2ЭМС_биомеханика.ppt
- Количество слайдов: 66



БИОМЕХАНИКА СКЕЛЕТНЫХ МЫШЦ

БИОМЕХАНИКА – Рисунок из книги Дж. Борелли De motu animalium Система рычагов, схема прикрепления мышц при сгибании в суставе и при разгибании. Скелетномышечная схема двух человек, по-разному удерживающих различный груз раздел естественных наук, изучающий на основе моделей и методов механики механические свойства живых тканей, отдельных органов и систем, или организма в целом, а также происходящие в них механические явления.

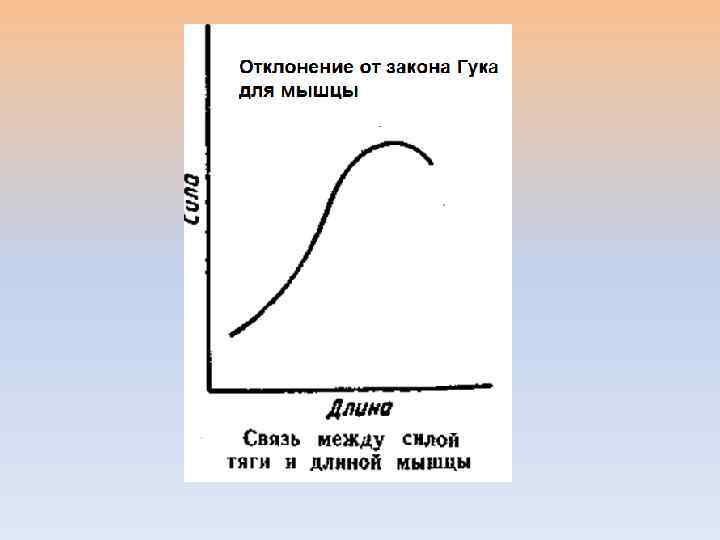
Упругое тело Гука ДЕФОРМАЦИЯ – ОТНОСИТЕЛЬНОЕ ИЗМЕНЕНИЕ ДЛИНЫ l – изменение длины l - начальная длина ЗАКОН ГУКА s- МЕХАНИЧЕСКОЕ НАПРЯЖЕНИЕ Е – МОДУЛЬ ЮНГА

Вязкая среда Остаточная деформация s - МЕХАНИЧЕСКОЕ НАПРЯЖЕНИЕ η - ВЯЗКОСТЬ

σ σ ε ε Механическая модель Фойгта, состоящая из параллельно соединённых пружины Е и поршня η в цилиндре, заполненном вязкой жидкостью. Модель Максвелла с последовательным соединением пружины Е и поршня в цилиндре η.
 1 – последовательный упругий компонент 2 – сократительный элемент")
МЕХАНИЧЕСКАЯ МОДЕЛЬ МЫШЦЫ (ПО ХИЛЛУ) 1 – последовательный упругий компонент 2 – сократительный элемент 3 - параллельный упругий компонент

СУММАЦИЯ И СЛИЯНИЕ ОДИНОЧНЫХ СОКРАЩЕНИЙ

РЕЖИМЫ СОКРАЩЕНИЯ ИЗОМЕТРИЧЕСКОЕ ИЗОТОНИЧЕСКОЕ

УСТАНОВКА ДЛЯ РЕГИСТРАЦИИ ИЗОМЕТРИЧЕСКОГО СОКРАЩЕНИЯ

1 - ОДИНОЧНОЕ ИЗОМЕТРИЧЕСКОЕ СОКРАЩЕНИЕ 2 – АКТИВНЫЙ КОМПОНЕНТ СОКРАТИТЕЛЬНОГО ЭЛЕМЕНТА ПРИ ОДИНОЧНОМ ИЗОМЕТРИЧЕСКОМ СОКРАЩЕНИИ

АКТИВНЫЕ И ПАССИВНЫЕ ХАРАКТЕРИСТИКИ СКЕЛЕТНОЙ МЫШЦЫ а – кривая длина –сила б – механическая модель мышцы 1 – кривая пассивного напряжения 2 – кривая активного изометрического напряжения

ЗАВИСИМОСТЬ НАПРЯЖЕНИЯ МЫШЕЧНОГО ВОЛОКНА И СТЕПЕНИ ПЕРЕКРЫТИЯ ПРОТОФИБРИЛЛ ОТ ИСХОДНОЙ ДЛИНЫ САРКОМЕРА

УСТАНОВКА ДЛЯ РЕГИСТРАЦИИ ИЗОТОНИЧЕСКОГО СОКРАЩЕНИЯ

ЗАВИСИМОСТЬ МЕЖДУ НАГРУЗКОЙ И ДЛИНОЙ МЫШЦЫ 1 – КРИВАЯ ПАССИВНОГО НАПРЯЖЕНИЯ 2 – КРИВАЯ ИЗОТОНИЧЕСКИХ МАКСИМУМОВ 3 – КРИВАЯ ПОСТНАГРУЗОЧНЫХ ИЗОТОНИЧЕСКИХ МАКСИМУМОВ
 - английский физиолог. Лауреат Нобелевской премии 1922 года в области")
Арчибалд Хилл (1886 -1977) - английский физиолог. Лауреат Нобелевской премии 1922 года в области медицины и физиологии за труды в области термодинамики мышечной деятельности.

а – ОДИНОЧНОЕ ИЗОТОНИЧЕСКОЕ УКОРОЧЕНИЕ ПРИ РАЗНЫХ НАГРУЗКАХ б – КРИВАЯ СИЛА - СКОРОСТЬ

УРАВНЕНИЕ ХИЛЛА P – нагрузка P 0 – максимальная нагрузка V – скорость сокращений а, b - эмпирические коэффициенты

БИОФИЗИКА МИОКАРДА

СТРУКТУРНЫЕ ОСОБЕННОСТИ МИОКАРДА
 ВОЛОКНА ПРОВОДЯЩЕЙ СИСТЕМЫ (ВОЛОКНА ПУРКИНЬЕ) КЛЕТКИ СИНОАТРИАЛЬНОГО И")
ТИПЫ КЛЕТОК КЛЕТКИ РАБОЧЕГО МИОКАРДА (КАРДИОМИОЦИТЫ) ВОЛОКНА ПРОВОДЯЩЕЙ СИСТЕМЫ (ВОЛОКНА ПУРКИНЬЕ) КЛЕТКИ СИНОАТРИАЛЬНОГО И АТРИОВЕНТРИКУЛЯРНОГО УЗЛОВ
 ВОЛОКНА ПУРКИНЬЕ КЛЕТКИ С-А И А-В УЗЛА D – 12 МКМ")
КАРДИОМИОЦИТЫ (Поперечно исчерчены) ВОЛОКНА ПУРКИНЬЕ КЛЕТКИ С-А И А-В УЗЛА D – 12 МКМ V 0. 9 – 1 М/С D – 40 МКМ V 3 М/С D – 2 – 3 МКМ V 0. 05 М/С ФУНКЦИЯ СОКРАЩЕНИЕ ФУНКЦИЯ ГЕНЕРАЦИЯ ПД ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ

ЭЛЕКТРИЧЕСКИЕ СВОЙСТВА МИОКАРДА

МИОКАРД – ЭЛЕКТРИЧЕСКИЙ СИНЦИТИЙ • Миокард является электрическим, но не морфологическим синцитием • Низкое сопротивление в области нексусов – наличие электрического синапса, имеющего коннексоны.
")
Р-КЛЕТКИ СИНОАТРИАЛЬНОГО УЗЛА ПОКАЗАНЫ ВСТАВОЧНЫЕ ДИСКИ (Н)


Возбуждение от клетки к клетке проходит без задержки.

Возбуждение распространяется за счет локальных токов, возникающих между возбужденным и невозбужденным участками.

Проведение возбуждения между кардиомиоцитами. Положительно заряженные ионы из кардиомиоцита А, в котором возник потенциал действия, через электрические синапсы переходят в смежные клетки (Б, В, Г, Д), вызывая в них деполяризацию (V)

МЕХАНИЗМ ГЕНЕРАЦИИ ПОТЕНЦИАЛОВ ДЕЙСТВИЯ РАЗЛИЧНЫМИ КЛЕТКАМИ МИОКАРДА


Ионтранспортные системы, участвующие в генерации ПД в различных клетках миокарда

ИОН-ТРАНСПОРТНЫЕ СИСТЕМЫ, УЧАСТВУЮЩИЕ В ГЕНЕРАЦИИ ПД РАЗЛИЧНЫХ КЛЕТОК СЕРДЦА

Быстрые потенциалозависимые Nа+ - каналы аналогичны Na-каналам в мембране нервного и скелетного мышечного волокна. Активируются на уровне ≈ -60 м. В, полностью инактивируются уже при деполяризации мембраны до ≈ 40 м. В. Блокируются ТТХ, однако, чувствительность их намного ниже, чем в нервных и скелетных мышечных волокнах.

МЕДЛЕННЫЕ Са 2+ КАНАЛЫ L-ТИПА ВЫСОКОВОЛЬТАЖНЫЕ: АКТИВИРУЮТСЯ ПРИ ОТНОСИТЕЛЬНО ВЫСОКИХ МП, ЗНАЧЕНИЯХ МЕДЛЕННО ИНАКТИВИРУЮТСЯ • ОСОБЕННОСТЬ: высокая чувствительность к ряду медиаторов и физиологически активных веществ, изменяющих внутриклеточный уровень ц. АМФ и ц. ГМФ.
 инактивируются")
ДВА МЕХАНИЗМА ИНАКТИВАЦИИ Са 2+каналов L-типа 1. Потенциалозависимая инактивируются деполяризацией 2. Кальцийзависимая (токозависимая) инактивируются ионами Са 2+, вошедшими в клетку во время развития этого тока.

БЫСТРЫЕ Са 2+ КАНАЛЫ T-ТИПА НИЗКОВОЛЬТАЖНЫЕ: АКТИВИРУЮТСЯ ПРИ НИЗКИХ ЗНАЧЕНИЯХ МП, БЫСТРО ИНАКТИВИРУЮТСЯ ЛОКАЛИЗАЦИЯ: ПРОВОДЯЩАЯ СИСТЕМА СЕРДЦА ОБЕСПЕЧИВАЮТ ЗАКОНОМЕРНЫЕ ПЕРИОДИЧЕСКИЕ КОЛЕБАНИЯ МП (В ПЕЙСМЕКЕРАХ)

КАЛИЕВЫЕ КАНАЛЫ

КАЛИЕВЫЕ КАНАЛЫ 1. К+- каналы утечки участвуют в формировании ПП клеток. 2. К+ - каналы аномального выпрямления инактивируются при деполяризации от больших исходных величин ПП, обеспечивают снижение К+- проводимости мембраны в начальный момент фазы 0 ПД 3. К+ - каналы задержанного выпрямления, аналогичны К+каналам в мембране нервного и скелетного мышечного волокна, обеспечивают формирование задержанного выходящего калиевого тока.

ХЕМОУПРАВЛЯЕМЫЕ ИОННЫЕ КАНАЛЫ

1. Са 2+ - активируемые К+ - каналы обеспечивают развитие кальций-индуцируемого выходящего калиевого тока По мере снижения свободного цитозольного Са 2+, каналы закрываются. 2. Nа+ - активируемые К+ - каналы. 3. АТФ - чувствительные К+ - каналы внутриклеточный АТФ ингибирует, а низкое соотношение концентраций АТФ/АДФ открывает каналы. Активность их зависит также от градиента р. Н снаружи и внутри клеток, очень слабо - от мембранного потенциала.

4. АЦХ – активируемые К+ - каналы – активируются при активации М 2 - мускариновых рецепторов кардиомиоцитов, которые через сопрягающий G-белок открывают специальные К+ каналы, противодействующие снижению калиевого тока, вызывая гиперполяризацию мембраны

СИСТЕМЫ АКТИВНОГО ТРАНСПОРТА

Na/K АТФаза
ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА")
Са 2+ АТФаза ЛОКАЛИЗАЦИЯ: САРКО- (ЭНДО)ПЛАЗМАТИЧЕСКИЙ РЕТИКУЛУМ ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА

ОБМЕННИКИ Na/Ca обменник Nа+/Са 2+ – обмен выносит ионы кальция, используя градиент к ионам натрия. При деполяризации мембраны – работает в обратном режиме, оставляя ионы кальция внутри

ПД ВОЛОКОН ПУРКИНЬЕ

ПД волокон Пуркинье Фаза 0 – быстрая деполяризация фаза 1 – ранняя быстрая реполяризация фаза 2 - «плато" фаза 3 – конечная быстрая реполяризация фаза 4 – медленная диастолическая деполяризация
")
фаза 0 - быстрый вход Na+ фаза 1 - выход K+ (или вход Cl-) фаза 2 - медленный вход Ca 2+ фаза 3 - выход K+ фаза 4 - выход K+ и вход Na+

Механизм ПД волокон Пуркинье фаза 0 - имеет овершут, активация Nаканалов и вход Nа+, 12 мсек. фаза 3 - закрытие Саканалов и выход ионов К+ до уровня ПП фаза 1 - закрытие Nаканалов, выход К+ или вход ионов Сl-. фаза 2 - (фаза плато - ПД) активация Са-каналов L-типа, вход ионов Са 2+ и Nа+. Ее длительность до 300 мсек. фаза 4 - приводит к развитию фазы 0 ПД, работа неспецифических каналов, пропускающих ионы Nа+ и К+.

ИЗМЕНЕНИЕ ПРОВОДИМОСТИ МЕМБРАНЫ ВО ВРЕМЯ ГЕНЕРАЦИИ ПД

ПД КАРДИОМИОЦИТОВ


ПД КАРДИОМИОЦИТОВ

ЭЛЕКТРОМЕХАНИЧЕСКОЕ СОПРЯЖЕНИЕ В КАРДИОМИОЦИТАХ


САРКОМЕР КАРДИОМИОЦИТА



1. ВОЗДЕЙСТВИЕ НА КАРДИОМИОЦИТ ПД СОСЕДНЕЙ КЛЕТКИ Са 2. ОТКРЫТИЕ ПОТЕНЦИАЛОЗАВИСИМЫХ КАНАЛОВ САРКОЛЕММЫ И ПОСТУПЛЕНИЕ ИОНОВ КАЛЬЦИЯ В КАРДИОМИОЦИТ 3. УВЕЛИЧЕНИЕ КОНЦЕНТРАЦИИ ИОНОВ КАЛЬЦИЯ В САРКОПЛАЗМЕ ЗАПУСКАЕТ ВЫХОД ИОНОВ КАЛЬЦИЯ ИЗ СПР 4. РЕЗКОЕ УВЕЛИЧЕНИЕ КОНЦЕНТРАЦИИ ИОНОВ КАЛЬЦИЯ В САРКОПЛАЗМЕ 5. ЗАПУСК СОКРАТИТЕЛЬНОГО АППАРАТА


ВНУТРИКЛЕТОЧНЫЕ КАЛЬЦИЕВЫЕ КАНАЛЫ НАХОДЯТСЯ В • РИАНОДИНОВЫЕ РЕЦЕПТОРЫ • IP 3 -РЕЦЕПТОРЫ СПР:


МЕХАНИЗМЫ СНИЖЕНИЯ КОНЦЕНТРАЦИИ ИОНОВ КАЛЬЦИЯ


2ЭМС_биомеханика.ppt