Лекция Т-лимфоциты МАПО_январь 2014_20_01.ppt
- Количество слайдов: 90



Биология Т-лимфоцитов: антигеннезависимое созревание Северо-Западный государственный медицинский университет им. И. И. Мечникова Кафедра клинической лабораторной диагностики 2014 год

Онтогенез Т-лимфоцитов 1. Развитие Т-клеток в тимусе 2. Развитие Т-клеток вне тимуса 3. Клиническое использование

Онтогенез Т-лимфоцитов: 1. Развитие Т-клеток в тимусе. 1. 1. Формирование антигенраспознающего репертуара 1. 2. Селекция клонов лимфоцитов 1. 3. Образование субпопуляций Тлимфоцитов

Онтогенез Т-лимфоцитов: 2. Развитие Т-клеток вне тимуса. 2. 1. Селекция Т-лимфоцитов в барьерных тканях 2. 2. Субпопуляционный состав Тклеток эпидермиса, слизистого слоя тонкого кишечника, lamina propria. 3. Клиническое использование
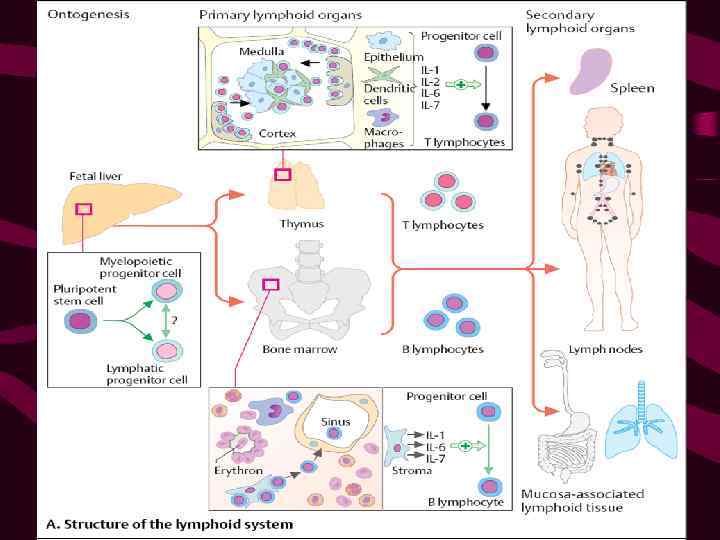
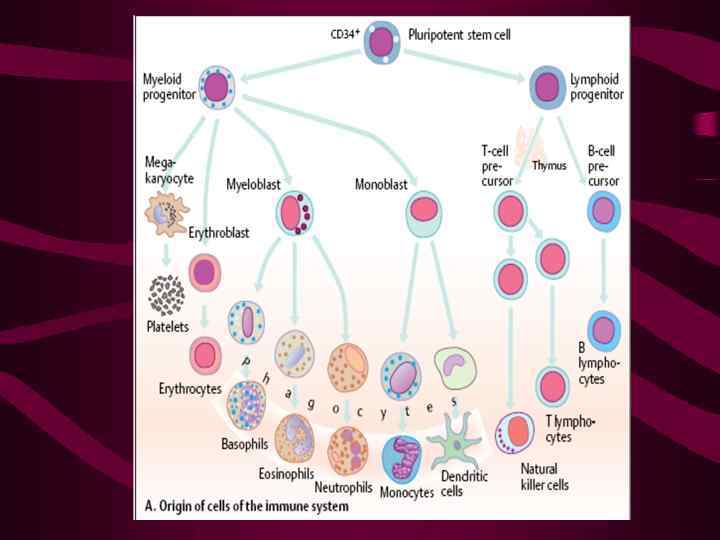
 в")
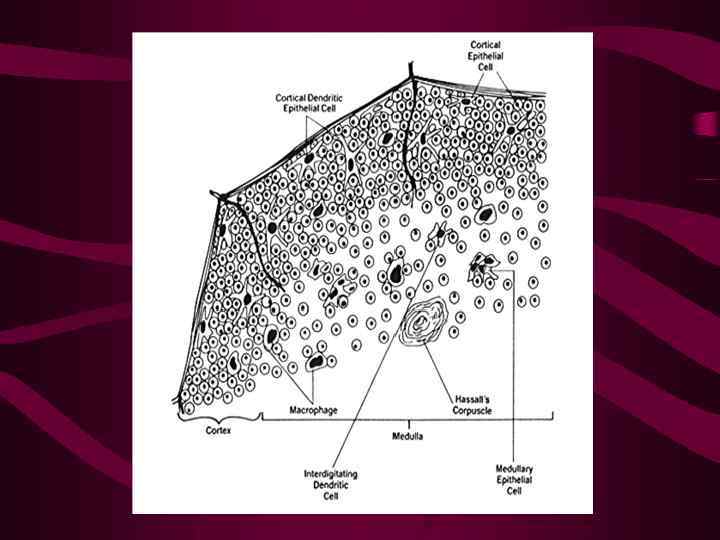
Клетки-предшественники мигрируют в тимус из кроветворных органов (печени эмбриона и костного мозга взрослых) в ответ на хемотаксические сигналы, которые обеспечиваются гуморальными продуктами ТЭК (преимущественно хемокинами)
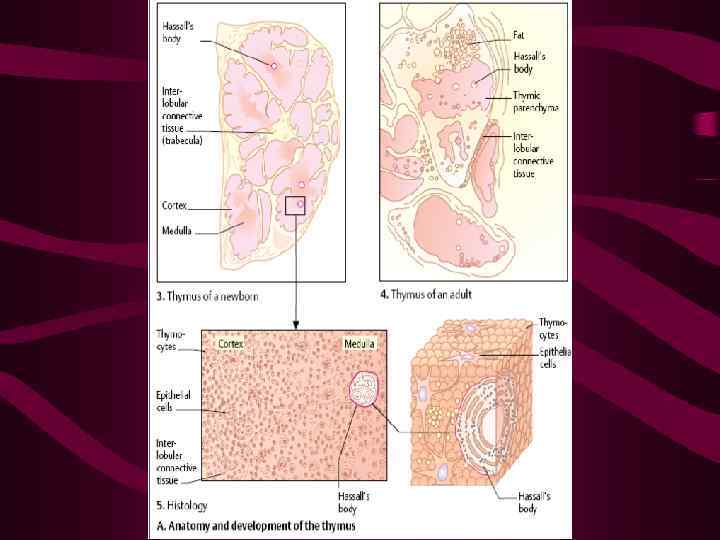

Внутреннее пространство тимуса ограничивают ТЭК Детали этих взаимодействий не изучены, известна роль молекул CD 44 и 1 интегринов, эксперссируемых тимоцитами, в качестве «рецепторов хоминга» в тимус

Этапы развития Т-клеток в тимусе Субкапсулярная зона CD 4 -CD 8(CD 44+CD 25 -CD 3 -) (CD 44 -CD 25+CD 3 -) Реанжировка -гена ТКР Экспрессия про-ТКР (CD 44 -CD 25+CD 3 lo) Реанжировка -гена ТКР (CD 44 -CD 25 - CD 3 lo) Экспрессия ТКР
 Положительная селекция (CD 4+CD")
Кортикальная зона CD 4+CD 8+ (CD 4+CD 8+CD 3 lo) Положительная селекция (CD 4+CD 8+CD 3 hi. CD 6)

Кортикально-медулярная зона и медулярная зона CD 4+CD 8+CD 3 hi CD 4+/CD 8+ CD 4 -CD 8+CD 3 hi Отрицательная селекция CD 4+CD 8 -CD 3 hi

В процессе дальнейшего развития тимоцитов наблюдаются изменения экспрессии молекул адгезии, с помощью которых они взаимодействуют с ТЭК Эти соответствия обеспечивают формирование контактов с ТЭК на всех этапах дифференцировки тимоцитов

Именно взаимодействие с ТЭК определяют основные события, связанные с развитием тимоцитов Клетки-предшественники, мигрирующие в тимус могут развиваться не только в Тлимфоциты, но и в миелоидные клетки, Влимфоциты, дендритные и НК-клетки

Их потенции сужаются по мере продвижения по стадиям развития Окончательная специализация в направлении Т-ряда устанавливается на стадии CD 44+CD 25+, когда включается наиболее ответственный процесс на пути развития Т-клеток - начинается перестройка генов антигенраспознающего рецептора, ТКР

Пусковые механизмы этого события пока не раскрыты. Показано участие в их запуске ИЛ-1, ИЛ-7 и других гуморальных факторов, но не вызывает сомнений ведущая роль (по крайней мере при формировании зрелых генов - и -цепей ТКР) неизвестных сигналов со стороны ТЭК

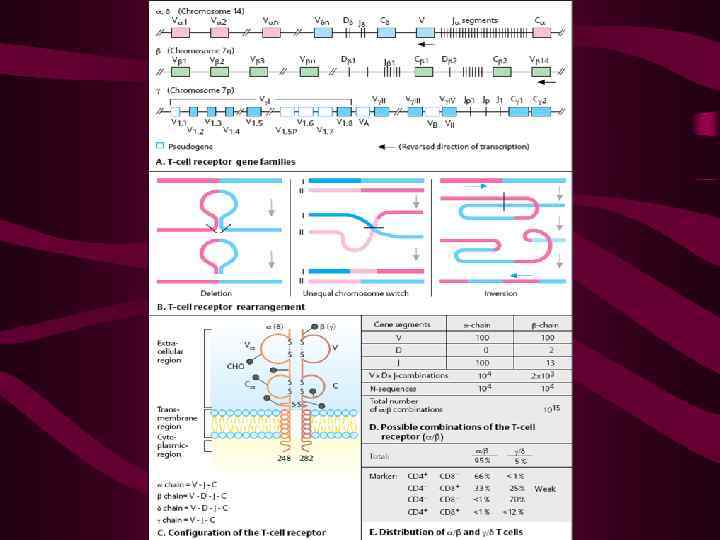
Ключевым событием при этом является экспрессия генов RAG 1 и RAG 2, катализирующие начальные этапы перестойки генов Гены различных цепей перестраиваются последовательно, перестройка - и -генов происходит практически одновременно и предшествует перестройке -гена

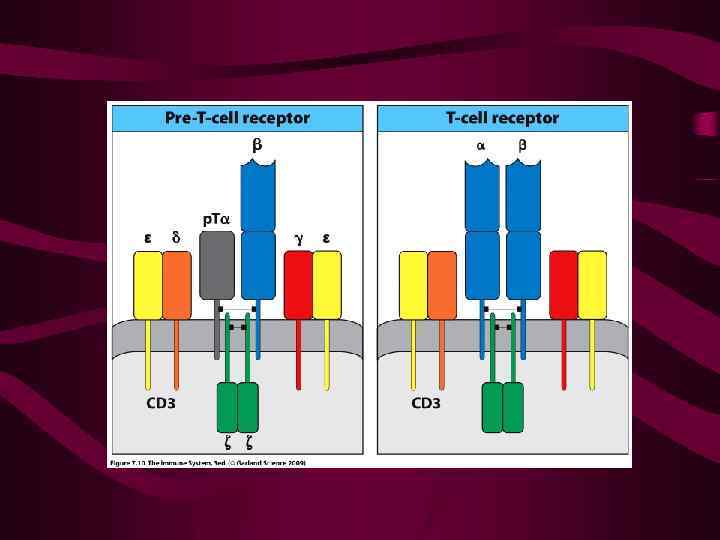
Перестройке -гена предшествует « селекция» - означающая «проверку» правильности формирования «проторецептора» , содержащего - цепь и пре- -цепь Контроль осуществляется с участием молекул ТЭК CD 81 и CD 27, и выражается в индукции пролиферации клеток с нормально экспрессированным проторецептором

Лишь после этого вновь индуцируется экспрессия RAG 1 и RAG 2 и начинается перестройка гена -цепи ТКР После ее завершения на поверхности тимоцитов экспрессируется ТКР -типа

Клетки, сформировавшие рецептор δ -типа, не нуждаются в тимусном микроокружении и рано покидают тимус -Т-клетки превращаются в CD 4+CD 8+ тимоциты, подвергаются положительной и отрицательной селекции и следующим этапам дифференцировки
 включает практически все «нужные» специфичности, т. е. рецепторы, обладающие сродством")
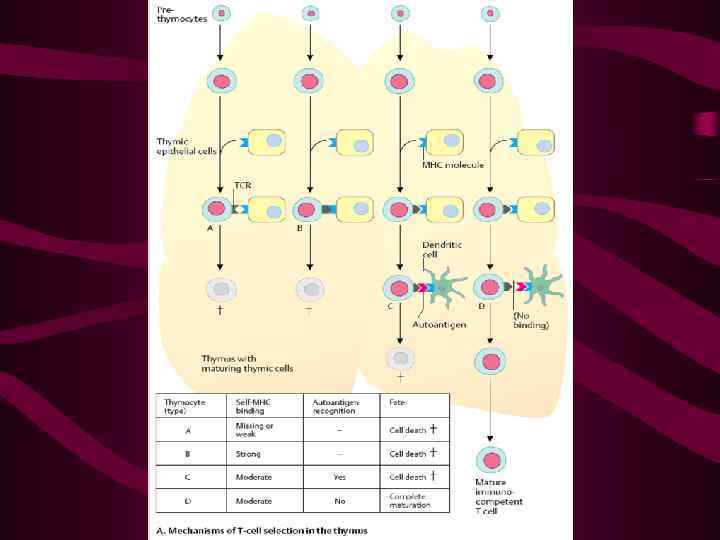
Первичный антигенраспознающий репертуар (ТКР) включает практически все «нужные» специфичности, т. е. рецепторы, обладающие сродством к молекулам МНС (ГКГС, HLA), комплексированным с пептидами Суть положительной селекции в тимусе состоит в отборе, т. е. в обеспечении выживания и пролиферации клонов тимоцитов, которые экспрессируют соответствующие ТКР (остальные клоны гибнут)

Это достигается путем «проверки» тимоцитов на способность распознавать комплексы «своих» молекул ГКГС, экспрессирующих любые пептиды Такая проверка осуществляется в форме контакта тимоцитов с кортикальными ТЭК Выжившие тимоциты затем подвергаются отрицательной селекции

На этом этапе выбраковываются тимоциты, способные распознавать «свои» молекулы МНС, экспрессирующие аутологичные пептиды Выбраковка осуществляется путем индукции апоптоза аутоспецифических клеток Выживают клетки, распознающие чужеродные пептиды в комплексе с аутологичными молекулами МНС

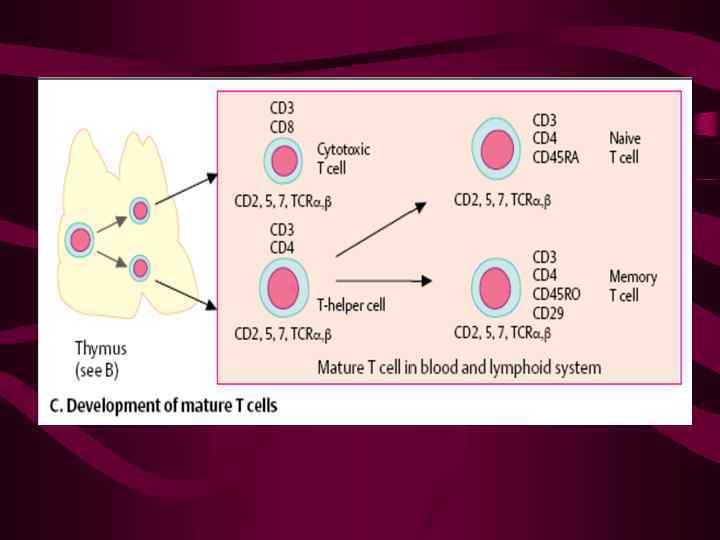
Отрицательная селекция тимоцитов и образование субпопуляций Т-лимфоцитов CD 4+CD 8+CD 3 hibcl-2 Контакт с ДК и ТЭК, способных распознавать «свои» молекулы МНС, экспрессирующие аутологичные пептиды CD 4+CD 8+CD 3+ bcl-2 Апоптоз не распознают аутологичный комплекс CD 4+CD 8+CD 3+ bcl-2+ Образование субпопуляций

CD 4+CD 8+CD 3+ bcl-2+ Контакт с медулярными МНС I / II+ ТЭК Образование субпопуляций Распознает МНС-IIпептидный комплекс Распознает МНС-Iпептидный комплекс CD 4+CD 8 -CD 3+ CD 4 -CD 8+CD 3+ Т-хелперы Т-киллеры (цитотоксические Т-лимфоциты)

CD 4+CD 8 -CD 3+ CD 4 -CD 8+CD 3+ Т-хелперы ЦТЛ мигрируют на периферию (CD 2, 1 -, 2 -интегрины, их рецепторы и другие адгезионные молекулы принимают участие во взаимодействии тимоцитов и ТЭК)

Выживают клетки, распознающие чужеродные пептиды в комплексе с аутологичными молекулами МНС Они и образуют вторичный антигенраспознающий репертуар, необходимый для защиты организма от клеток, экспрессирующих «измененное свое» , но не способных осуществлять аутоагрессию

Контроль на аутоспецифичность осуществляется в форме контакта тимоцитов с дендритными клетками или медулярными ТЭК Процесс отрицательной селекции завершается вне тимуса

С селекцией клонов тимоцитов сопряжен важный этап созревания Т-клеток, на котором происходит их разделение на субпопуляции (CD 4+CD 8 - и CD 4 -CD 8+) и который также нуждается в контакте с ТЭК Этот процесс включает два события: 1) утрату одного из двух корецепторов 2) подтверждение «правильности» этого выбора

Правильным является сохранение того корецептора, который совпадает по специфичности в отношении молекул МНС I или II корецептора с ТКР: • клетки, экспрессирующие рецептор, распознающий антигенный пептид в составе MHC-II, должны экспрессировать CD 4, обладающий сродством к МНС-II • клетки с рецептором, распознающим комплекс пептид-МНС-I, должны экспрессировать CD 8

Механизм блокады генов одного из рецепторов неизвестен (хотя и считается, что она включается сигналом со стороны ТЭК), а отбор «правильных» комбинаций ТКРкорецептор осуществляется 1) на основе контактов тимоцитов с ТЭК 2) и распознавания комплексов пептид-МНС на их поверхности

В тимусе происходит также ряд других, не менее принципиальных событий дифференцировки, которые запускаются в результате контакта с ТЭК: 1) формирование функциональных потенций хелперов и киллеров 2) экспрессия рецепторов хоминга и др. После чего созревшие Т-клетки покидают тимус
 привлечением")
Таким образом, большая часть событий, отражающих процесс развития Тклеток в тимусе, обусловлен: 1) привлечением в тимус клетокпредшественников 2) взаимодействие развивающихся клеток с ТЭК
 осуществление основных этапов дифференцировки тимоцитов 4) избирательное включение пролиферации и апоптоза тимоцитов, обуславливающее")
3) осуществление основных этапов дифференцировки тимоцитов 4) избирательное включение пролиферации и апоптоза тимоцитов, обуславливающее селекцию клонов 5) формирование антигенраспознающего репертуара
 блокада развития тимоцитов приводит к дезорганизации")
Однако и тимоциты оказывают влияние на ТЭК: 1) блокада развития тимоцитов приводит к дезорганизации эпителиального каркаса, нарушению дифференцировки и пролиферации кортикальных и медулярных ТЭК
 опустошение тимоцитов у взрослых особей (экспериментальные животные) приводит к активации ТЭК, что служит")
2) опустошение тимоцитов у взрослых особей (экспериментальные животные) приводит к активации ТЭК, что служит предпосылкой ускоренного восстановления популяции тимоцитов 3) цитокины, выделяемые тимоцитами, регулируют секреторную активность и пролиферацию ТЭК (например, тимоциты стимулируют секрецию ИЛ-6 ТЭК)
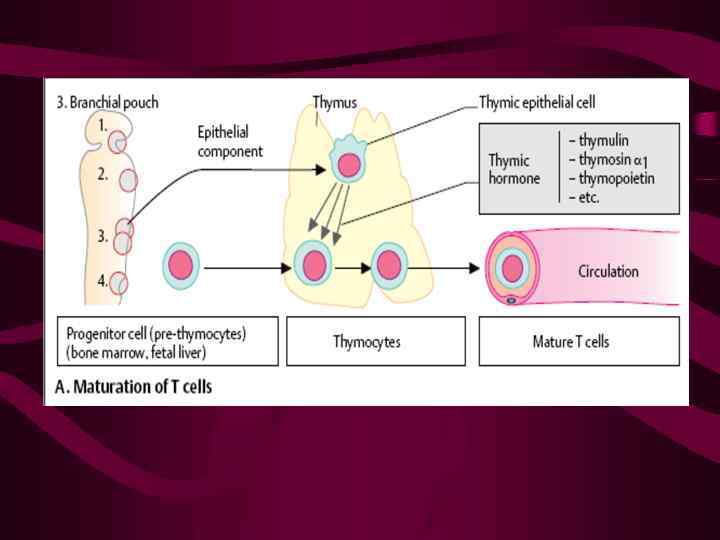
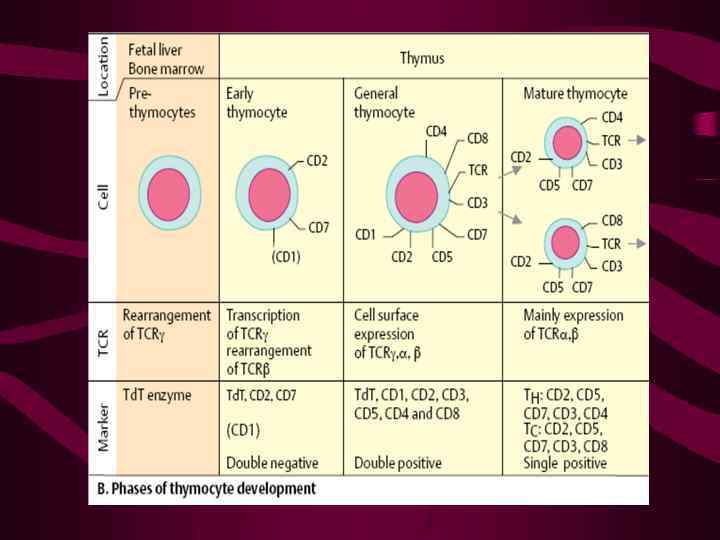


Значительно менее изучены взаимодействия зрелых Т-лимфоцитов с ЭК барьерных тканей Субпопуляционный состав Т-клеток в эпидермисе, слизистом слое тонкого кишечника и lamina propria весьма различен
 CD 3+ (Т)")
Фенотип Т-лимфоцитов в слизистом слое тонкого кишечника CD 3 -RAG+ (пре-Т) CD 3+ (Т) 10% 85% ТКР+ CD 4 -CD 810% 75% CD 4+ 7% CD 8+ 75% CD 4+CD 8+ 7% δ ТКР+ 15% CD 4 -CD 870% CD 8+… 30%

Активационные маркеры Тлимфоцитов в слизистом слое тонкого кишечника CD 69+CD 25 -CD 28 lo. CD 44+CD 62 Llo 4 7+ E 7+
 ТКР+ δТКР+ 87% 8% CD 4+")
Фенотип Т-лимфоцитов в lamina propria CD 3+ (Т) ТКР+ δТКР+ 87% 8% CD 4+ 73% CD 8+ 25% CD 4 -CD 85%

Фенотип Т-лимфоцитов в эпидермисе CD 3+ δТКР+ 10% 90% CD 4+ 50% CD 8+ 45% CD 4 -CD 885% CD 8 + 15%

Соотношение - и δ-, а также CD 4+ и CD 8+ Т-клеток в lamina propria примерно соответствует таковому в лимфатических узлах и селезенке, что отражает принадлежность всех этих клеток к рециркулирующему пулу
 высокий процент δТ-клеток")
В слизистой тонкого кишечника среди Тлимфоцитов, обозначаемых как внутриэпителиальные Т-клетки: 1) высокий процент δТ-клеток 2) большинство имеют фенотип CD 4 -CD 83) одновременно часть Т-клеток экспрессирует гомодимерную форму CD 8 ( ), что свидетельствует о внетимусном происхождении этих клеток
 среди Т-клеток преобладают CD 8 + лимфоциты 5) велик процент CD 4 -CD")
4) среди Т-клеток преобладают CD 8 + лимфоциты 5) велик процент CD 4 -CD 8 - и CD 4+CD 8+ клеток
 доля δТ-клеток также достаточно велика и среди них содержатся CD 8")
В эпидермисе 1) доля δТ-клеток также достаточно велика и среди них содержатся CD 8 + лимфоциты внетимусного происхождения 2) среди Т-клеток CD 4+ лимфоциты сильно преобладают над CD 8+ клетками

Особенности состава популяций Т-лимфоцитов эпидермиса и слизистой оболочки тонкого кишечника могут быть объяснены двумя процессами: 1) избирательностью миграции в эти участки организма Т-клеток тимусного происхождения 2) внетимусным (возможно локальным) развитием Т-лимфоцитов

Cвидетельства избирательной миграции: 1. Поступление в эпидермис CD 4+ клеток, экспрессирующих CLA+ E 7+ 2. CD 8+ клетки, обладающие сродством к слизистой тонкого кишечника, также имеют определенные фенотипические особенности: 2. 1. экспрессируют интегрин E 7+

2. 2. Лишены 2 - интегрина, LFA 1 и костимулирующей молекулы CD 28 2. 3. Имеют “активированный” фенотип (CD 69+CD 25 -CD 44+CD 62 L-) Хемотаксические сигналы осуществляют преимущественно хемокины МСР-1 и МIP-1

δ Т-клетки

δ и αβ Т-клетки

Развитие Т-клеток вне тимуса В слизистой кишечника обнаружены морфологические структуры, в которых осуществляется развитие Тклеток Ими оказались криптопатчи - скопления незрелых (CD 3 -TCR-Ig. M-) лимфоцитов, контактирующих с ЭК слизистой

Развитие Т-клеток вне тимуса CD 3 -CD 4 -CD 8 -CD 45 R+Thy-1+ IL-7 R+ Cлизистая кишечника (криптопатчи) Печень δ ТКР+CD 3 V 5 Миграция в легкие ТКР+CD 3 CD 8 +Fc. RI- V 4/ V 6 Слизистая кишечника ТКР+CD 3 СD 4 -CD 8 -СD 45 R+ ?

Формирование криптопатчей зависит от ИЛ-7 В криптопатчах образуются δ Т-клетки изотипов V 5 и V 4/Vδ 6 и Т-клетки, экспрессирующие CD 8 и -цепь Fc. RI V 5 -клетки мигрируют в слизистую бронхолегочного аппарата, а другие Т-клетки остаются в кишечнике или мигируют в слизистые оболочки других органов

Есть сведения, что в каждом криптопатче формируются клетки одного клона, причем в разных криптопатчах (а их число у мышей составляет около 1500) клоны отличаются по специфичности и в сумме они формируют ограниченный антигенраспознающий репертуар

Известен еще один участок организма, в котором происходит дифференцировка Тклеток в контакте с ЭК. Это синусоиды печени В синусоидах печени образуются клетки фенотипа CD 4 -CD 8 - c промежуточным уровнем экспрессии рецепторного комплекса TRC-CD 3 Однако об этом пути внетимусной дифференцировки известно еще меньше, чем о развитии Т-клеток в криптопатчах

Клетки, формирующиеся вне тимуса, не подвергаются селекции в той форме, как этот процесс осуществляется в тимусе Однако в барьерных тканях происходят процессы, приводящие к селекции и формированию клонов с антигенраспознающим репертуаром, который можно назвать третичным

Суть этой формы селекции состоит в том, что Т-клетки, первично активированные бактериальными антигенами в пейеровых бляшках и дифференцировавшиеся в лимфатических узлах, в процессе рециркуляции возвращаются в кишечник и повторно испытывают действие тех же антигенов Результатом является накопление в организме Т-клеток памяти, специфичных к антигенам, с которыми организм реально контактирует в повседневной жизни

Формирование этого «актуального» третичного репертуара происходит во всех барьерных тканях параллельно с атрофией тимуса В определенной степени этот процесс делает излишней деятельность тимуса по формированию «необязательных» клонов.
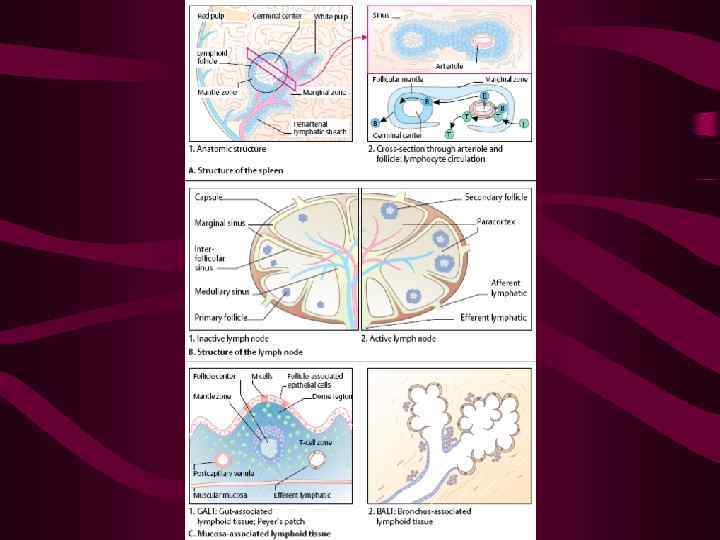

Зачем нужен тимус? События, происходящие в тимусе, и их уникальность

В тимусе происходят ключевые события развития Т-лимфоцитов: • формирование и экспрессия Т-клеточного рецептора (ТКР, TCR) • cелекция клонов тимоцитов • разделение единой линии Т-клеток на субпопуляции

CD 3 -CD 4 -CD 8 экспрессия генов рекомбиназ RAG-1 и RAG-2 под влиянием ИЛ-7 перестройка генов , и δ TCR (ТКР антигенраспознающего рецептора)

Суть перестройки состоит в сближении генетических сегментов V, D и J и формировании зрелого V- гена Клетки делают выбор между дифференцировкой в сторону лимфоцитов, экспрессирующих рецепторы типов TCR и TCR δ В случае выбора TCR δ клетки покидают тимус

При выборе - пути на поверхности появляется предшественник рецептора с цепью и пре-TCR -цепь в сочетании с комплексом CD 3, а также CD 4 и CD 8 Такие клетки размножаются (подвергаются « -селекции» ) После нескольких делений происходит перестройка -гена TCR

После перестройки -гена экспрессируется полноценный рецептор TCR- Перестройка генов происходит в клетке случайно и независимо, и на каждой Т-клетке появляется уникальный по специфичности рецептор

На уровне целой популяции тимоцитов формируется набор рецепторов, способный распознавать практически все мыслимые конфигурации молекул, который составляет первичный антигенраспознающий репертуар

Зрелые Т-лимфоциты, как известно, распознают чужеродные пептиды в комплексе с аутологичными молекулами главного комплекса гистосовместимости - МНС (от англ. Major Histocompatibility Complex). Для того, чтобы привести первичный репертуар в соответствие с этой потребностью, клоны тимоцитов подвергаются двухэтапной селекции

На этапе положительной селекции в результате контакта с эпителиальными клетками стромы тимуса отбираются клоны тимоцитов, распознающие любые пептиды в комплексе с аутологичными молекулами ГКГ (МНС-молекулами) Клетки, соответствующие этому условию, защищаются от апоптоза и пролиферируют, остальные клетки гибнут вследствие развития апоптоза

На этапе отрицательной селекции выбраковываются потенциально опасные клоны, распознающие комплексы аутологичных пептидов с аутологичными молекулами МНС, которые представлены на поверхности ДК тимуса В случае, если такое распознавание произошло, в клетку поступает сигнал к апоптозу, и она гибнет, тогда как остальные выживают и продолжают развитие

Параллельно с селекцией популяция незрелых CD 4+CD 8+ тимоцитов разделяется на субпопуляции зрелых Т-клеток, несущих корецепторы CD 4 или CD 8 (но не оба вместе) Корецептор CD 4 обладает сродством к молекуле МНС II класса Корецептор CD 8 - к молекуле МНС I класса В случае расхождения специфичностей рецептора и корецептора клетка гибнет

Таким образом, • все рассмотренные события специфичны для тимуса • основа уникальности процессов, осуществляющих в тимусе, заключается в уникальном сочетании, создающем «конвеер» , который обеспечивает оптимальную пространственную организацию процесса развития Т-клеток

• практически все клеточные и гуморальные факторы тимуса в той или иной форме представлены в периферическом отделе иммунной системы

ТЭК продуцируют: Цитокины: ИЛ-7, ИЛ-1 , ИЛ-3, ИЛ-6, ФНО , ГМ-КСФ, Г-КСФ, М-КСФ, ИФН , ТФР 1 Пептиды: тимулин, 1 -тимозин, тимопоэтины, ростовой гормон, окситоцин, вазопрессин, пролактин, релизинг фактор, ростовой фактор нервов, соматотонин и др.

Тимус является органом, относящимся одновременно к двум системам - иммунной и эндокринной: • ТЭК вырабатывают пептидные субстанции, часть которых поступает в циркуляцию и обеспечивает поддержание на должном уровне функциональной активности Т-лимфоцитов • эндокринная функция тимуса взаимосвязана с функциями других желез

• эндокринная функция тимуса взаимосвязана с функциями других желез: - на выработку тимулина оказывает стимулирующее действие соматотропный гормон (СТГ), пролактин, гормоны щитовидной железы, половые гормоны - регуляция 1 -тимозина, тимопоэтинов изучается

• изучается влияние на выработку гормонов тимуса цитокинов, которые в отличие от гормонов являются по преимуществу близкодействующими факторами и многие из них образуются в тимусе и участвуют в регуляции выработки гормонов его ТЭК • имеются сведения о стимулирующем действии на выработку тимулина интерлейкина-1

Следующая лекция Онтогенез Т-лимфоцитов антигензависимое созревание

Контроль знаний настоящей лекции

Онтогенез Т-лимфоцитов: 1. Развитие Т-клеток в тимусе. 1. 1. Формирование антигенраспознающего репертуара 1. 2. Селекция клонов лимфоцитов 1. 3. Образование субпопуляций Тлимфоцитов

Онтогенез Т-лимфоцитов: 2. Развитие Т-клеток вне тимуса. 2. 1. Селекция Т-лимфоцитов в барьерных тканях 2. 2. Субпопуляционный состав Тклеток эпидермиса, слизистого слоя тонкого кишечника, lamina propria. 3. Клиническое использование
Лекция Т-лимфоциты МАПО_январь 2014_20_01.ppt