синтез белка.ppt
- Количество слайдов: 65



Биосинтез белков. Регуляция транскрипции и трансляции. Повторить § 15, выучить § 16

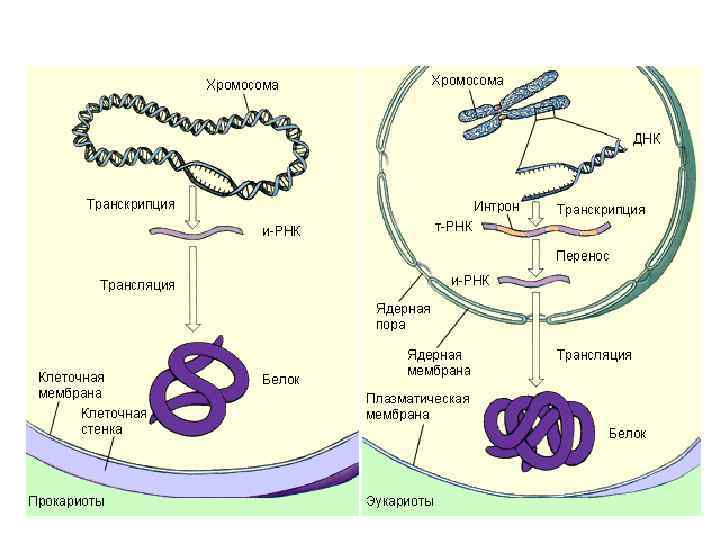
Структура гена у прокариот

Структура генома эукариот • Главной особенностью генетического материала эукариот в сравнении с прокариотами является наличие избыточной ДНК. • около 95% генома бактерий состоит из кодирующих (генных) последовательностей. • Например, у человека насчитывают приблизительно 5 х104 генов. В то же время размер генома человека - 3 x 109 п. н. Это означает, что кодирующая часть его генома составляет всего 15 -20% от всей ДНК. • Существуют виды, геном которых в десятки раз больше генома человека, например некоторые рыбы, хвостатые амфибии, лилейные растения. • Избыточная ДНК характерна для всех эукариот. • У эукариот не распространен оперонный тип расположения генов, т. е. объединение в блоки генов, находящихся под общим контролем. Гены, контролирующие даже последовательные биохимические реакции, расположены в различных районах хромосомы и даже в различных хромосомах.

Структура гена у эукариот

Этапы биосинтеза белка Биосинтез белка состоит из двух процессов: транскрипции и трансляции. Транскрипция - синтез и. РНК на ДНК, идет по принципу комплиментарности с помощью фермента РНК-полимеразы. У прокариот обычно копируется один или несколько рядом стоящих генов (ферментов, катализирующих один каскад реакций) - этот участок называется опероном. В начале оперона имеется «посадочная площадка» для фермента РНКполимеразы - эта площадка называется промотор. В транскрипции различают четыре стадии: связывание, инициация синтеза, элонгация синтеза и терминация.

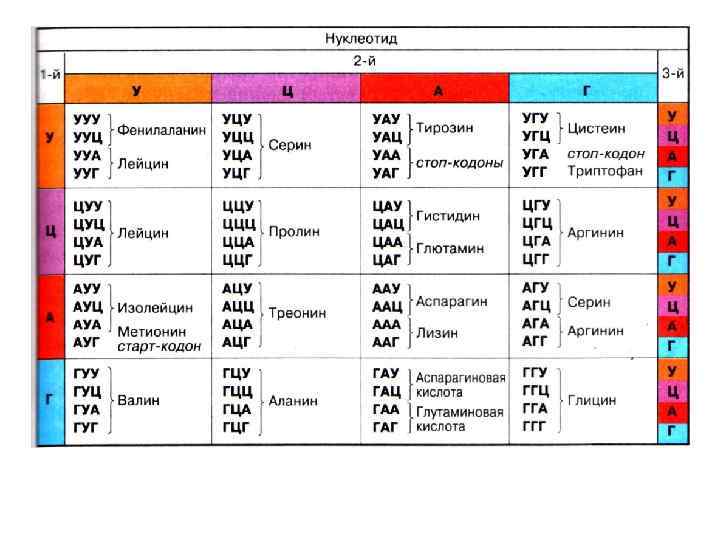
Этапы биосинтеза белка. Транскрипция ДНК АТГ АГГ ЦЦЦ ГГА АТТ ГГЦ АЦГ ЦАГ ЦЦГ ААТ АТА ТАГ ЦГЦ ЦЦА АЦЦ ТАЦ ТЦЦ ГГГ ЦЦТ ТАА ЦЦГ ТГЦ ГТЦ ГГЦ ТТА ТАТ АТЦ ГЦГ ГГТ ТГГ и. РНК АУГ АГГ ЦЦЦ ГГА АУУ ГГЦ АЦГ ЦАГ ЦЦГ ААУ АУА УАГ ЦГЦ ЦЦА АЦЦ ДНК и. РНК
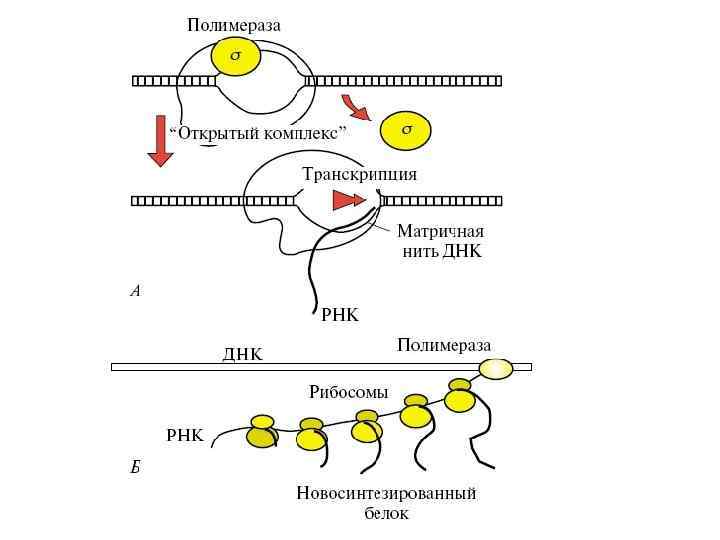

Основные правила транскрипции: 1. Процесс протекает только при наличии специфических ферментов. Основной фермент ДНК-зависимая РНК-полимераза. У бактерий одна РНК-полимераза, и все бактериальные ферменты очень похожи друг на друга. У высших организмов (эукариотов) – несколько ферментов, они называются РНК-полимераза I, РНК-полимераза III, они также имеют сходство с бактериальными ферментами, но устроены сложнее, в их состав входит больше белков. Каждый вид эукариотической РНК-полимеразы обладает своими специальными функциями, то есть транскрибирует определенный набор генов. 2. РНК-полимераза способна самостоятельно расплетать дуплекс ДНК, а затем его восстанавливать. 3. РНК-полимераза полимеризует рибонуклео-тидтрифосфаты в соответствии с последовательностью матричной цепи. Ферменту не нужна затравка – праймер.

Бактериальная РНК-полимераза изучена очень подробно. Она состоит из нескольких белковых-субъединиц: двух α-субъединиц (это маленькие субъединицы), β- и β΄-субъединиц (большие субъединицы) и ωсубъединицы. Вместе они образуют так называемый минимальный фермент, или кор-фермент. К этому кор-ферменту может присоединяться σ-субъединица. σсубъединица необходима для начала синтеза РНК, для инициации транскрипции. После того, как инициация осуществилась, σ-субъединица отсоединяется от комплекса, и дальнейшую работу (элонгацию цепи) ведет кор-фермент. При присоединении к ДНК σ-субъединица распознает участок, на котором должна начинаться транскрипция. Он называется промотор. Промотор - это последовательность нуклеотидов, указывающих на начало синтеза РНК. Без σ-субъединицы кор-фермент промотор распознать не может. σсубъединица вместе с кор-ферментом называется полным ферментом, или холоферментом.

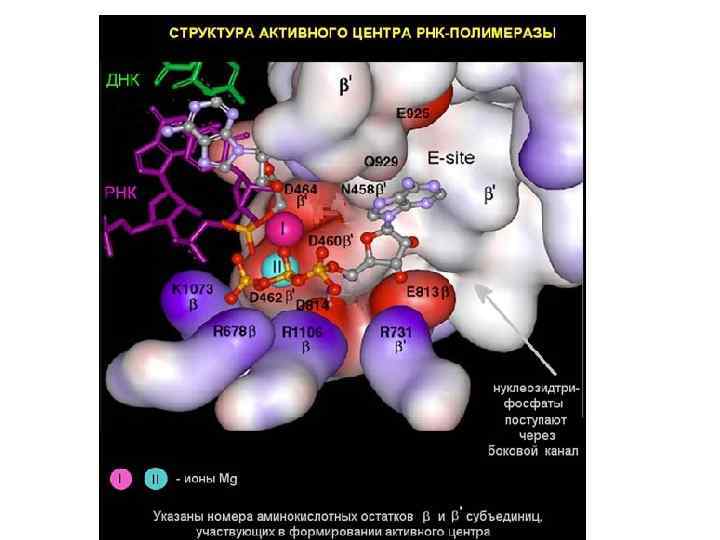
Кор-фермент состоит из α- и β-субъединиц. Они соединены так, что образуют как бы «пасть» или «клешню» . α-субъединицы находятся в основании этой «клешни» , и выполняют структурную функцию. С ДНК и РНК они, по-видимому, не взаимодействуют. ω-субъединица – небольшой белок, который также выполняет структурную функцию. Основная часть работы приходится на долю β- и β΄-субъединиц. Внутри «пасти» , которая называется главным каналом, находится активный центр фермента. Именно здесь происходит соединение нуклеотидов, образование новой связи при синтезе РНК. Главный канал в РНКполимеразе – это то место, где во время элонгации находится ДНК. Еще в этой структуре сбоку есть так называемый вторичный канал, по которому подаются нуклеотиды для синтеза РНК.

РНК-полимераза работает как молекулярная машина. Часть β΄субъединицы удерживает передний ДНК-дуплекс. Эта часть называется "заслонкой". После связывания с ДНК заслонка опускается, и зажимает ДНК так, чтобы она не могла выпасть в процессе транскрипции. Внутри находится активный центр РНК-полимеразы, где непосредственно происходит комплементарное взаимодействие поступившего по боковому каналу рибонуклеоиздтрифосфата с ДНК-матрицей. Если вновь прибывший нуклеотид комплементарен матрице, то он ферментативно пришивается к свободному 3' –концу РНК. В транслокации, то есть перемещении РНКполимеразы по нити ДНК, участвует α-спиральная структура, снизу вверх торчащая из β-субъединицы. В перемещении по ДНК участвует элемент РНКполимеразы F-спираль, которая при этом изгибается, перемещается вместе с комплексом РНК-ДНК, освобождается от них и опять выпрямляется.

В процессе транскрипции можно выделить три этапа. Первый этап - инициация транскрипции – начало синтеза нити РНК, образуется первая связь между нуклеотидами. Затем идет наращивание нити, ее удлинение – элонгация, и, когда синтез завершен, происходит терминация, освобождение синтезированной РНК.

Инициация транскрипции Инициация осуществляется с участием σ-субъединицей. Она взаимодействует со структурой ДНК, которая называется промотор. За десять нуклеотидов до точки инициации находится ТАТА-бокс последовательность для взаимодействия с σ-субъединицей. Еще примерно за 35 нуклеотидов до него находится структура, называемая « 35» . Эту последовательность также распознает σ-субъединица.
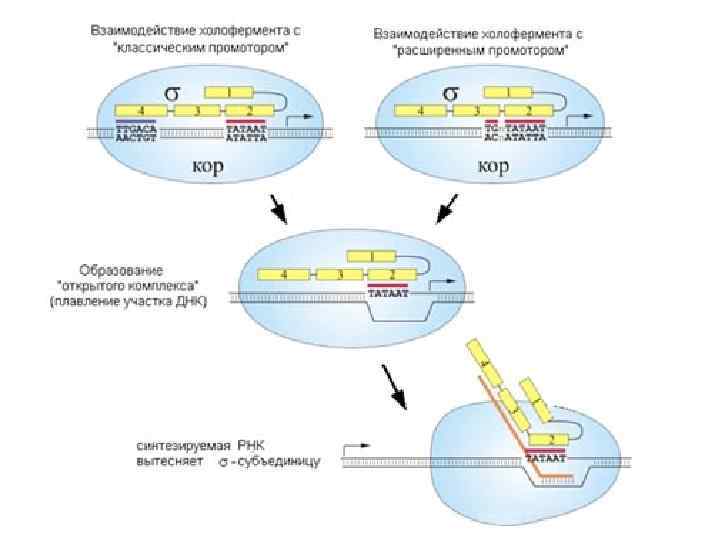

Инициация транскрипции σ-субъединица РНК-полимеразы садится на промотор в ДНК и разными частями белковой молекулы взаимодействует с частями промотора. Распознает его σсубъединица через большую бороздку ДНК. После того, как σ-субъединица в составе кор-фермента связалась с промотором, ДНК на этом участке начинает плавиться (расплетаются нити ДНК). Растущая цепь РНК выталкивает σсубъединицу и происходят еще другие изменения, которые вызывают диссоциацию σ-субъединицы от корфермента.

Элонгация транскрипции РНК наращивается на 3΄-конце. Присоединением каждого нуклеотида кор-фермент делает шаг по ДНК и сдвигается на один нуклеотид. Так как все в мире относительно, то можно сказать, что кор-фермент неподвижен, а сквозь него «протаскивается» ДНК. Понятно, что результат будет таким же. Но мы будем говорить о движении по молекуле ДНК. Размер белкового комплекса, составляющего корфермент, 150 Ǻ. Размеры РНК-полимеразы - 150× 115× 110Ǻ. То есть это такая наномашина. Скорость работы РНК-полимеразы – до 50 нуклеотидов в секунду. Комплекс кор-фермента с ДНК и РНК называется элонгационным комплексом. В нем находится ДНК-РНК гибрид. То есть это участок, на котором ДНК спарена с РНК, и 3΄-конец РНК открыт для дальнейшего роста. Размер этого гибрида – 9 пар оснований. Расплетенный участок ДНК занимает примерно 12 пар оснований.
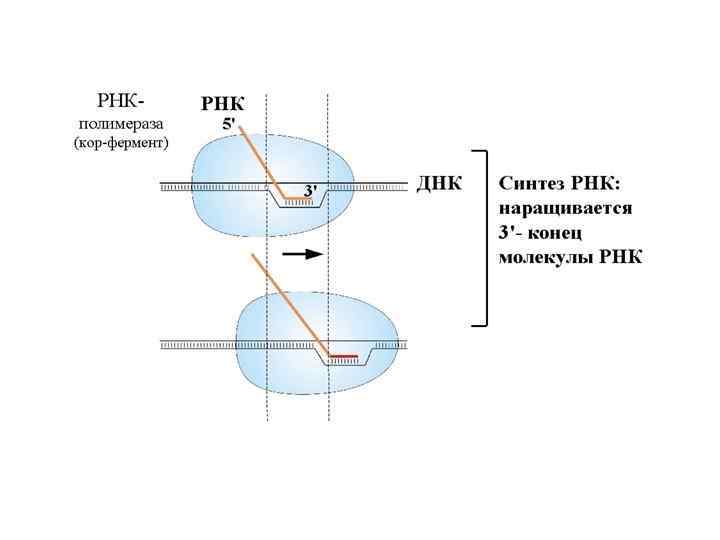

РНК-полимераза связанна с ДНК перед расплетенным участком. Этот участок называется передним дуплексом ДНК, его размер – 10 пар оснований. Полимераза связана также с более длинной частью ДНК, называемой задним дуплексом ДНК. Размер матричных РНК, которые синтезируют РНК-полимеразы у бактерий, могут достигать 1000 нуклеотидов и больше. В эукариотических клетках размер синтезируемых РНК может достигать 100000 и даже нескольких миллионов нуклеотидов.

Стадии транскрипции: • Терминатор – это последовательность ДНК, означающая окончание транскрипции, в результате чего РНК-полимераза прекращает транскрипцию и диссоциирует с матрицы. • Основным событием терминации является узнавание особой точки, в которой синтез ДНК должен завершиться. • Когда последнее основание будет присоединено к цепи РНК, транскрипционный глазок разрушается: гибридный дуплекс ДНК -РНК плавится, восстанавливается двухцепочечная структура района ДНК, фермент диссоциирует с матрицы.

Терминация транскрипции прокариот может происходить двумя способами: с участием r-фактора и без него; rнезависимая терминация обеспечивается РНК шпилькой с последующей олигоуридиловой последовательностью. На последовательности РНК должен присутствовать одноцепочечный и богатый остатками цитидина rutучасток, служащий местом посадки rфактора. Сам r-фактор представляет собой гексамерную NTPазу, способную использовать энергию гидролиза нуклеозидтри-фосфатов для продвижения по РНК в сторону ее 3’конца. Когда r-фактор “догоняет” работающую РНК-полимеразу, что также случается в местах ее временной остановки, происходит терминация. В этом процессе также участвуют ассоциированные с РНК-полимеразой факторы Nus. A и Nus. G.

Основные правила транскрипции: 1. 2. 3. 4. 5. 6. 7. Синтез РНК осуществляется ферментом РНК – полимеразой Транскрипция идет от 5’-конца к 3’-концу. Инициация транскрипции происходит только в особых участках ДНК (промотор) и строго контролируется специальными системами. Транскрипция ДНК в РНК консервативна. В результате каждой реакции образуется одноцепочечная молекула РНК, а дуплексная ДНК сохраняется без изменений. Для транскрипции необходимо локальное расплетание дуплексной структуры ДНК. Этот процесс катализирует сама РНК-полимераза. Транскрипция ДНК в РНК происходит непрерывно. В результате реакции образуется одна ковалентнонепрерывная молекула РНК. Скорость полимеризации рибонуклеотидов составляет примерно 40 нуклеотидов в секунду, что эквивалентно 15 аминокислотным остаткам в секунду.

Регуляция биосинтеза белка у прокариот А - образование открытого комплекса ДНК с РНК-полимеразой, начало транскрипции; Б – одновременный процесс транскрипции и трансляции, образование полирибосом; В – индуктор связывается с белком-репрессором, который на может тормозить синтез и. РНК Г – молекула ДНК изгибается, белок- активатор вступает в контакт с РНКполимеразой, активизируя ее.

Регуляция биосинтеза белка у эукариот

5, АТГ АГГ ЦЦЦ ГГА АТТ ГГЦ АЦГ ЦАГ ЦЦГ ААТ АТА ТАГ ЦГЦ ЦЦА АЦЦ ДНК РНК 3, 5, ТАЦ ТЦЦ ГГГ ЦЦТ ТАА ЦЦГ ТГЦ ГТЦ ГГЦ ТТА ТАТ АТЦ ГЦГ ГГТ ТГГ АУГ АГГ ЦЦЦ ГГА АУУ ГГЦ АЦГ ЦАГ ЦЦГ ААУ АУА УАГ ЦГЦ ЦЦА АЦЦ

Синтез белка у прокариот
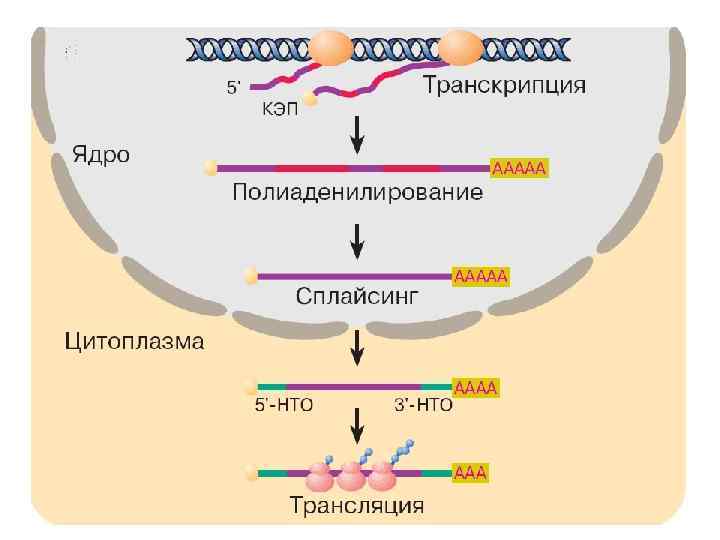
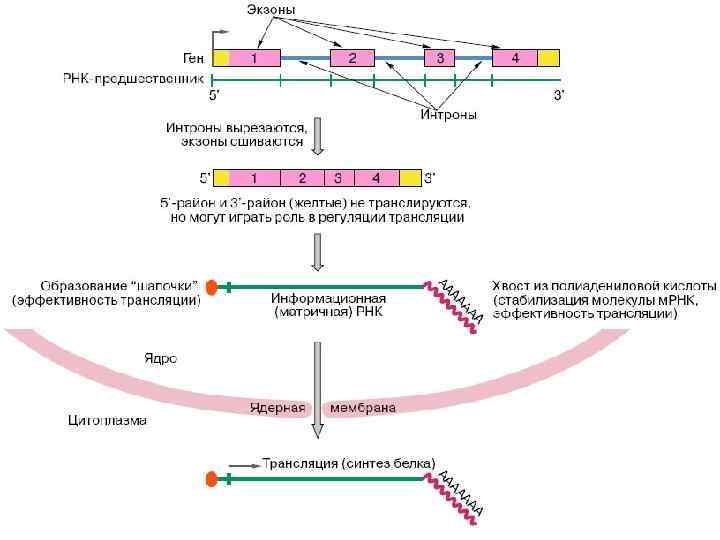

Созревание РНК Процесс «созревания» клеточных РНК сопровождается химическими модификациями РНК-предшественника и укорочением его размеров. Созревание представляет собой регулируемый многостадийный путь, по ходу которого осуществляется взаимодействие новообразованного предшественника РНК с белками и другими РНК, обеспечивающими образование функционально активной молекулы РНК. Регуляция процесса созревания РНК — это способ регуляции активности гена, кодирующего данную молекулу РНК. При созреваний информационной РНК имеет место каталитический процесс удаления из РНК интронов и объединения экзонов, кодирующих отдельные участки белка. Совокупность этих химических реакций носит название сплайсинга. Сплайсинг проходит в специальной внутриядерной многокомпонентной структуре — сплайсосоме, включающей десятки белков и набор так называемых малых ядерных РНК, обеспечивающих сплайсинг.
 приводят к образованию и. РНК, кодирующей белки с")
Разные пути сплайсинга РНК-предшественника (альтернативный сплайсинг) приводят к образованию и. РНК, кодирующей белки с разными свойствами. Красные прямоугольники обозначают экзоны, используемые при обоих путях альтернативного сплайсинга. Зеленые и синие прямоугольники обозначают экзоны, которые при альтернативном сплайсинге могут вырезаться и, следовательно, ведут себя как интроны. Линии, соединяющие экзоны над прямоугольниками и под ними, указывают разные пути сплайсинга.

• Сплайсинг осуществляется благодаря комплементарным взаимодействиям нуклеотидов РНК-предшественника и малых ядерных РНК. Образующиеся сложные структуры определяют точность сплайсинга. Кроме того, велика роль белков, способных специфично «узнавать» не только участки РНК но и друга. Сплайсинг определяется последовательными упорядоченными взаимодействиями компонентов машины сплайсинга с предшественниками РНК. • Многокомпонентность машины сплайсинга позволяет регулировать образование м. РНК за счет изменения концентраций отдельных компонентов или химической модификации, меняющей биологическую активность молекул, определяющих сплайсинг. • Изменение характера сплайсинга экзонов в разных тканях одного и того же РНК-предшественника может приводить к образованию разных РНК (альтернативный сплайсинг). В результате РНК, транскрибируемые с одного гена, будут кодировать белки с разными свойствами. • Выбор путей сплайсинга РНК-предшественника — это способ регуляции активности генов в разных клетках и тканях организма.
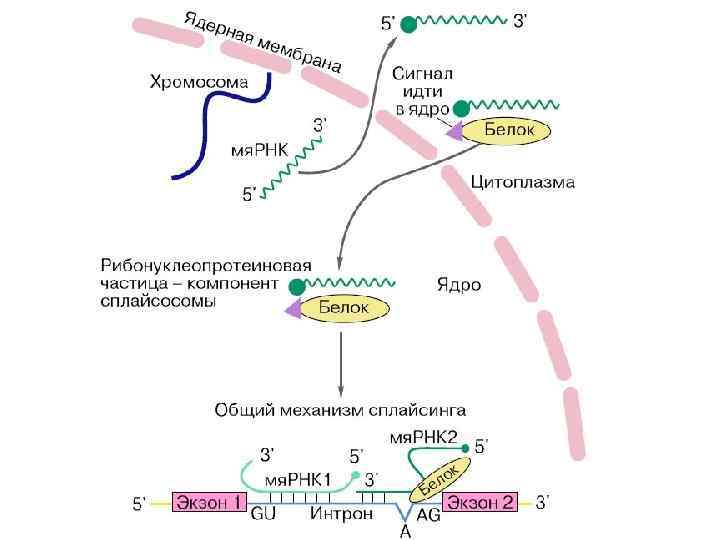

Механизм сплайсинга с участием малых ядерных РНК. Взаимодействия комплементарных нуклеотидных пар показаны штрихами. Ломаной линией показаны G-G-взаимодействия, обусловленные водородными связями.

Трансляция
 аминокислоты в полипептидную")
Основные реакции трансляции Три последовательные химические реакции приводят к включению (добавлению) аминокислоты в полипептидную цепь строящегося белка: 1) аминокислота + АТФ аминоацил-аденилат + пирофосфат, 2) аминоацил-аденилат + т. РНК' аминоацил-т. РНК' + АМФ, 3) пептидил(n)-т. РНК + аминоацил-т. РНК‘ т. РНК + пептидил(n + 1)-т. РНК'.

Трансляция – синтез белка, проводится рибосомами. Рибосома состоит из двух субчастиц: большой и малой.
. Молекулы т. РНК относительно небольшие, имеют")
В синтезе белка участвует транспортная РНК (т. РНК). Молекулы т. РНК относительно небольшие, имеют общую вторичную структуру. За счет спаривания комплементарных участков молекулы т. РНК образуется три "стебля" с петлями на концах и один "стебель", образованный 5'- и 3'-концами молекулы т. РНК. Изображение этой структуры похоже на крест или клеверный лист. "Голова" на этом листе представлена антикодонной петлей, здесь находится антикодон – те три нуклеотида, которые комплементарно взаимодействуют с кодоном в м. РНК. Противоположный антикодонной петле стебель, образованный концами молекулы, называется акцепторным стеблем – сюда присоединяется соответствующая аминокислота. Распознают подходящие другу т. РНК и аминокислоты специальные ферменты, называемые аминоацил -т. РНК синтетазами. Для каждой аминокислоты есть своя аминоацил -т. РНК синтетаза.

Этапы биосинтеза белка. Трансляция

Этапы биосинтеза белка. Трансляция

Этапы биосинтеза белка. Трансляция
. С кодоном (тремя нуклеотидами) м. РНК комплементарно")
В рибосоме находится матричная РНК (м. РНК). С кодоном (тремя нуклеотидами) м. РНК комплементарно связан антикодон транспортной РНК, на которой висит остаток аминокислоты. На рисунке видна такая структура (т. РНК вместе с аминокислотой, которая называется аминоцил-т. РНК).

Процесс трансляции, также как и процесс транскрипции, связан с перемещением вдоль молекулы нуклеиновой кислоты, разница в том, что рибосома шагает на три нуклеотида, в то время как РНК-полимераза - на один. Аминоцил т-РНК входит в рибосому, комплементарно связываясь с кодоном м. РНК, затем происходит реакция при которой аминокислотные остатки связываются друг с другом, а т-РНК удаляется.

Этапы инициации трансляции

Элонгация пептида на рибосоме а - инициаторная аминоацил-т. РНК находится в Р-участке, и первая элонгаторная аминоацил-т. РНК соответствующая следующему кодону (GUG), приходит в А-участок; б - транспептидация приводит к переносу аминокислотного остатка от инициаторной т. РНК на аминоацил-т. РНК в А-участке: образуется дипептидил-т. РНК ); в - транслокация перемещает т. РНК из Аучастка в Р-участок, и эта т. РНК увлекает за собой связанный с ней кодон м. РНК. Таким образом, м. РНК оказывается сдвинутой относительно рибосомы на один триплет нуклеотидов, и в А-участке устанавливается очередной кодон (CUG); г - аминоацил-т. РНК, комплементарная этому кодону, связывается с А-участком; д - транспептидация переносит дипептид на аминоацил-т. РНК в А-участке: образуется трипептидил-т. РНК; e - транслокация перемещает т. РНК из Аучастка в Р-участок, что приводит к сдвигу м. РНК еще на один триплет.
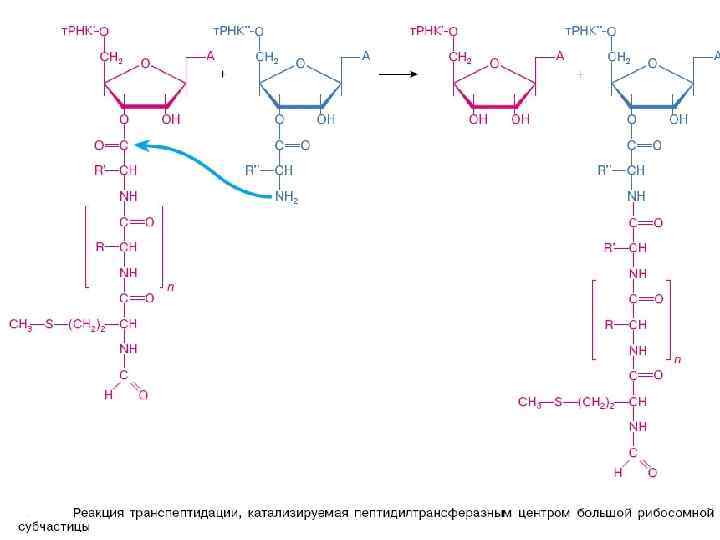

Терминация трансляции: а- после добавления последнего аминокислотного остатка к растущему полипептиду в А-участке устанавливается триплет, не кодирующий никакой аминокислоты - кодон терминации (UAG, UAA или UGA). Завершенный полипептид остается ковалентно связанным с т. РНК, которая принесла последний аминокислотный остаток в рибосому; б - А-участок с кодоном терминации воспринимает специальные белки факторы терминации RF 1 (или RF 2), непосредственно узнающий кодон терминации на малой субчастице рибосомы, и RF 3, взаимодействующий с большой субчастицей рибосомы вблизи пептидилтрансферазного центра;

е -пептидилтрансферазный центр рибосомы под действием факторов терминации катализирует реакцию переноса Сконца синтезированного полипептида от т. РНК на молекулу воды: происходит гидролиз связи между т. РНК и полипептидом и полипептид освобождается из рибосомы в среду; г - вследствие гидролиза ГТФ, т. РНК освобождается из рибосомы; д - "пустая" рибосома легко диссоциирует на субчастицы. Удалению деацилированной т. РНК и диссоциации рибосом может содействовать специальный белок -фактор освобождения (RRF)

Этапы трансляции Инициация Элонгация
 аминокислоты в полипептидную")
Основные реакции трансляции Три последовательные химические реакции приводят к включению (добавлению) аминокислоты в полипептидную цепь строящегося белка: 1) аминокислота + АТФ аминоацил-аденилат + пирофосфат, 2) аминоацил-аденилат + т. РНК' аминоацил-т. РНК' + АМФ, 3) пептидил(n)-т. РНК + аминоацил-т. РНК‘ т. РНК + пептидил(n + 1)-т. РНК'.
 из одного участка в")
Способность рибосомы перебрасывать сравнительно большие молекулярные массы (молекулы т. РНК) из одного участка в другой в каждом элементарном элонгационном цикле предполагают ее механическую подвижность. Взаимная подвижность двух рибосомных субчастиц может быть основным видом крупноблочной подвижности рибосомы в ходе ее работы.

• Существует модель, согласно которой рибосома при прохождении элонгационного цикла осциллирует между двумя конформационными состояниями: закрытым (сомкнутым) и открытым (разомкнутым). • В сомкнутом состоянии рибосомные лиганды (т. РНК) зажаты между субчастицами, связаны максимальным количеством контактов с рибосомой и не имеют внутририбосомной подвижности. • В разомкнутом состоянии рибосомы лиганды более подвижны, контакты с рибосомой менее полны, и имеется возможность их входа и выхода из рибосомы.

Этапы инициации трансляции Механизм потриплетного сканирования м. РНК в ходе элонгации предполагает участие молекул т. РНК, которые взаимодействуют прежде всего с малой рибосомной субчастицей. Малая субчастица в составе полной транслирующей рибосомы имеет два т. РНКсвязывающих участка, обозначаемых как аминоацил-т. РНК-связывающий участок (Аучасток) и пептидил-т. РНК-связывающий участок (Р-участок) В результате кодон-антикодонового взаимодействия м. РНК с т. РНК на малой субчастице рибосомы происходит декодирование триплета м. РНК: именно тот аминокислотный остаток, который был привешен к т. РНК с комплементарным антикодоном, оказался в рибосоме.

Далее молекулы пептидил-т. РНК и аминоацил-т. РНК, расположенные рядом в рибосоме, реагируют друг с другом: пептидильныи остаток переносится на аминогруппу молекулы аминоацил-т. РНК. Это второй шаг элементарного элонгационного цикла - транспептидация, когда полипептидная цепь удлиняется на одну аминокислоту — на ту, которую принесла т. РНК, связавшаяся с А-участком. А сама т. РНК, принесшая эту аминокислоту, так и осталась с ней связанной и, таким образом, связанной с удлиненным полипептидом. В этом состоянии, однако, новообразованная пептидил-т. РНК занимает "не положенный ей" А-участок, а в Р-участке "сидит" деацилированная (без пептидильного или аминоацильного остатков) т. РНК.

Элонгация пептида на рибосоме а - инициаторная аминоацил-т. РНК находится в Р-участке, и первая элонгаторная аминоацил-т. РНК соответствующая следующему кодону (GUG), приходит в А-участок; б - транспептидация приводит к переносу аминокислотного остатка от инициаторной т. РНК на аминоацилт. РНК в А-участке: образуется дипептидил-т. РНК ); в - транслокация перемещает т. РНК из А-участка в Ручасток, и эта т. РНК увлекает за собой связанный с ней кодон м. РНК. Таким образом, м. РНК оказывается сдвинутой относительно рибосомы на один триплет нуклеотидов, и в Аучастке устанавливается очередной кодон (CUG); г - аминоацил-т. РНК, комплементарная этому кодону, связывается с А-участком; д транспептидация переносит дипептид на аминоацил-т. РНК в Аучастке: образуется трипептидилт. РНК; e - транслокация перемещает т. РНК из А-участка в Р-участок, что приводит к сдвигу м. РНК еще на один триплет.

Терминация трансляции: апосле добавления последнего аминокислотного остатка к растущему полипептиду в Аучастке устанавливается триплет, не кодирующий никакой аминокислоты кодон терминации (UAG, UAA или UGA). Завершенный полипептид остается ковалентно связанным с т. РНК, которая принесла последний аминокислотный остаток в рибосому;

Задачи для самоконтроля Задача 1 Бета-лактоглобулин - белок коровьего молока имеет генетические варианты А, В и с, различающиеся по аминокислотам в позициях В и С. На других участках аминокислоты, входящие в состав белка, одинаковые. Аминокислотный состав в этих позициях следующий: ала Ала – глу – про – глу - глн - сер – лей - вариант В - цис вал вариант С позиции 111 – 112 – 113 – 114 - 115 - 116 - 117 - 118 - 119 Определите последовательность нуклеотидов в матричной и комплиментарной ей нитях молекулы ДНК в варианте В. Какие изменения произошли в последовательности нуклеотидов в молекуле ДНК в варианте С? Определите антикодоны в т-РНК.

Задача 2 Установите длину гена, кодирующего синтез белка коровьего молока беталактоглобулина, если известно, что он состоит из 350 аминокислот, а расстояние между нуклеотидами ДНК составляет 0, 34 нм. Найдите молекулярную массу гена, если средняя молекулярная масса нуклеотида равна 340 дальтон. Задача 3. В одной из цепочек молекулы ДНК нуклеотиды расположены в такой последовательности: АЦГ ТТА ГЦТ АГТ … Какова последовательность нуклеотидов в другой цепочке белка этой же молекулы? Какой аминокислотный состав имеет белок, кодируемый данной ДНК? Задача 4 Длина гена, контролирующего синтез белка составляет 335, 24 нм. Определите, сколько аминокислот входит в состав белка, если расстояние между двумя нуклеотидами составляет 3, 4 А 0. . Найдите молекулярную массу гена, если средняя молекулярная масса нуклеотида равна 340 дальтон.
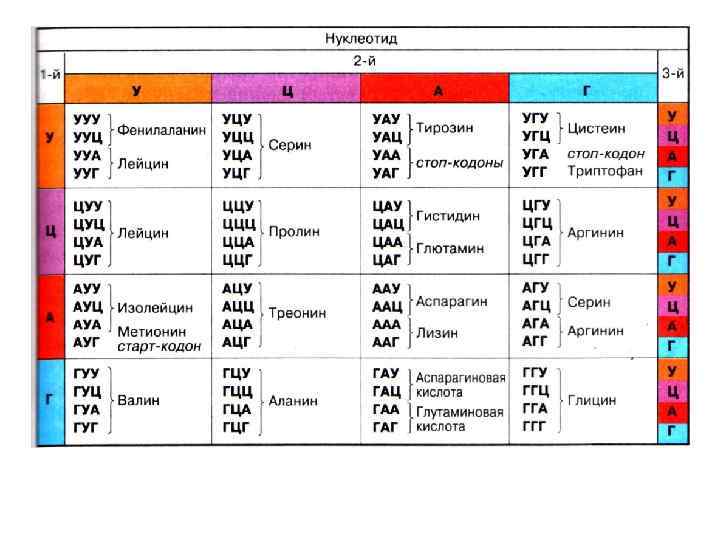

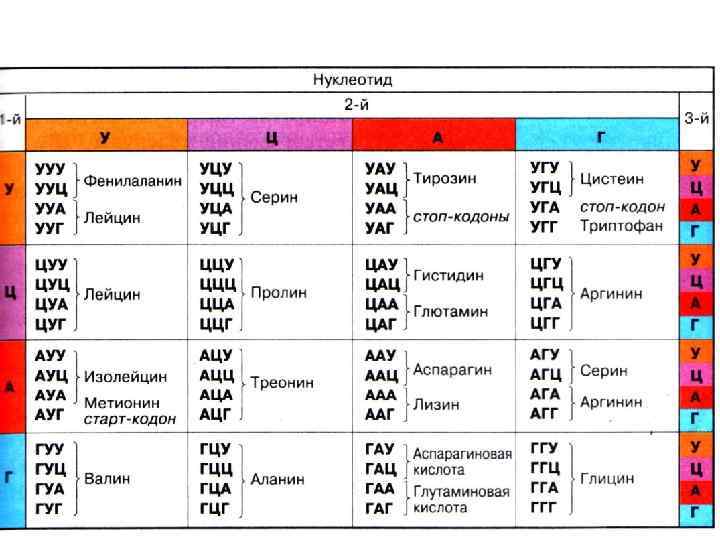
Задача 5 Укажите последовательность аминокислот в белковой молекуле, кодируемой ДНК: . . АТА ЦТГ АЦА ТТА ГАА Какой будет последовательность аминокислот, если между 10 и 11 нуклеотидом произойдет вставка гуанина? Задача 6 Длина гена, контролирующего синтез белка составляет 7799, 6 А 0. Определите, сколько аминокислот входит в состав белка, если расстояние между двумя нуклеотидами составляет 3, 4 А 0 Найдите молекулярную массу гена, если средняя молекулярная масса нуклеотида равна 340 дальтон. Задача 7 Выпишите нуклеотиды м-РНК, кодирующие белковой молекулы в следующих вариантах: а) аспарагин – аланин – тирозин- лизин; б) фенилаланин –изолейцин- валин-глицин; аминокислотный состав
 Допустим,")
Задача 8 Одна цепь участка ДНК имеет следующую последовательность оснований: ГТАГЦЦТАЦЦЦАТАГГ 3 а) Допустим, что с этой ДНК транскрибируется м-РНК, причем матрицей служит комплиментарная цепь. Какова будет последовательность и. РНК? б) Сколько пептидов кодирует эта и-- РНК? Задача 9 Одна цепь участка ДНК имеет следующую последовательность оснований: ГТАГЦЦТАЦЦЦАТАГГ 3 Допустим, что с этой ДНК транскрибируется последовательность и-РНК? м-РНК, какова будет Задача 10 При неполном окислении 38 молекул глюкозы образовалось 18 молекул молочной кислоты. Сколько молекул АТФ, кислорода и углекислого газа образовалось при окислении глюкозы?
синтез белка.ppt