
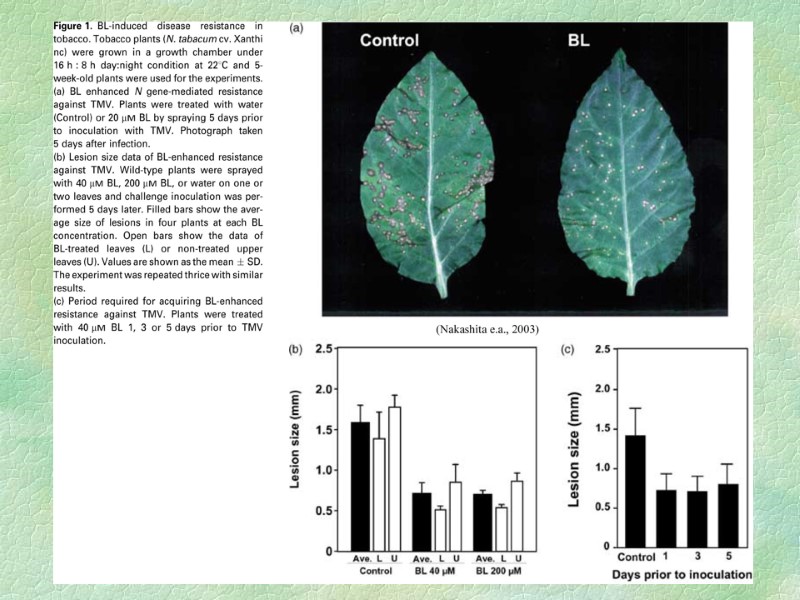
Брассиностероиды В растительных организмах выявлено более 60 веществ,


Брассиностероиды
, которые синтезируются")
В растительных организмах выявлено более 60 веществ, называемых брассиностероидами (БС), которые синтезируются главным образом из кампестерола. Больше всего БС содержат генеративные органы растений. Брассинолид и кастастерон

Одной из особенностей БС, отличающей их от других фитогормонов, является способность стимулировать ростовые процессы в очень низких концентрациях (1061012 М). Стимулирующее действие БС на ростовые процессы связано с активацией и деления, и растяжения клеток. Наиболее специфичный для БС эффект заключается в активации роста второго междоузлия фасоли или гороха. Еще более чувствительным биотестом на БС является рост эпикотилей маша. Стимулирующий эффект БС проявляется в концентрации 1010 М и ниже. БС участвуют в регуляции ростовых процессов во взаимодействии с другими фитогормонами. Выявлено, что БС и гиббереллины оказывают аддитивное действие на рост гипокотилей огурца. Обнаружен сильный синергический эффект ИУК и брассинолида. БС в низких концентрациях повышает устойчивость растений к таким стрессовым воздействиям, как резкие колебания температуры, засуха, засоление, аноксия и воздействие патогенов.
, брассинолид-дефицитный мутант det2 (в центре), брассинолид-дефицитный мутант dim")
16-дневные проростки арабидопсиса дикий тип (слева), брассинолид-дефицитный мутант det2 (в центре), брассинолид-дефицитный мутант dim (справа).
 используют как биотест на БС. Действие брассинолида (справа)")
Этиолированные проростки риса (слева) используют как биотест на БС. Действие брассинолида (справа) на наклон листовой пластинки риса (справа).
, которая является негативным регулятором")
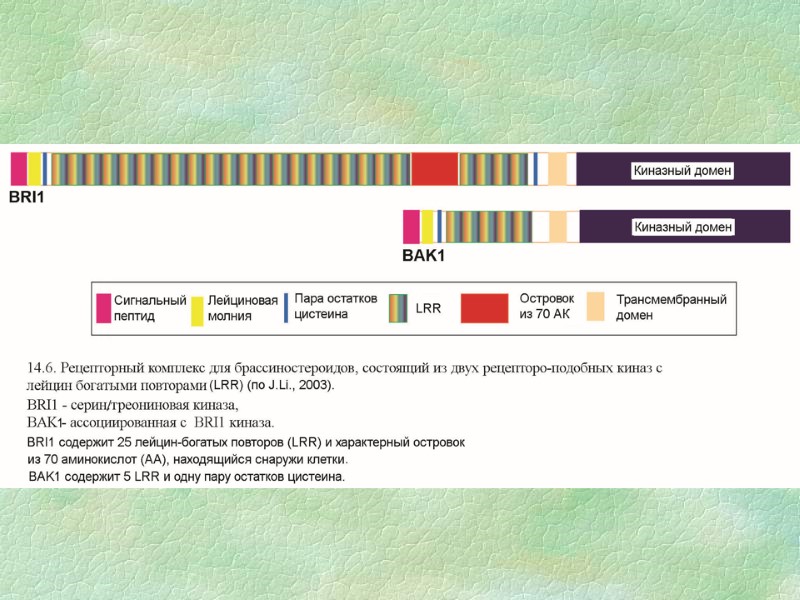
Ключевым элементом в ходе трансдукции БС-сигнала является протеинкиназа BIN2 (BRASSINOSTEROID-INSENSITIVE), которая является негативным регулятором процессов, следующих после рецептора BRI1. Протеинкиназа BIN2 взаимодействует с белками – BZR1 (BRASSINASOLE RESISTANT1) и BES1 (BRI1- Еthyl methane sulphonate-SUPPESSOR1), которые функционируют как позитивные регуляторы БС-сигнала и обладают способностью контролировать экспрессию БС-зависимых генов. (А) В отсутствии брассиностероидов белки BZR1 и BES1 фосфорилируются киназой BIN2, что делает их мишенями для деградации в 26S протеасомах. (Б) При связывании брассинолида с рецепторным комплексом происходит образование димера BRI1-BAC1 и последующая активация этих рецепторных киназ посредством их трансфосфорилирования. Это приводит к подавлению негативного регулятора BIN2. Дальнейшая передача БС-сигнала осуществляется белками BES1 и BZR1, которые перемещаются в ядро и функционируют, как активатор и репрессор (соответственно) БС-зависимых генов. (В) После рецепции и передачи БС-сигнала происходит удаление рецепторного комплекса из плазматической мембраны эндоцитозом, сопровождающееся его протеолизом Выявлены десятки генов, чувствительные к БС, которые контролируют биосинтез элементов клеточной стенки и цитоскелета, синтез гормонов и их сигналинг (в особенности ауксина), мишенью для БС может являться вакуолярная H+-АТРаза

Модель рецепции и передачи жасмонатного сигнала с участием белков COI1–JAZ у арабидопсиса (по Acosta, Farmer, 2010, с изменениями). JA-Ile — жасмонил-L-изолейцин ; SCFCOI1 — комплекс, который функционирует, как Е3-тип убиквитин лигаз; COI — рецептор жасмоната, белок F-бокса убиквитин лигазного комплекса; 26S — протеасома; Z (ZIM) и J (Jas) — домены JAZ-белка, отвечающие за белок-белковые взаимодействия; ТФ — транскрипционные факторы жасмонат-зависимых генов

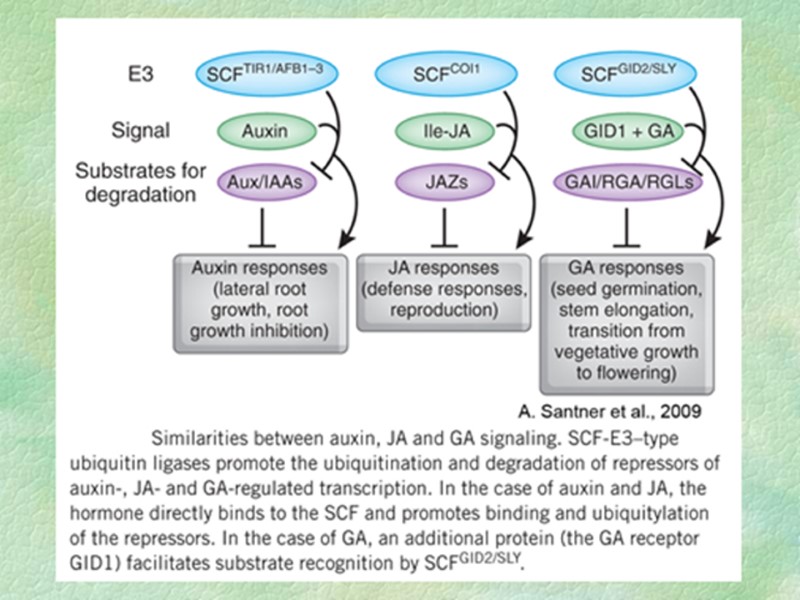
К жасмонатам, для которых получены наиболее веские доказательства их биологической активности, относятся (Acosta, Farmer, 2010): (+)-7-изо-жасмонил-L-изолейцин — играет важную роль в жасмонатной сигнализации в листьях (возможно, и цветках) арабидопсиса; жасмонил-L-триптофан — ингибитор передачи ауксинового сигнала в корнях арабидопсиса; ОФДК и динор-ОФДК - обладают сигнальными функциями, независящими от канонического жасмонатного сигналинга, или используют его частично. Для расшифровки жасмонатного сигнала очень полезным оказался мутант coi1, устойчивый к бактериальному токсину коронатину и метилжасмонату (коронатин является структурным аналогом и имитирует действие ЖК). Оказалось, что белок COI1 (coronatine insensitive1) необходим практически для всех процессов, контролируемых ЖсК. В мутантах coi1 жасмонат-индуцируемые гены не экспрессируются. В дальнейшем было показано, что белок COI1 представляет собой субъединицу F-бокса убиквитин-лигазного комплекса типа SKP1-CUL1-F-box (SCF) и является рецептором жасмоната (см. Приложение 2. Рецептор жасмоновой кислоты). Механизм рецепции и трансдукции жасмонатного сигнала приведен на рис. 8.37.

Жасмоновая кислота контролирует устойчивость растений к насекомым и патогенам, созревание плодов и рост корня, изгиб усиков и производство жизнеспособной пыльцы. Содержание ЖК в тканях растений возрастает при таких механических раздражениях, как изменение тургорного давления при водном дефиците, движения усиков, взаимодействие корневых волосков с частицами почвы. Жасмонаты растений


Наиболее важной функцией ЖК является ее участие в ответных реакциях при повреждении растений насекомыми и патогенами. Поврежденные ткани отличаются очень высокой концентрацией этого фитогормона, что резко повышает устойчивость растений к вредителям. Повышение содержания жасмоната активирует экспрессию целого каскада генов, которые кодируют: белки, формирующие механические барьеры в клеточной стенке на пути инфекции; 2) ферменты, участвующие в синтезе фитоалексинов; 3) ингибиторы протеаз, обеспечивающие защиту растений от повреждения насекомыми; 4) белки, обладающие фунгицидной активностью, тионин, осмотин и др.

Развитие пыльцы Растрескивание пыльника Развитие тычинок Поранение, Вредители, Патогены, Элиситоры Защитные реакции Привлечение насекомых Биосинтез жасмонатов у арабидопсиса хлоропласт цитоплазма пероксисома OPDA: 12-oxo-phytodienoic acid

Салициловая кислота
 катализируется 2-гидроксилазой бензойной кислоты (цитохром-Р450-монооксигеназой), который")
Превращение бензойной кислоты в салициловую (СК) катализируется 2-гидроксилазой бензойной кислоты (цитохром-Р450-монооксигеназой), который использует молекулярный кислород для гидроксилирования бензойной кислоты. СК может образовывать метиловый эфир и (или) связываться с глюкозой под действием глюкозилтрансферазы. Метилсалицилат является летучим соединением и легко превращается в тканях-мишенях в СК. Синтез салициловой кислоты из фенилаланина

Когда растение подвергается инфекции в одном месте, часто происходит формирование устойчивости к последующему нападению во всех тканях и органах организма. Это явление названо системной приобретенной устойчивостью (system aquiered resistance, SAR), или системным приобретенным иммунитетом (ПИ). Иммунитет развивается в течение нескольких дней и проявляется в повышенной устойчивости всего растения к повторному заражению широким набором видов патогенов (грибы, бактерии, вирусы) и насекомых-фитофагов. Т.е. процесс иммунизации растительных тканей неспецифичен в отношении патогена. Системный приобретенный иммунитет растений

Вещества, индуцирующие системный иммунитет растений В регулировании процессов системной устойчивости наиболее важное значение имеют салициловая (СК) и жасмоновая кислоты. В формировании приобретенного иммунитета растений ключевая роль принадлежит салициловой кислоте. Заражение растений патогеном индуцирует накопление СК как в инфицированных, так и в неинфицированных тканях. Обработка СК и ее аналогами (INA, BTH) активирует PR-гены и устойчивость растений без инокуляции патогенами. Не накапливающие салицилат мутантные растения не способны к ПИ.
 SRK – S-locus receptor kinase")
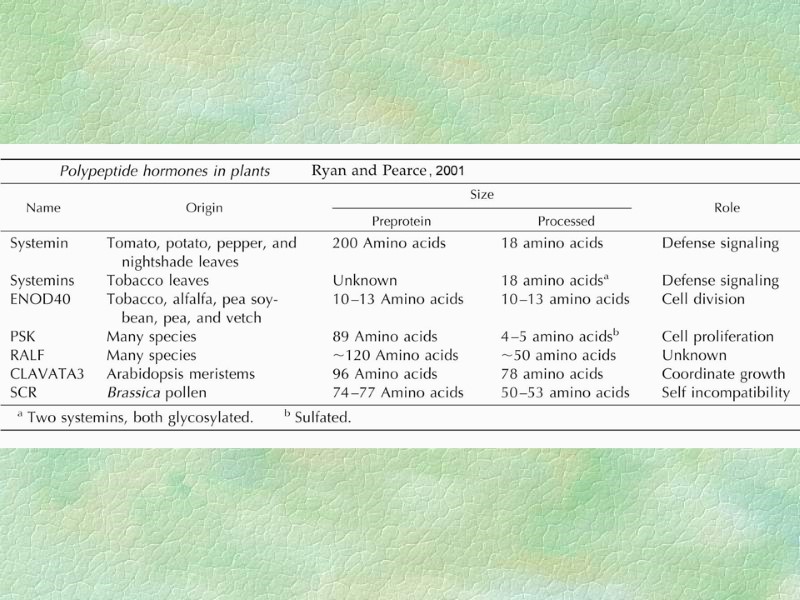
SCR – S-locus cysteine-rich protein (SP11 – S-locus protein) SRK – S-locus receptor kinase Пептидные гормоны растений
 SRK – S-locus receptor kinase")
SCR – S-locus cysteine-rich protein (SP11 – S-locus protein) SRK – S-locus receptor kinase Пептидные гормоны растений

Системин

Ответные реакции растения на повреждение фитофагами У томатов системин включается в системную регуляцию более 20 генов, контролирующих устойчивость к повреждению фитофагами, патогенами, поранению, а также к ряду абиотических стрессовых воздействий. Регуляция экспрессии некоторых “защитных” генов системином может осуществляться вместе с другими гормонами - АБК, этиленом и жасмоновой кислотой.

Растения томатов, поврежденные homworm larvae: Слева растения дикого типа. Справа мутанты spr2 (suppressor of prosystemin-mediated responses2) с нарушением синтеза системина.

CLAVATA3 Рецептором CLV3 являются белки CLV1 и CLV2 (рецепторно-киназный комплекс молекулярной массой 185 кDa). После связывания CLV3 с рецептором происходит автофосфорилирование CLV1 и дальнейшая передача сигнала.

SCR – S-locus cysteine-rich protein SRK – S-locus receptor kinase

ENOD40

Летучие выделения растений
 – аттрактанты для насекомых")
Летучие выделения растений (каиромоны) – аттрактанты для насекомых
 при повреждении растений насекомыми.")
Летучие выделения растений, формируемые (локально или системно) при повреждении растений насекомыми.

Электрофизио-логический ответ насекомых-фитофагов на летучие выделения растений – каиромоны.
5130-bs-zhk-sk.ppt
- Количество слайдов: 32