тема5.ppt
- Количество слайдов: 64



Биохимические свойства микроорганизмов Тема 5

Характеристика выделенной культуры. Свойства: Ø Культуральные - характер роста микроорганизма на питательных средах. Ø Биохимические - способность ферментировать различные субстраты (углеводы, белки, аминокислоты и др. ), образовывать в процессе жизнедеятельности различные биохимические продукты за счет активности различных ферментных систем и особенностей обмена веществ.

Питание бактерий № 1

Типы питания бактерий

Питание бактерий Автотрофы Гетеротрофы Источник углероданеорганические соединения Сапрофиты Паразиты Большая часть бактерий источник 0, 1% видов

В отличие от животных и растительных организмов, микроорганизмы характеризуются разнообразием типов питания, которые выделяют по трем основным критериям –– источник углерода, источник энергии и донор электронов (водорода). В зависимости от природы источника углерода все микроорганизмы разделены на 2 большие группы –– автотрофы, использующие углекислоту, и гетеротрофы, требующие для роста и размножения готовых органических веществ. С учетом разнообразия источников энергии и доноров электронов эти группы подразделены на подгруппы, в результате чего у микроорганизмов выделены 8 типов питания. Каждый тип питания характерен для определенных микроорганизмов и отражает их физиологобиохимические свойства. Большинство микроорганизмов, в том числе и патогенных, имеют тип питания, при котором источником углерода, энергии и донорами электронов являются органические вещества. 6
. Ряд бактерии (в т.")
Для большинства бактерий источником энергии служит окисление химических веществ (хемотрофы). Ряд бактерии (в т. ч. пурпурные и зелёные бактерии, цианобактерии) в ходе фотосинтеза преобразуют энергию света в энергию химических связей органических соединений (фототрофы). Если окислению подвергаются неорганические вещества (т. е. они используются в качестве доноров водорода), бактерии называют литотрофами (хемосинтетиками), если же вещества органической природы, - органотрофами. Исходя из этого выделяют 8 типов питания и соответствующих им групп бактерии: фотолитоавтотрофы (цианобактерии, анаэробные пурпурные бактерии), фотолитогетеротрофы (некоторые анаэробные бактерии), фотоорганогетеротрофы (несерные пурпурные бактерии), фотоорганоавтотрофы (редкий тип питания, свойственный некоторым пурпурным бактериям), хемолитоавтотрофы (например, нитрификаторы, тионовые бактерии), хемолитогетеротрофы (многие сульфатвосстанавливающие бактерии), хемоорганоавтотрофы (мн. водородные бактерии), хемоорганогетеротрофы (основной массив бактерийорганотрофов). Известны облигатные паразиты (паратрофы), использующие только сложные органические вещества, образуемые организмом-хозяином.

Потребности микроорганизмов в органических источниках углерода весьма разнообразны. Некоторые виды являются ≪всеядными≫ и могут потреблять различные по химической природе вещества, другие отличаются большей избирательностью и используют лишь некоторые из них. Специфичность набора органических соединений, свойственного каждому виду микроорганизмов, учитывается при создании элективных и дифференциально-диагностических сред, широко применяемых в клинической микробиологии для быстрой идентификации определенных групп микроорганизмов. Обычно микроорганизмы усваивают определенные оптические изомеры –– сахара, относящиеся к D-ряду, аминокислоты –– к L-ряду. Очень мало микроорганизмов обладают ферментами, под действием которых один оптический изомер превращается в другой. Использовать такие биополимеры как крахмал, полисахариды, белки, жиры могут только те микроорганизмы, у которых синтезируются определенные гидролитические ферменты –– амилазы, протеазы, липазы в форме экзоферментов, т. е. ферментов, выделяемых клеткой в среду обитания. 8

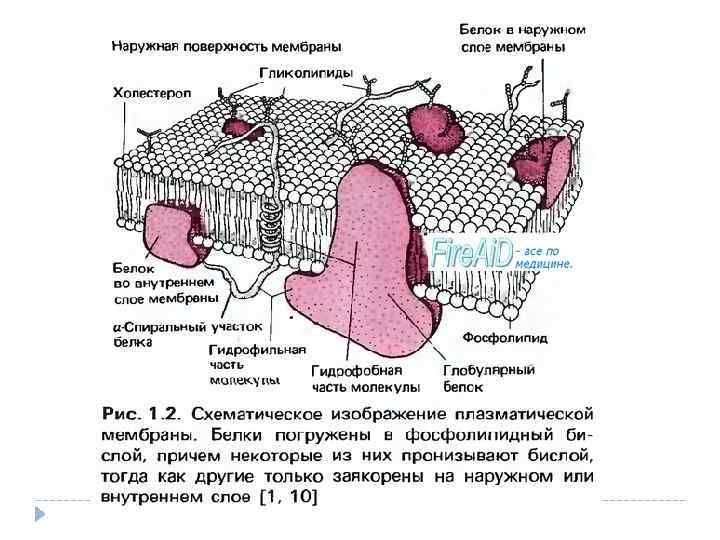
Поступление веществ в клетку и выделение продуктов обмена в окружающую среду происходит у микроорганизмов через всю поверхность тела. У микроорганизмов очень большая по сравнению с объемом всасывающая пищу поверхность клетки, что обусловливает весьма активный обмен веществ. Возможность проникновения веществ извне в клетку обусловлена многими факторами: величиной и структурой их молекул; способностью растворяться в компонентах цитоплазматической мембраны или вступать с ними в химические соединения; концентрацией веществ в клетке и в среде; электрическим зарядом клетки и другое. Вещества питательной среды могут поступать в клетку только в растворенном состоянии. Нерастворимые сложные органические соединения должны подвергнуться расщеплению на более простые вне клетки, что происходит с помощью экзоферментов микроорганизмов. Клеточная стенка проницаема и задерживает лишь макромолекулы. Цитоплазматическая мембрана обладает полупроницаемостью. Она служит осмотическим барьером, проницаемость её для различных веществ неодинакова.
 простая")
Пути поступления метаболитов и ионов в микробную клетку Пассивный транспорт (без энергетических затрат) простая диффузия Активный транспорт (с затратой энергии, против градиента концентрации; при этом происходит взаимодействие субстрата с белком-переносчиком на поверхности цитоплазматической мембраны) облегченная диффузия (по градиенту концентрации, с помощью белковпереносчиков).
. Активная")
Наиболее известны два пути проникновения веществ в клетку: осмос и адсорбция (специфический перенос). Активная роль в этих процессах принадлежит цитоплазматической мембране. О с м о с представляет собой диффузию веществ в растворах через полупроницаемую мембрану. Как известно, через такие мембраны могут диффундировать вещества, находящиеся в состоянии истинных растворов. Возникает осмос под действием разности осмотических давлений в растворах по обе стороны полупроницаемой мембраны. Величина осмотического давления раствора зависит от молярной концентрации растворённых в нём веществ. В относительно слабых растворах осмотическое давление изменяется пропорционально концентрации растворённых веществ. Чем больше разность осмотических давлений (концентраций растворов) по обе стороны полупроницаемой мембраны, тем с большей интенсивностью осмотирует растворитель (вода) в раствор с большим осмотическим давлением. Осмотируют растворённые в воде вещества; при этом каждое диффундирует в соответствии с его собственным (парциальным) осмотическим давлением, то есть в раствор с его меньшей концентрацией.

Таким образом, при осмотическом проникновении питательных веществ в клетку движущей силой служит разность осмотических давлений между средой и клеткой. Такой пассивный перенос веществ не требует затраты энергии и протекает до выравнивания концентрации с наружным раствором. Поскольку поступившие в клетку вещества включаются в реакции конструктивного и энергетического обмена, концентрация некоторых из них будет ниже, чем в среде, и поступление данных веществ возможно до полного исчерпания их из субстрата. Если осмотическое давление микробных клеток, обусловленное растворёнными в клеточном соке веществами, несколько выше, чем в среде, то за счёт притока из неё воды в клетке создаётся определённое упругое напряжение, называемое тургором. Протопласт клетки при этом прижимается к клеточной оболочке, слегка растягивая её. Если микроорганизм попадает в субстрат, осмотическое давление которого выше, чем в клетке, то цитоплазма отдает воду во внешнюю среду. Питательные вещества в клетку не поступают, содержимое клетки уменьшается в объёме, и протопласт отстаёт от клеточной оболочки. Это явление называется плазмолизом клетки. При чрезмерно низком осмотическом давлении внешней среды может наступить явление, обратное плазмолису, когда вследствие высокой разности осмотических давлений цитоплазма быстро переполняется водой. Это может привести к разрыву клеточной оболочки, что наблюдается, например, при помещении бактерий в дистиллированную воду.

Второй путь поступления веществ в клетку – активный – путём переноса их особыми, локализованными в цитоплазматической мембране веществами ферментной природы. Эти переносчики, называемые пермеазами, обладают субстратной специфичностью. Каждый транспортирует только определённое вещество, имеющее сходную с белком-переносчиком стереохимическую структуру молекулы. На внешней стороне цитоплазматической мембраны переносчик адсорбирует вещество – вступает с ним во временную связь и диффундирует комплексно через мембрану, отдавая на внутренней стороне её транспортируемое вещество в цитоплазму. Вещество может поступать и тогда, когда концентрация его в клетке больше, чем в среде. При таком переносе веществ затрачивается энергия. При этом транспортируемое вещество может подвергнуться изменению, например из не растворимого в мембране переходит в растворимое состояние. Цитоплазматическая мембрана, таким образом, является не только осмотическим барьером, но и обладает избирательной проницаемостью.
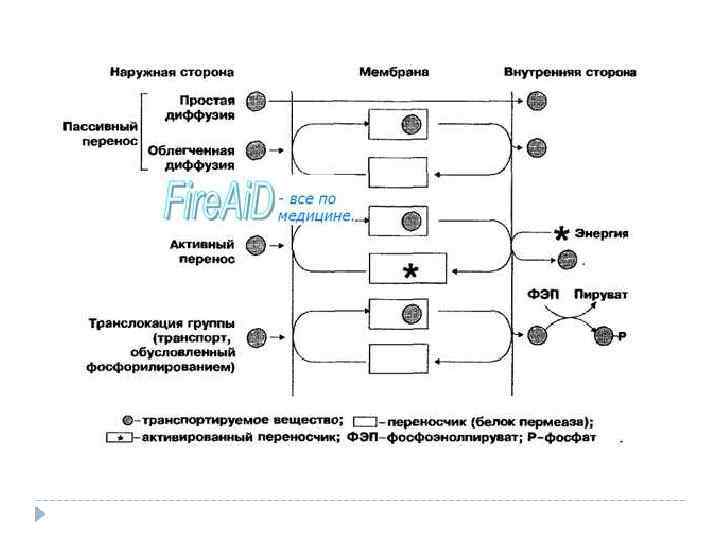

Концентрация некоторых веществ в бактериальной клетке может в сотни раз превышать их содержание в окружающей среде. Транспорт таких веществ в клетку происходит против градиента концентрации перемещаемого вещества, требует затрат энергии и реализуется при помощи специфических переносчиков. Например, в случае активного транспорта, как и при облегчённой диффузии, перемещение конкретного вещества через ЦПМ осуществляет специфическая для такого вещества пермеаза. Активный транспорт сопровождается увеличением свободной энергии. Это обстоятельство делает процесс активного транспорта энергозависимым. У бактерий подобный тип поступления веществ доминирует; с его помощью транспортируются многие сахара, белки и другие вещества. Нередко у грамотрицательных бактерий в активном переносе участвуют специальные белки, отличные от пермеаз, но действующие в ассоциации с ними. Эти белки локализованы в периплазматическом пространстве. После проникновения вещества в клетку комплекс «субстрат — белок периплазмы — пермеаза» диссоциирует и для попавшего в клетку вещества вероятность выхода наружу резко снижается.

Примером активного транспорта служит концентрирование лактозы в клетках кишечной палочки. Эта система известна как β-галактозидпермеазная система. Энергия используется для снижения сродства пермеазы к лактозе на внутренней стороне ЦПМ по сравнению с ее сродством к тому же субстрату на внешней. В результате скорость выхода вещества наружу становится меньше, чем скорость его поступления внутрь клетки, и концентрация лактозы в клетке возрастает. Если блокировать образование энергии, то βгалактозидпермеазная система теряет способность осуществлять активный транспорт и переключается на облегчённую диффузию, обнаруживая одинаковое сродство к β-галактозидам по обе стороны мембраны.

Транспорт, обусловленный фосфорилированием — энергозависимый процесс, используемый при утилизации углеводов. Основной механизм транспорта связан с фосфорилированием субстрата, что делает невозможным его выход из клетки. Первоначально происходит фосфорилирование мембранного фермента (так называемый фермент 2, или Ф 2) в цитоплазме за счёт фосфоенолпирувата. Фосфорилированный фермент связывает углевод (например, глюкозу или маннозу) на поверхности ЦПМ и транспортирует его в цитоплазму. Затем комплекс диссоциирует с высвобождением углевода, связанного с фосфатной группой. За счёт фосфорилирования углевод аккумулируется в клетке и не способен выходить из неё. Данный тип транспорта не рассматривают как активный, поскольку концентрация неизменённого питательного вещества внутри клетки может быть одинаковой с его внеклеточным содержанием. Но в целом этот процесс напоминает активный транспорт против градиента концентрации вещества, так концентрация химически изменённого питательного соединения внутри клетки может значительно превышать концентрацию неизменённого соединения в среде.

Экзо- и эндоферменты № 2

Все питательные вещества и любые элементы, подвергающиеся взаимодействиям и превращениям с участием бактерий, вступают в реакции при участии ферментов. Ферменты [от лат. fermentum, закваска], или энзимы [от греч. enzyme, дрожжи или закваска], — специфичные и эффективные белковые катализаторы, присутствующие во всех живых клетках. За каждое превращение одного соединения в другое ответственен особый фермент. • Ферменты снижают энергию активации, обеспечивая протекание таких химических реакций, которые без них могли бы проходить только при высокой температуре, избыточном давлении и при других нефизиологических условиях, неприемлемых для живой клетки. • Ферменты увеличивают скорость реакции примерно на 10 порядков, что сокращает полупериод какой-либо реакции с 300 лет до одной секунды. • Ферменты «узнают» субстрат по пространственному расположению его молекулы и распределению зарядов в ней. За связывание с субстратом отвечает определённый участок молекулы ферментативного белка — его каталитический центр. При этом образуется промежуточный ферментсубстратный комплекс, который затем распадается с образованием продукта реакции и свободного фермента.
 Простые Сложные (трипсин, пепсин) (Цитохромоксидаза, пероксидаза) (по действию) Экзоферменты")
ФЕРМЕНТЫ БАКТЕРИЙ (по составу) Простые Сложные (трипсин, пепсин) (Цитохромоксидаза, пероксидаза) (по действию) Экзоферменты (выделяются во внешнюю среду – факторы агрессии) Эндоферменты (необходимы для внутренних процессов жизнедеятельности клетки)

В зависимости от акцептора протонов и электронов среди бактерий различают аэробы, факультативные анаэробы и облигатные анаэробы. Для аэробов акцептором является кислород. Факультативные анаэробы в кислородных условиях используют процесс дыхания, в бескислородных – брожение. Для облигатных анаэробов характерно только брожение, в кислородных условиях наступает гибель микроорганизма из-за образования перекисей, идет отравление клетки. В микробной клетке ферменты являются биологическими катализаторами. По строению выделяют: 1) простые ферменты (белки); 2) сложные; состоят из белковой (активного центра) и небелковой частей; необходимы для активизации ферментов. Различают также: 1) конституитивные ферменты (синтезируются постоянно независимо от наличия субстрата); 2) индуцибельные ферменты (синтезируются только в присутствии субстрата). Набор ферментов в клетке строго индивидуален для вида. Способность микроорганизма утилизировать субстраты за счет своего набора ферментов определяет его биохимические свойства.
 экзоферменты (действуют вне клетки; принимают участие в процессе распада")
По месту действия выделяют: 1) экзоферменты (действуют вне клетки; принимают участие в процессе распада крупных молекул, которые не могут проникнуть внутрь бактериальной клетки; характерны для грамположительных бактерий); 2) эндоферменты (действуют в самой клетке, обеспечивают синтез и распад различных веществ). В зависимости от катализируемых химических реакций все ферменты делят на шесть классов: 1) оксидоредуктазы (катализируют окислительно-восстановительные реакции между двумя субстратами); 2) трансферазы (осуществляют межмолекулярный перенос химических групп); 3) гидролазы (осуществляют гидролитическое расщепление внутримолекулярных связей); 4) лиазы (присоединяют химические группы по двум связям, а также осуществляют обратные реакции); 5) изомеразы (осуществляют процессы изомеризации, обеспечивают внутреннюю конверсию с образованием различных изомеров); 6) лигазы, или синтетазы (соединяют две молекулы, вследствие чего происходит расщепление пирофосфатных связей в молекуле АТФ).

• Получение микробных ферментов — важнейшая отрасль промышленной микробиологии. Например, для улучшения пищеварения применяют готовые препараты ферментов — амилазы, целлюлазы, протеазы, липазы, облегчающих соответственно гидролиз крахмала, целлюлозы, белка и липидов. При изготовлении сладостей для предупреждения кристаллизации сахарозы применяют инвертазу дрожжей, для осветления фруктовых соков — пектиназу. Коллагеназа клостридий и стрептокиназа стрептококков, гидролизующие белки, способствуют заживлению ран и ожогов. Литические ферменты бактерий, секретируемые в окружающую среду, действуют на клеточные стенки патогенных микроорганизмов и служат эффективным средством в борьбе с последними, даже если они обладают множественной устойчивостью к антибиотикам. В качестве инструментария в биоорганической химии, генной инженерии и генотерапии используют выделенные из бактерий рибонуклеазы, дезоксирибонуклеазы, полимеразы, ДНК-лигазы и прочие ферменты, направленно модифицирующие нуклеиновые кислоты.

Тест на наличие каталазы Каталаза гидролизует превращение: Н 2 О 2 → вода + O 2 газ Staphylococcus aureus Enterococcus faecium
, которая катализирует транспорт")
Тест на наличие оксидазы Тест определяет наличие цитохром оксидазы (дыхательный фермент), которая катализирует транспорт электронов к конечному акцептору (O 2) Колония помещается на фильтровальную бумагу, импрегнированную тетраметил-p-фенилендиамином дигидрохлоридом

Тест на наличие коагулазы Staphylococcus epidermidis Staphylococcus aureus

Ферментация глюкозы: 1 – незасеянная пробирка, 2 А – отсутствие ферментации, 2 В – ферментация до кислоты, 3 А и 3 В – ферментация до кислоты и газа)

Гидролиз желатины
")
Определение ДНКазы (справа – положительный результат)
")
Способности дезаминировать фенилаланин (две пробирки справа – положительный результат)
 и")
Наличие уреазы Среда содержит индикатор (изменяет окраску при р. Н более 8, 4) и мочевину 1. Контроль 2. Proteus vulgaris 3. E. coli

Определение протеолитических бактерий на молочно – пептонном агаре

Полоски с реактивом Ковача на индол

Пластический и энергетический обмен № 3
 белковый; 2) углеводный; 3) липидный; 4) нуклеиновый. Белковый")
Основными видами пластического обмена являются: 1) белковый; 2) углеводный; 3) липидный; 4) нуклеиновый. Белковый обмен характеризуется катаболизмом и анаболизмом. В процессе катаболизма бактерии разлагают белки под действием протеаз с образованием пептидов. Под действием пептидаз из пептидов образуются аминокислоты. Распад белков в аэробных условиях называется тлением, в анаэробных – гниением.
 Синтез сложных соединений из более простых, Образование и обновление Катаболизм")
Метаболизм Анаболизм (конструктивный обмен) Синтез сложных соединений из более простых, Образование и обновление Катаболизм (энергетический обмен) Расщепление сложных соединений до низкомолекулярных с высвобождением энергии клеточных структур Восстановительные реакции Окислительные реакции

Конструктивный метаболизм микроорганизмов в целом направлен на синтез четырех основных типов биополимеров –– белков, нуклеиновых кислот, полисахаридов и липидов. Для биосинтеза белков и нуклеиновых кислот важнейшим элементом, кроме углерода, является азот. Самым доступным источником азота для большинства микроорганизмов являются ионы аммония, которые они могут получать из солей органических и неорганических кислот, аминокислот, белков и других азотсодержащих веществ. Для многочисленной группы бактерий, главным образом патогенных, в качестве источников азота необходимы азотсодержащие органические вещества. Если таким источником азота являются аминокислоты, микроорганизмы могут их использовать непосредственно для синтеза белков, либо предварительно осуществить их дезаминирование, а выделившиеся при этом аминогруппы использовать для синтеза собственных аминокислот, белков. 38

Однако, для роста некоторых микроорганизмов необходимы определенные аминокислоты, которые они не могут синтезировать сами. К числу микроорганизмов, нуждающихся в таких ≪незаменимых≫ аминокислотах, относятся золотистый стафилококк, гемолитический стрептококк, молочнокислые бактерии и некоторые другие. В зависимости от физиологических особенностей микробов, количество ≪незаменимых≫ аминокислот различно –– для золотистого стафилококка обязательно наличие в питательной среде лишь триптофана и цистина, а для гемолитического стрептококка –– 17 аминокислот. Белки, как источники азота, доступны только тем микроорганизмам, которые образуют протеолитические ферменты, выделяемые в среду (т. е. в экзоформе). Под действием этих ферментов белки расщепляются до более низкомолекулярных веществ –– пептонов и аминокислот. 39

Энергетический метаболизм микроорганизмов, как и конструктивный, характеризуется разнообразием биохимических механизмов. В этом метаболизме различают три основных типа ––аэробное дыхание, анаэробное дыхание и брожение, наиболее распространенным из которых является аэробное дыхание. В этом процессе органическое вещество окисляется до углекислоты и воды с максимальным выделением энергии, заключенной в этом веществе. Многие микроорганизмы с аэробным дыханием –– строгие аэробы, однако некоторые из них относятся к факультативным аэробам, поскольку они могут образовывать АТФ и в анаэробных условиях ––путем брожения. 40

Простейшим по метаболизму типом энергетического метаболизма является брожение. Процессы брожения протекают в анаэробных условиях и сопровождаются выделением энергии. Основным субстратом брожения являются углеводы, однако бактерии могут сбраживать органические кислоты, в том числе и аминокислоты, а также пурины и пиримидины. Известно много типов брожения, каждый из которых вызывается особой группой микроорганизмов и в соответствии с механизмом сопровождается образованием специфических конечных продуктов. Конечными продуктами брожений обычно являются различные органические кислоты –– молочная, уксусная, янтарная, лимонная и др. , а также спирты (этиловый, бутиловый, пропиловый), углекислый газ и водород. По выходу основного конечного продукта выделяют и соответствующие типы брожения. В силу того, что при брожении не происходит полного окисления субстрата и в среду выделяются частично окисленные вещества, еще содержащие энергию –– органические кислоты, спирты и др. , общий выход АТФ в этом процессе на 1 моль сбраживаемого субстрата значительно (~ в 30 раз) ниже, чем при метаболизме того же субстрата в процессах дыхания. 41
 спиртовое (характерно для грибов);")
В зависимости от конечных продуктов выделяют следующие виды брожения: 1) спиртовое (характерно для грибов); 2) пропионионово-кислое (характерно для клостридий, пропиони-бактерий); 3) молочнокислое (характерно для лактококков); 4) маслянокислое (характерно для сарцин); 5) бутилденгликолевое (характерно для бацилл). Наряду с основным анаэробным распадом (гликолизом) могут быть вспомогательные пути расщепления углеводов (пентозофосфатный, кетодезоксифосфоглюконатный и др. ). Они отличаются ключевыми продуктами и реакциями.

Липидный обмен осуществляется с помощью ферментов – липопротеиназ, летициназ, липаз, фосфолипаз. Липазы катализируют распад нейтральных жирных кислот, т. е. ответственны за отщепление этих кислот от глицерина. При распаде жирных кислот клетка запасает энергию. Конечным продуктом распада является ацетил-Ко. А. Биосинтез липидов осуществляется за счет ацетилпереносящих белков. При этом ацетильный остаток переходит на глицерофосфат с образованием фосфатидных кислот, а они уже вступают в химические реакции с образованием сложных эфиров со спиртами. Эти превращения лежат в основе синтеза фосфолипидов. Бактерии способны синтезировать как насыщенные, так и ненасыщенные жирные кислоты, но синтез последних более характерен для аэробов, так как требует кислорода. Нуклеиновый обмен бактерий связан с генетическим обменом. Синтез нуклеиновых кислот имеет значение для процесса деления клетки. Синтез осуществляется с помощью ферментов: рестриктазы, ДНК-полимеразы, лигазы, ДНК-зависимой-РНК-полимеразы. Рестриктазы вырезают участки ДНК, убирая нежелательные вставки, а лигазы обеспечивают сшивку фрагментов нуклеиновой кислоты. ДНК-полимеразы ответственны за репликацию дочерней ДНК по материнской. ДНК-зкависимые-РНКполимеразы отвечают за транскрипцию, осуществляют построение РНК на матрице ДНК.

Идентификация микроорганизмов и их внутривидовое типирование № 4

Цель идентификации — установить таксономическое положение исследуемого штамма на основании сравнения его свойств с изученными и принятыми (официально зарегистрированными) видами. Поэтому результатом идентификации обычно является отождествление исследуемого микроорганизма с каким-нибудь видом или отнесение к определенному роду.

Идентификация выделенного штамма микроорганизма является наиболее трудоёмким и материально затратным исследованием. Одним из наиболее часто применяемых в мировой практике в настоящее время способов быстрой идентификации различных бактерий, позволяющих стандартно и качественно проводить данный вид исследования, является использование высушенных питательных сред с субстратами, помещенными в лунки пластмассовых пластин для микротитрования. В них, в ходе инкубации испытуемого штамма, протекают биохимические реакции, результаты которой регистрируются визуально по изменению цвета индикатора. В большинстве тест-систем идентификации микроорганизмов время инкубации, в зависимости от вида набора, составляет 4 - 48 часов.

Основным условием правильной идентификации микроорганизмов, вне зависимости от используемого при этом метода, является наличие чистой культуры. В связи с этим отколу колонии и последующей макро- и микроскопической проверке чистоты культуры необходимо уделять особое внимание. Следует избегать «откола» сразу нескольких колоний даже при их кажущейся однотипности. Желательно использовать петлю с маленькой головкой или иглу. Не рекомендуется охлаждать петлю, прикасаясь к питательной среде без признаков роста. В отдельных случаях даже откол отдельно лежащей колонии не гарантирует получения в последующем чистой культуры. Например, колонии слизеобразующих бактерий могут содержать клетки других микроорганизмов, захваченные вместе со слизью. Культуры актиномицетов и представителей рода Bacillus могут иметь контаминанты в полостях между цепочками клеток. При раннем отборе колоний с селективной среды загрязнение может быть связано с захватом контаминантов, находящихся в подавленном, но жизнеспособном состоянии. Даже на неселективных средах откол не следует проводить слишком рано, в связи с возможным наличием медленно растущих микроорганизмов.

Работа по идентификации бактерий начинается еще до выделения чистой культуры - с изучения культуральных свойств, то есть характера роста микроорганизмов на питательной среде. Культуральные свойства бактерий, выращенных на плотной питательной среде , включают: форму, размер колоний, наличие пигмента, способность к росту при определенной температуре и концентрации углекислого газа и кислорода, на среде определенного состава и т. д. Кроме культуральных свойств еще до выделения чистой культуры иногда могут быть определены морфологические, тинкториальные свойства (например, путем микроскопии мазка из колонии, окрашенного по Граму), некоторые биохимические свойства (например, наличие ферментов оксидазы, каталазы), антигенные свойства (например, с помощью реакции агглютинации или реакции иммунофлуоресценции) и некоторые другие признаки. В отдельных случаях этого бывает достаточно для выдачи предварительного ответа о виде обнаруженного микроорганизма, но чаще всего изучение указанных признаков позволяет лишь наметить путь дальнейших исследований, которые уже выполняются с чистой культурой. Решающее значение при идентификации большинства видов микроорганизмов имеет изучение их ферментативной активности.

Выявление ферментативной активности основано на одном из пяти феноменов: Реакции на основе изменения p. H. На этом принципе основаны многие тесты, результат которых учитывается через 18 -24 час. Рост микроорганизмов на питательной среде сопровождается ферментацией содержащихся в ней субстратов. Расщепление углеводов сопровождается накоплением кислых продуктов, в то время как утилизация белковых веществ, как правило, приводит к защелачиванию среды. Реакции за счет конститутивных (предсинтезированных) ферментов. Такого рода реакции лежат в основе рапид-систем. Этот же принцип заложен в системах идентификации медленно растущих микроорганизмов или микроорганизмов, требующих особых условий культивирования (например, анаэробы в таких тест-системах могут быть идентифицированы без создания анаэробных условий инкубации). Как правило, такие тесты основаны не на изменении p. H, а на использовании хромогенных или флюорогенных субстратов.

Использование меченых источников углерода. Этот метод с использованием меченых тетразолием или радиоактивным углеродом соединений позволяет получить ответ в короткие сроки до появления видимого роста. Выявление летучих и нелетучих кислот. Метод основан на выявлении конечных продуктов метаболизма в жидкой культуре при помощи газожидкостной хроматографии. Компьютерный анализ результатов и их сравнение с библиотечными данными позволяют осуществить идентификацию культуры. Визуальное выявление роста. Этот метод используется в системах идентификации грибов и бактерий. Метод основан на выявлении способности микроорганизма к ассимиляции субстрата. Появление помутнения расценивается как положительный результат. Иногда могут быть использованы дополнительные тесты, основанные на других принципах. Существует несколько подходов к идентификации микроорганизмов. Традиционный, сложившийся еще в начале века, способ предусматривал постепенное приближение к ответу по ветвям дихотомического информационного дерева.
 тестов. Одни и те же")
. При таком подходе используется ограниченное количество пробирочных (чашечных) тестов. Одни и те же тесты могут использоваться неоднократно на разных ветвях информационного дерева. Очень важным при таком подходе оказывается ранжирование тестов по их значимости и выбор «ключевых» тестов. Это связано со следующими обстоятельствами: Изучаемый признак должен встречаться (или отсутствовать) у всех штаммов данного вида, т. е. в 100% случаев. В противном случае атипичные по этому признаку штаммы никогда не будут правильно идентифицированы. Воспроизводимость всех тестов должна приближаться к 100%. Традиционная клиническая микробиология, базировавшаяся на указанных выше принципах, была не только трудоемкой и материалоемкой, но и требовала высокой квалификации персонала. Вторая система идентификации, построенная на принципах нумерической (числовой) таксономии получила широкое распространение с начала 70 -х годов. Основные ее положения были разработаны еще в 50 -е годы. Прорыв в развитии вычислительной техники и особенно появление в конце 70 -х персональных компьютеров, а также разработка коммерческих тест-систем позволили реализовать эти положения на уровне практических лабораторий. Нумерическая таксономия базируется на следующих принципах: • 1. Необходимо изучить максимально возможное количество признаков (в современных коммерческих тест-системах их количество колеблется от 12 до 25 -30); • 2. Все изучаемые признаки имеют равный вес; • 3. Результат идентификации выглядит как коэффициент соответствия свойств выделенной культуры свойствам эталонных штаммов.

Способы изучения биохимических свойств бактерий № 5

Importance of Accurate Bacterial Identification Determine clinical significance of particular pathogen Guide physician care of patient Determine detection of antimicrobial resistance is warranted Determine appropriate type of antimicrobial therapy Determine whether susceptibility profiles are unusual Determine if organism is a risk to other patients (hospital, public or lab workers)

Среды Гисса с индикатором и углеводами: глюкозой или лактозой, сахарозой, маннитом, сорбитом, фруктозой

СИБ

Rapid Biochemical Identification? There around 20 or so different biochemical tests Fermentation tubes: many different compds as carbon source (adds to the number of possible differential tests that can be done) All these tests take time, use a lot of tubes and materials, and the results of many tests can be difficult to organize and make sense of A clinical micro lab must be able to identify samples quickly The microbiologist must do many tests in order to identify an organism accurately

Существует два основных типа таких систем. Наиболее широкое распространение получили индивидуальные системы, расчитанные на идентификацию одной культуры. К ним относятся отечественные тестситемы ПБДЭ, ПБДС (НПО «Диагностические системы» , г. Н. Новгород), семейство систем API (Bio. Merieux, Франция) и многие другие. Второй тип систем идентификации основан на использовании многорядных планшетов, в которых каждый ряд лунок позволяет идентифицировать одну культуру. Как правило, используются стандартные 96 -луночные полистироловые планшеты аналогичные тем, что применяются в иммуноферментном анализе. К системам этого типа относятся наиболее широко распространенные в нашей стране тест-системы фирмы Lachema (Чехия), а также идентификационные наборы ММТ Е (НПО «Аллерген» , г. Ставрополь). Для небольших и средних лабораторий более удобны индивидуальные тест-системы. Попытки использовать панели на 8 -12 культур в несколько приемов могут стать причиной дополнительных ошибок, так как при термостатировании происходят изменения в неиспользуемых лунках.

Идентификация выделенной культуры состоит из следующих этапов: приготовление суспензии и инокуляции лунок; инкубация; учет результатов; интерпретация.

Для приготовления суспензии, как правило, используется стерильный изотонический раствор, однако в некоторых случаях необходимо использовать специальные буферы или инокуляционные среды, входящие в состав набора. Концентрация микроорганизмов в суспензии определяется по оптическому стандарту мутности. В международной практике используется шкала Mc. Farland, в нашей стране – оптические стандарты мутности, производства Российского Государственного института медицинских и биологических препаратов им. Л. А. Тарасевича. Неправильное приготовление суспензии (слишком высокая или наоборот низкая концентрация микроорганизмов, использование старой культуры и т. д. ) может явиться причиной неправильного срабатывания системы.

Изучение биохимических свойств возбудителя путем посева на дифференциальнодиагностические среды API 20 E (Bio. Merieux Vitek Inc. , Hazelwood MO)

Manufactured Test Kits E. g. , commonly used to rapidly identify Enterics Enterotube II (BD Bioscience, Cockeysville, MD)

In hospitals more sophisticated Rapid ID systems are used often process is automated, including inoculation Automated system: more tests can be done, multiple samples can be run simultaneously, and results can even be 'read' by optical sensors and analyzed by the computer software Although this testing seems like it makes identification simple there are limitations

Limitations Many organisms don't grow well in these kinds of tests (tests rely on good growth during the incubation period) E. g. , Mycobacterium - a very slow growing organism having fastidious growth requirements A rapid ID test has not been devised for this organism - more traditional methods are still used E. g. , Strict anaerobes also more difficult to ID
тема5.ppt