеющие тканевую специфичность - белки - факторы роста. Представляют собой белки, усиливающие митотическую активность в определенных тканях (тканях - мишенях). Их действие опосредуется специфическими рецепторами на плазмолемме клеток. к ним относятся: эпидермальный фактор роста (ЭФР), тромбоцитарный фактор роста (ТРФР), инсулиноподобные факторы роста (ИФР), фактор роста фибробластов (ФРФ) и т. д. ТОРМОЗЯЩИЕ: • Антионкогены - гены, продукты которых - супрессоры опухолевого роста - угнетают митотическую активность клеток • Кейлоны представляют собой класс гормоноподобных регуляторов, угнетающих клеточное размножение. Они являются полипептидами или гликопротеинами и обладают тканевой и клеточной специфичностью. Кейлоны образуются всеми зрелыми дифференцированными клетками и локально воздействуют на незрелые клетки этой же ткани, способные к делению. Они обеспечивают гомеостаз численности клеточной популяции, а их выделение контролируется механизмом отрицательной обратной связи. Уменьшение численности популяции клеток (например, потеря клеток эпидермиса при ранении или лейкоцитов при кровотечении) вызывает снижение ингибирующего воздействия кейлонов и подъем митотической активности в соответствующей ткани. Кейлоны участвуют в регуляции роста тканей, заживлении ран, иммунных реакций и других процессов.
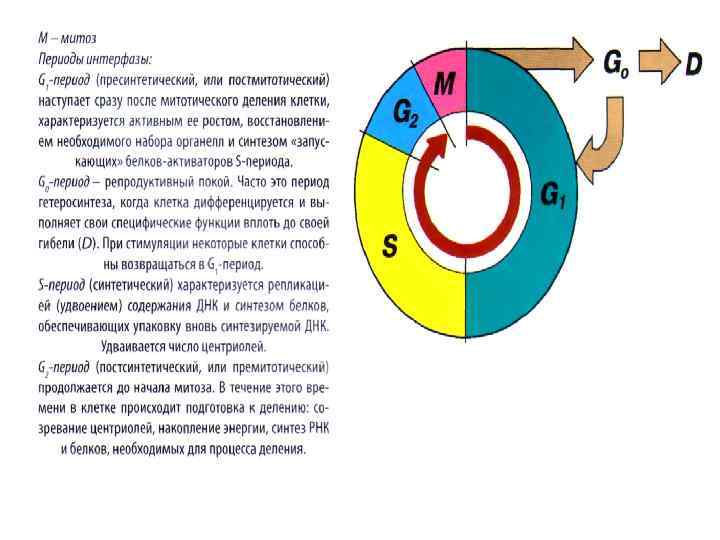

Рис. Митохондрия

В некоторых клетках встречаются кристы в виде трубочек и пузырьков– тубулярно -везикулярные кристы. Характерны для клеток, синтезирующих стероидные гормоны (клетки коркового вещества надпочечников, фолликулярные клетки и клетки желтого тела яичника, клетки Лейдига яичка). В таких клетках ферменты стероидогенезап локализуются частично в а. ЭПС, а частично – на внутренней митохондриальной мембране. В ходе синтеза стероидов промежуточные продукты неоднократно перемещаются между этими органеллами).

Кристы – складки внутренней мембраны толщиной 20 нм; располагаются чаще всего перпендикулярно длине митохондрии, но могут лежать и продольно. Форма крист в митохондриях большинства клеток – пластинчатая (ламеллярная)

В некоторых клетках встречаются кристы в виде трубочек и пузырьков– тубулярно-везикулярные кристы. Характерны для клеток, синтезирующих стероидные гормоны (клетки коркового вещества надпочечников, фолликулярные клетки и клетки желтого тела яичника, клетки Лейдига яичка). В таких клетках ферменты стероидогенезап локализуются частично в а. ЭПС, а частично – на внутренней митохондриальной мембране. В ходе синтеза стероидов промежуточные продукты неоднократно перемещаются между этими органеллами).






")
Рис. Стадии митоза (схема)

Соединительная ткань. Рыхлая волокнистая соединительная ткань Эта ткань является самой распространенным видом соединительных тканей и характеризуется сравнительно невысоким содержанием волокон в межклеточном веществе, относительно большим объемом основного аморфного вещества, многочисленным и разнообразным клеточным составом. Эта ткань обнаруживается повсеместно, во всех органах: • она образует их строму (основу) (в частности, междольковые прослойки и прослойки между слоями и оболочками); • заполняет пространства между функциональными элементами других тканей; • сопровождает нервы и сосуды; • входит в состав кожи и слизистых оболочек. Эта ткань является переходной между жидкими тканями и твердыми соединениями. В строении обнаруживаются признаки тех и других тканей. ФУНКЦИИ ТКАНИ: • Трофическая. • Регуляторная (регулирует обмен веществ). • Опорная (механическая) • Защитная [1) функция паралона и защита от механических повреждений; 2) клетки, которые фагоцитируют вредные вещества]. Как уже отмечалось выше - характерным признаком этой ткани является разнообразие клеточного состава.

Клетки рыхлой волокнистой соединительной ткани представляют собой сложную гетерогенную популяцию функционально разнообразных и взаимодействующих между собой и с компонентами межклеточного вещества элементов, которые условно объединяют в несколько групп. По признаку постоянства присутствия в составе рыхлой волокнистой соединительной ткани ее клетки подразделяют на: • Оседлые (фиксированные, резидентные) клетки, т. е. образуются и постоянно пребывающие в этой ткани. К этой группе относят фибробласты, адвентициальные клетки, фиброциты и жировые (адипоциты). В зрелой рыхлой волокнистой соединительной ткани содержанеие оседлых клеток относительно стабильно. • Блуждающие клетки (иммигранты) - подвижные клетки, поступающие в соединительную ткань из крови. В эту группу включают все виды лейкоцитов (гранулоцитов и агранулоцитов). Содержание этих клеток в отдельных участках соединительной ткани может существенного изменяться при различных иммунных реакциях и воспалении. По источникам развития выделяют три группы клеток: • Клетки линии механоцитов - адвентициальные клетки, фибробласты, фиброциты, адипоциты - развиваются из особой стволовой клетки этой клеточной линии, которая имеет мезенхимное происхождение. К линии механоцитов относят также клетки других тканей ретикулярной (ретикулярные клетки), скелетных тканей (хондроциты и остеоциты), поскольку вырабатываемые ими продукты (компоненты межклеточного вещества) обеспечивают механические свойства тканей. • Клетки - потомки стволовой клетки крови (СКК) - макрофаги (гистиоциты), плазматические и тучные клетки, лейкоциты (гранулоциты и агранулоциты) - развиваются, как следует из названия группы, из СКК, которая происходит из мезенхимы. • Клетки нейрального происхождения - пигментные клетки (развиваются из предшественников, которые выселяются из нервного гребня).

ФИБРОБЛАСТЫ - наиболее распространенные и функционально ведущие клетки рыхлой волокнистой соединительной ткани, относящиеся к клеточной линии механоцитов. Функции фибробластов: • Продукция всех компонентов межклеточного вещества (волокон и основного аморфного вещества). • Поддержание структурной организации и химического гомеостаза межклеточного вещества (за счет сбалансированных процессов его выработки и разрушения). • Регуляция деятельности других клеток соединительных тканей и влияние на другие ткани. Развитие фибробластов: стволовая клетка линии механоцитов → полустволовая клеткапредшественник → малодифференцированный (юный) фибробласт → зрелый (дифференцированный) фибробласт → фиброцит. Фибробласты - это крупные отросчатые клетки с нерезкими границами и светлым ядром, содержащим мелкодисперсный хроматин и 1 - 2 ядрышка. Цитоплазма слабо базофильна и характеризуется диплазматической дифференцировкой - нерезким разделением на внутреннюю, более плотную часть, окружающую ядро, - эндоплазму и периферическую, сравнительно светлую и образующую отростки - эктоплазму. Эндоплазма содержит большую часть органелл мощно развитого синтетического аппарата, а эктоплазма заполнена преимущественно элементами цитоскелета. Цистерны гр. ЭР часто растянуты, содержат мелкозернистый материал низкой электронной плотности. В цитоплазме располагаются также лизосомы, митохондрии, липидные капли и многочисленные пузырьки. Все элементы цитоскелета хорошо выражены. Фибробласт обладает незначительной способностью к движению, изменять свою форму.

Большинство фибробластов разрушается в процессе жизнедеятельности, но часть их превращается в малоактивнуюю долгоживущую форму - фиброциты. Фиброциты - конечная форма развития фибробласта - узкая веретенообразная, неспособная к пролиферации клетка с длинными тонкими отростками, которые часто имеют уплощенную крыловидную форму. Функция этих клеток состоит в регуляции метаболизма и поддержании стабильности межклеточного вещества, синтез его компонентов осуществляется ими очень слабо. Фиброциты располагаются между пучками коллагеновых волокон. Макрофаги (гистиоциты) - вторые по численности (после фибробластов) клетки рахлой волокнистой соединительной ткани. В этих клетках развит АГ, гр. ЭР, митохондрии, большое количество лизосом. Они могут пребывать в одном из двух взаимообратных состояний: • покоящихся клеток, обладающих низкой функциональной активностью; • 2) блуждающих клеток с высокой функциональной активностью. ФУНКЦИИ ГИСТИОЦИТОВ: • Защитная - распознование, поглощение и переваривание поврежденных, зараженных, опухолевых и погибших клеток, компонетов межклеточного вещества, а также экзогенных материалов и микрорганизмов. • Участие в индукции иммунных реакций посредством захвата, переработки (процессинга) антигенов и представления их лимфоцитам (играют роль антигенпредставляющих клеток). • Регуляция деятельности клеток других типов (фибробластов, лимфоцитов, тучных клеток, эндотелиоцитов и др. ).

Перициты - имеют отросчатую форму и в виде корзинки окружают кровеносные сосуды, это клетки небольшого диаметра, плотно прилегают к базальной мембране и их отростки контактируют через поры с эндотелиальными клетками. Благодаря этому контакту: • участвуют в транспорте веществ через стенку сосудов; • отвечают за поддержание формы кровеносных сосудов. Имеют сократительные элементы, которые регулируют просвет сосудов; • способны регулировать размножение эндотелиоцитов. Адвентициальные клетки - малоспециализированные клетки, сопровождающие мелкие сосуды. Имеют уплощенную или веретенообразную форму. Участвуют в формировании внешней соединительнотканной оболочки кровеносных и лимфатических сосудов. Тучные клетки имеют удлиненную или округлую форму. Они в 1, 5 - 2 раза крупнее базофилов. Цитоплазма тучных клеток содержит умеренно развитые органеллы, элементы цитоскелета, липидные капли и гранулы. Содержимое гранул тучных клеток: гепарин, гистамин, дофамин. ФУНКЦИИ ТУЧНЫХ КЛЕТОК: • Гомеостатическая, которая осуществляется в физиологических условиях путем медленного выделения небольших количеств биологически активных веществ, способных влиять на различные тканевые функции - в первую очередь, на проницаемость и тонус сосудов и поддержание баланса жидкостей в тканях. • Защитная и регуляторная, которая обеспечивается путем локального выделения медиаторов воспаления и хемотаксических факторов, обеспечивающих: а) мобилизацию эозинофилов и различных эффекторных клеток, участвующих в так называемых реакциях поздней фазы; б) воздействие на рост и созревание соединительной ткани в зоне воспаления. • Участие в развитии аллергических реакций вследствие наличия высокоаффинных рецепторов к иммуноглобулинам класса Е (Ig Е) на их плазмолемме и функциональной связи этих рецепторов с секреторным механизмом.

Плазмоциты - имеют мелкие размеры, располагаются поодиночке или группами и обладают высокой синтетической и секреторной активностью, вырабатывая и выделяя антитела (иммуноглобулины) и обеспечивая тем самым гуморальный иммунитет. Лейкоциты (гранулоциты и агранулоциты) являются нормальными компонентами рыхлой волокнистой соединительной ткани, в (или через) которую они мигрируют для выполнения своих функций после выхода из кровеносного русла. Лимфоциты, в отличие от других видов лейкоцитов, способны из соединительной ткани, в отличие от других лейкоцитов, способны из соединительной ткани через оттекающую лимфу вновь попадать в кровь (осуществлять рециркуляцию). Жировые клетки (адипоциты) - располагаются, как правило, около кровеносных сосудов. Накапливаясь в больших количествах, эти клетки образуют жировую ткань. В них запас энергии. В организме жировая ткань играет и механическую роль, образуя в некоторых органах подстилки (например, в коже). Ретикулоциты - клетки, имеющие звездчатое очертание, от тела клетки отходят различной длины и толщины отростки цитоплазмы, которые соединяются с такими же отростками соседних клеток. Они концентрируются по ходу кровеносных капилляров. При воспалении способны терять отростки и заниматься фагоцитозом. А также способны синтезировать межклеточное вещество (коллагеновые волокна III типа). Пигментные клетки имеют нейральное происхождение. Пигментные клетки имеют отросчатую форму и подразделяются на два вида - меланоциты, которые вырабатывают пигмент и меланофоры, способные накапливать его в цитоплазме Численное преобладание пигментных клеток над другими клеточными элементами соединительной ткани характерно для радужки и сосудистой оболочки глаза, где им принадлежит функционально ведущая роль.
.")
Межклеточное вещество рыхлой волокнистой соединительной ткани Состоит из волокон и основного аморфного вещества (матрикс). Оно является продуктом деятельности этих клеток этой ткани, в первую очередь, фибробластов. Матрикс - гелеобразное вещество, основу которого составляют гликозаминогликаны и сульфатированные гликозаминогликаны, обычно в комплексе с белками. ФУНКЦИИ МЕЖКЛЕТОЧНОГО ВЕЩЕСТВА: • Обеспечение архитектоники, физико-химических и механических свойств ткани. 2. Участие в создании оптимального микроокружения для деятельности клеток. • Объединение в единую систему всех клеток соединительной ткани и обеспечение передачи информации между ними. • Воздействие на многочисленные функции различных клеток (пролиферацию, дифференцировку, подвижность, синтетическую и секреторную активность, чувствительность к действию различных стимулирующих, ингибирующих (тормозящих) и повреждающих факторов и т. п. ). Этот эффект может осуществляться путем контактного взаимодействия компонентов межклеточного вещества и клеток, а также благодаря его способности накапливать и выделять факторы роста. Волокна, входящие в состав межклеточного вещества рыхлой волокнистой соединительной ткани, коллагеновые и эластические. КОЛЛАГЕНОВЫЕ ВОЛОКНА. Образованы белками коллагенами, которые получили свое название из-за способности содержащих их тканей при длительном вываривании давать животный клей. До настоящего времени обнаружено 19 видов (обозначаемых римскими цифрами), но наиболее изученные разберем позднее.

Основные функции коллагеновых волокон: 1. Обеспечение высоких механических свойств соединительной ткани. Чем выше содержание коллагеновых волокон в данной ткани, тем большей прочностью она обладает. Эти волокна практически не растяжимы; при увеличении нагрузки они лишь слегка распрямляются. • Определение (в значительной мере) архитектоники соединительной ткани. • Обеспечение взаимодействия между клетками и межклеточным веществом, а также связь между отдельными компонентами межклеточного вещества. • Влияние на пролиферацию, дифференцировку, миграцию и функциональную активность различных клеток. Коллагеновые волокна располагаются беспорядочно (в разных направлениях). Коллагены I, III, V типов наз. фибриллярными, или интерстициальными, т. к. они образуют фибриллы, которые входят в состав соединительных тканей. Коллаген IV, VII, IХ типа относят к аморфным (образует плоские сети). Распределение основных типов коллагена в организме Типы коллагена I Основные участки распределения в организме Соединительнотканная часть кожи (дерма), кость, волокнистый хрящ, дентин, связки, сухожилия, роговица глаза, рыхлая волокнистая соединительная ткань в различных органах. II Гиалиновый, эластический и (частично) волокнистый хрящ, стекловидное тело, хорда (эмбриона). III Ретикулярные волокна в кроветворных тканях, в стенке крупных кровеносных сосудов, кишке, печени, легком, клапанах сердца, гладкомышечной ткани, нервах. IV Базальные мембраны, капсула хрусталика V Базальные мембраны, стенка кровеносных сосудов, кожа, связки, дентин, роговица, гладкая и поперечно-полосатая мышечная ткани. VII Прикрепление базальной мембраны к близлежащим клеткам. IХ В состав матрикса эмбриона птиц

ЭЛАСТИЧЕСКИЕ ВОЛОКНА имеют меньший диаметр, чем у коллагеновых. Для них характерна прямолинейная направленность. Эластин - главный белковый компонент эластических волокон. Он составляет более 90 % их массы и представлен гликопротеиновыми молекулами, имеющих в состоянии покоя форму скрученных нитей. При растяжении они рапрямляются, а после прекращения действия нагрузки, вновь закручиваются. ФУНКЦИИ ЭЛАСТИЧЕСКИХ ВОЛОКОН: • Определение архитектоники ткани. • Обеспечение способности ткани к обратимой деформации и возвращению к исходной форме после ее временного изменения. ПЛОТНАЯ ВОЛОКНИСТАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ Образована теми же компонентами, что и рыхлая волокнистая соединительная ткань, отличаясь от нее: • очень высоким содержанием волокон (преимущественно коллагеновых); • малым количеством основного аморфного вещества в составе межклеточного вещества; • сравнительно низким содержанием межклеточных элементов; • преобладанием одного (основного) типа клеток - фиброцитов - над остальными (особенно плотной офоромленной ткани). Главное свойство плотной волокнистой соединительной ткани - очень высокая механическая прочность - обусловлено присутствием мощных пучков коллагеновых волокон. Ориентация этих волокон соответствует направлению действия сил, вызывающих деформацию ткани.

Межклеточное вещество: матрикс и коллагеновые, эластические волокна. Матрикс - в основном гликозаминогликаны. Клеточный состав: • Фиброциты • Фибробласты • Гистиоциты • Лейкоциты • Тучные клетки. В зависимости от расположения пучков коллагеновых волокон плотную волокнистую соединительную ткань подразделяют на: ОФОРМЛЕННУЮ Содержит толстые пучки коллагеновых волокон, располагающиеся параллельно другу (в направлении действия нагрузки), которые связаны небольшим количеством аморфного вещества. Между ними можно выявить сети эластических волокон. Содержание клеток невелико, среди них подавляющее большинство составляют фиброциты. Такая ткань образует сухожилия, связки, фасции и апоневрозы. НЕОФОРМЛЕННУЮ Характеризуется неупорядоченным расположением пучков коллагеновых волокон в трех различных плоскостях, которые переплетаются между собой, формируя трехмерную сеть, которая обеспечивает прочность ткани при воздействии деформирующих сил любой направленности. Имеются также эластические волокна, также формирующие трехмерную сеть. Содержание основного аморфного вещества невелико, клетки немногочисленны. Среди клеток преобладают фиброциты и фибробласты, но встречаются тучные клетки, гистиоциты, лейкоциты. Такая ткань образует глубокий (сетчатый) слой дермы (соединительнотканной части кожи), капсулы различных органов. Ткань, образующая капсулы, отличается более упорядоченным расположением коллагеновых волокон (преимущественно параллельно поверхнос-и органа), чем в сетчатом слое дермы.

СОЕДИНИТЕЛЬНЫЕ ТКАНИ СО СПЕЦИАЛЬНЫМИ СВОЙСТВАМИ Включают жировую, ретикулярную, слизистую и пигментную ткани. Они выполняют ряд важнейших специализированных функций и (за исключением широко распространенной в организме жировой ткани) характеризуются строго определенной топографией. Жировая ткань. Представляет собой особую разновидность соединительной ткани со специальными свойствами. Основной объем этой ткани занимают жировые клетки - адипоциты. Эта ткань повсеместно распространена в организме и составляет в норме около 15 - 20 % массы тела у мужчин и порядка 20 - 25 % - у женщин. Абсолютная масса жировой ткани (10 - 20 кг у здорового человека) спосорбна резко изменяться при патологических состояниях. При ожирении (которым страдает в развитых странах не менее 30 % взрослого населения) она увеличивается до 40 - 100 кг и более, при голодании или нервной анорексии (потере аппетита) - может снижаться до 3 % нормального уровня. Аномалии содержания и распределения жировой ткани связаны с рядом генетических нарушений и эндокринных растройств и нередко служат диагностически важными признаками заболеваний. ФУНКЦИИ ЖИРОВОЙ ТКАНИ: Энергетическая (трофическая) - благодаря накоплению липидов, служащих в организме резервными источниками энергии (легче формируются в периоды избыточного питания и обеспечивают необходимые потребности организма в периоды голодания). Опорная, защитная, пластическая - жировая ткань полностью или частично окружает различные органы (почки, глазное яблоко, лимфатические узлы, сосудисто-нервные пучки, суставы и др. ) и заполняет пространства между ними; смягчая удары, она защишает их от возможных механических травм, служит опорным и фиксирующим элементом (резкое похудание, например, может привести к смещению почек). Она замещает ткань некоторых органов после их инволюции (тимуса, молочной железы, костного мозга). Теплоизолирующая - жировая ткань обладает свойствами теплоизолятора, благодаря чему она препятствует чрезмерной потере тепла организмом (что особенно важно для человека, в отличие от животных, лишенного шерсти). С этим ее свойством, вероятно связано то, что у северных народов, например, подкожная жировая клетчатка обычно лучше, чем у живущих в средней полосе.

Теплопродуцирующая - часть энергии, образованной вследствие окисления энергоемких молекул жиров, превращается в тепло. Один из видов жировой ткани (бурая жировая ткань) специализирован на выработке значительного количества тепла в результате преобразования в него почти всей полученной при окислении жиров энергии, отчего такую ткань называют "химической печкой". Регуляторная (в процессах миелоидного кроветворения) - жировые клетки входят в состав стромального компонента красного костного мозга, формируя микроокружение развивающихся форменных элементов крови, обеспечивая их питательными веществами и воздействуя на них факторами роста. Изменяя свой объем, жировые клеики влияют на давление внутри мелких костных полостей, содержащих красный костный мозг, и тем самым участвуют в регуляции скорости миграции созревших элементов в сосуды. Депонирующая - жировая ткань накапливает жирорастворимые витамины (А, D, Е, К) и служит крупным депо стероидных гормонов (особенно эстрогенов - женских половых гормонов). Эндокринная - синтезирует эстрогены и гормон, регулирующий потребление пищи лептин. [Лептин - пептидный "фактор насыщения" (открыт в 1994 году) вырабатываемый жировой тканью и способствующий поддержанию постоянства ее массы в организме. Он оказывает действие на уровне гипоталамуса, вызывает ощущение сытости и снижает потребление пищи]. У млекопитающих, включая человека, имеется два вида жировой ткани - белая и бурая, которые различаются: • по цвету (что отражается в их названиях), • рапределению в организме, • метаболической активности, • строению образующих их клеток (адипоцитов), • степени кровеснабжения. БЕЛАЯ жировая ткань. Является преобладающим видом жировой ткани у человека. Она нередко имеет желтоватый оттенок из-за высокого содержания каротиноидов, растворенных в жировой капле адипоцитов.

Эта ткань распределена в организме неравномерно и образует скопления, которые подразделяются на поверхностные и глубокие. ПОВЕРХНОСТНЫЕ скопления располагаются преимущественно подкожно и образуют гиподерму (слой подкожной жировой клетчатки). ГЛУБОКИЕ (висцеральные) скопления белой жировой ткани сосредоточены в области сальника, брыжейки кишки, в забрюшинном пространстве. Различия в зависимости от пола по распрделению жировой ткани в организме (у мужчин преимущественно в верхней половине тела, у женщин - в нижней) обусловливают характерные половые особенности контуров фигуры. Они возникают под влиянием половых гормонов при половом созревании, до которого топография жировой ткани у мальчиков и девочек сходна. СТРОЕНИЕ белой жировой ткани. Имеет дольчатое строение, т. е. состоит из долек (компактных скоплений адипоцитов), разделенных тонкими прослойками рыхлой волокнистой соединительной ткани, несущими кровеносные сосуды и нервы. Кровеносные капилляры и отдельные нервные волокна проникают внутрь долек, располагаясь в узких щелевидных пространствах между адипоцитами. Хотя адипоциты занимают основную часть объема жировой ткани, они составляют, по разным оценкам, лишь 20 - 60 % числа ее клеток. Остальная часть приходится на клетки предшественники адипоцитов, макрофаги, клетки сосудов и лейкоциты крови. Общее число адипоцитов в жировой ткани человека составляет 20 -30 х 109 клеток; при ожирении оно может достигать 100 х 109 клеток. Химически белая жировая ткань представлена: • липидами - 60 - 85 %; • водой - 5 - 30 %; • белками - 2 - 3 %. Адипоциты - крупные (диаметром от 25 - 50 до 150 - 250 мкм) клетки сферической формы, которые в жировых дольках плотно прилегают друг к другу. Ядро адипоцита уплощено и смещено к краю клетки вместе с тонким ободком окружающей его цитоплазмы. Оно содержит умеренно конденсированный хроматин.

БУРАЯ ЖИРОВАЯ ТКАНЬ Содержится у человека в небольшом количестве и, в отличие от белой жировой ткани, сосредоточена лишь в нескольких четко очерченных участках тела (между лопаток, в подмышечных впадинах, на задней поверхности шеи и между ее сосудами, в воротах почек). Она сравнительно хорошо представлена у плода человека и новорожденных (составляя у них 2 - 5 % массы тела). У взрослых бурая жировая ткань почти не обнаруживается, кроме вышеупомянутых участков, однако, полностью она, по-видимому, не исчезает. Ее содержание может даже увеличиваться у пожилых людей и при некоторых заболеваниях. Более того, она служит источником развития некоторых видов доброкачественных опухолей жировой ткани (липом). Важной особенностью бурой жировой ткани является то, что ее содержание мало меняется при недостаточном и избыточном питании. Строение. Бурая жировая ткань, как и белая, образована дольками, состоящими из адипоцитов бурой жировой ткани, среди которых могут находиться отдельные клетки белой жировой ткани. Соединительнотканные прослойки очень тонкие, а кровоснабжение долек чрезвычайно обильное. Адипоциты бурой жировой ткани имеют более мелкие размеры, чем адипоциты белой жировой ткани. Их округлое ядро располагается в центре или эксцентрично (в последнем случае оно не смещено к периферии адипоцита). Цитоплазма содержит множественные жировые капли различных размеров - до 25 мкм. В цитоплазме располагается мелкий АГ, сравнительно слабо развита ЭПС, отдельные рибосомы и включения гликогена. Значительную часть объема цитоплазмы занимают многочисленные митохондрии с высоким содержанием параллельно расположенных крист. При стимуляции активность окислительных процессов в бурой жировой ткани возрастает в сотни раз, что сочетается с многократным усилением кровотока в ее сосудах. Именно в связи с функцией термогенеза бурая жировая ткань сравнительно хорошо развита у новорожденных детей, обладающих несовершенной функцией терморегуляции, и у животных, в особенности, впадающих в спячку. Также бурая жировая ткань играет роль депо жиров высококалорийного резервного материала.
")
РЕТИКУЛЯРНАЯ ТКАНЬ Представляет собой специализированную соединительную ткань, которая входит в качестве структурной основы (стромы) в состав кроветворных тканей - миелоидной и лимфоидной. В этих тканях ее элементы (ретикулярные клетки и ретикулярные волокна) образуют трехмерную сеть, в петлях которой развиваются клетки крови. Ретикулярные клетки имеют отростки цитоплазмы различной длины и толщины, которые соединяются с такими же отростками соседних клеток. При различных воздействиях (воспалительные реакции и т. п. ) клетки ретикулярной ткани могут отделяться, становиться округлыми, свободными, приобретать амебоидную подвижность и превращаться в активных фагоцитов (макрофаги). Функции ткани: • создает необходимые условия для развивающихся клетиок крови; • опорная; • трофическая; • секреторная (ретикулоциты выделяют цитокины и факторы роста); • фагоцитарная; • метаболическая (макрофаги способны захватывать свободное железо → объединяют железо с белками → образуется комплекс, который передается эритробластам, которые дорабатывают его до Нв. СЛИЗИСТАЯ ТКАНЬ Представляет собой видоизмененную рыхлую волокнистую соединительную ткань с резким количественным преобладанием межклеточного вещества, в котором волокнистый компонент развит очень слабо. Вследствие этого она имеет желеобразную консистенцию. В отличие от типичной рыхлой волокнистой ткани, слизистая ткань не содержит мелких кровеносных и лимфатических сосудов и нервных волокон. Эта ткань имеется у плода, у которых она составляет основу пупочного канатика и известна под названием вартонова студня (соединяет плод с плацентой матери). Он имеет плотную оболочку, 1 артерию, 2 вены. У взрослых близкое строение имеет ткань, образующая так называемое стекловидное тело глазного яблока. Клетки слизистой ткани в большинстве сходны с фибробластами, однако нередко содержат в цитоплазме значительное количество гликогена. Они имеют отросчатую форму, часто контактируют друг с другом, синтезируют преимущественно основное аморфное вещество, и лишь в очень незначительных количествах - коллагеновые волокна. Помимо них в небольшом числе присутствуют также макрофаги и лимфоциты.

ПИГМЕНТНАЯ ТКАНЬ Напоминает рыхлую волокнистую соединительную ткань, однако содержит значительно большее количество пигментных клеток, которые являются ее численно преобладающими и функционально ведущими клеточными элементами. В ней имеется большое количество кровеносных сосудов. Наиболее характерными участками расположения этой ткани служат радужка глаза и сосудистая оболочка глаза. Клетки пигментной соединительной ткани представлены многочисленными фибробластами, фиброцитами, гистиоцитами, тучными клетками, лейкрцитами и пигментными клетками. Пигментные клетки подразделяются Межклеточное вещество включает многочисленные коллагеновые, ретикулярные и эластические волокна, формирующие трехмерные сети, а также основное аморфное вещество. меланоциты Отросчатые клетки, контактирующие с другими клетками этой ткани и волокнами межклеточного вещества. Ядро - удлиненное, с многочисленными вдавливаниями кариолеммы. Цитоплазма содержит развитый синтетический аппарат и большое число гранул (меланосом), постепенно заполняющихся темным пигментом – меланином По мнению некоторых авторов, истинные меланоциты располагаются преимущественно в эпителии. меланофоры Удлиненные или отросчатые клетки со слабо развитым синтетическим аппаратом и значительным числом зрелых меланиновых гранул в цитоплазме. Эти клетки не способны к синтезу меланина, меланиновые гранулы поглощаются ими после того, как они выделяются синтезировавшими их меланоцитами.
Консультация бакалавров.ppt