Презентация лекции 5_БО_Мх и ОФ.ppt
- Количество слайдов: 42


 лекции Т. В. ЖАВОРОНОК доцент кафедры биохимии и молекулярной биологии")
Биологическое окисление (тканевое дыхание) лекции Т. В. ЖАВОРОНОК доцент кафедры биохимии и молекулярной биологии Сиб. ГМУ

• Клетки превращают питательные вещества в клеточные компоненты и извлекают энергию из окружающей среды при помощи высоко интегрированной системы химических реакций, называемой «обмен веществ» или «метаболизм» . • Здесь «вещества» – это метаболиты. Все процессы жизнедеятельности сопровождаются переносом веществ и энергии, которые тесно связаны между собой: 1) Пластические процессы – структурное обеспечение клеток и организма (биомембраны, рост…) 2) Энергетические процессы – обеспечение энергией Метаболизм основан на высоко интегрированной мультиферментной системе

ВИДЫ ОБМЕНА ВЕЩЕСТВ • Внешний включает процессы ассимиляции, т. е. поглощения химических соединений из внешней среды (белки, жиры, углеводы, соли, О 2, Н 2 О…) • Промежуточный (тканевой) состоит из 2 фаз: 1) катаболизм, в котором происходит расщепление сложных органических молекул до простых конечных продуктов; при этом высвобождается энергия, заключённая в сложной структуре больших органических молекул. На определённых этапах эта энергия реализуется (трансформируется): • в макроэргические связи остатков фосфорной кислоты молекул АТФ; • в восстановительные эквиваленты (запас в виде НАДФН·Н), обеспечивающие процессы синтеза.
 анаболизм, в котором из малых молекул-предшественников (строительных блоков) синтезируются сложные органические макромолекулярные компоненты")
2) анаболизм, в котором из малых молекул-предшественников (строительных блоков) синтезируются сложные органические макромолекулярные компоненты клеток (белки, нуклеиновые кислоты, жиры, углеводы), что сопровождается расходом энергии. Эти 2 фазы протекают в клетках одновременно и связаны веществом и энергией: (Б, У, Л) специфические катаболизм АТФ (~Р) анаболизм низкомолекулярные продукты энергия в-ва НАДФН·Н молекулы-предшественники

• АТФ – молекула, богатая энергией, поскольку её трифосфатный компонент содержит две фосфоангидридные связи. При гидролизе АТФ→АДФ+Фн высвобождается большое количество свободной энергии ΔG = 7, 3 ккал/моль • АТФ – главный непосредственный ДОНОР свободной энергии, а не форма запасания энергии. Весь АТФ расходуется в клетке в течение 1 минуты • Чтобы получить необходимые ежедневно среднестатистическому человеку 2500 ккал нужно 40 кг АТФ, а в организме всего 50 г = АТФ+АДФ+АМФ → каждая молекула АТФ в сутки тысячи раз фосфорилируется и дефосфорилируется, что снижает её время жизни → постоянный синтез АМФ
 I стадия")
Существует 3 стадии генерирования энергии при окислении питательных веществ: Белки углеводы липиды(жиры) I стадия (специфи- 20 АК глюкоза глицерол+СЖК ческая) ДОАФ пируват (СН -СО-СООН) II стадия СО 2 (неспецифи- 3 ацетил~Ко. А (СН -СО~SКо. А) 3 ческая) III стадия ЦТК ẽẽ 2 СО 2 (ФАДН 2 3 НАДН·Н) дыхательная цепь митохондрий (общий этап обмена Б, Ж, У) О 2+2 H+→Н 2 О

• I стадия – превращение полимеров в мономеры, осуществляют пищеварительные ферменты. • II стадия – превращение мономеров в ключевые метаболиты (ацетил. Ко. А и пируват), одинаковые вещества образуются в разных видах обмена, за счёт них обменные процессы взаимосвязаны. • III стадия – ЦТК и окислительное фосфорилирование. Представляют собой конечные общие пути в окислении топливных молекул. ЦТК – циклическая последовательность из 8 реакций, начинается с ОА. Тканевое дыхание (биологическое окисление) – процесс превращения Б, У, Ж в аэробных условиях до конечных продуктов СО 2 и Н 2 О, а энергия химических связей этих соединений реализуется в виде макроэргических связей остатков Н 3 РО 4 молекул АТФ. СО 2 образуется в ПДГ-комплексе и ЦТК (две реакции декарбоксилирования), Н 2 О образуется в конце дыхательной цепи при переносе ẽẽ и H+ на кислород.
 заканчивается: 1) переносом части окисляемых H+")
• Аэробное декарбоксилирование пирувата СН 3 СОСООН(окислительное) заканчивается: 1) переносом части окисляемых H+ на НАД с образованием НАДН·Н → в дыхательную цепь; 2) переносом образуемой в результате удаления СО 2 уксусной кислоты на коэнзим А с образованием Ас~Ко. А → в ЦТК. • Большинство топливных молекул вступают в ЦТК после превращения в Ас~Ко. А. В ЦТК идёт катаболизм (распад) Ас~Ко. А до 2 СО 2, часть энергии аккумулируется в виде ГТФ→АТФ за счет субстратного фосфорилирования, а 4 H+ переносятся на 1 молекулу ФАДН 2 и 3 молекулы НАДН·Н и поступают → в дыхательную цепь. • ЦТК выполняет ещё и анаболическую функцию – поставляет промежуточные продукты для биосинтезов: - для глюкозы (в процесс глюконеогенеза), - для аминокислот (в процесс переаминирования)

СН 3 -СООН 2 С → 2 СО 2 4 Н → 4 дегидрогеназы ЦТК (1 ФАДН 2 и 3 НАДН·Н) СН 3 -СО~SКо. А

НАДН·Н и ФАДН 2 - богатые энергией молекулы, т. к. содержат по 2ẽ с высоким потенциалом переноса: у НАДН·Н редокс-потенциал=-0, 32 В (первичный донор дыхательной цепи), у молекулы О 2 редокс-потенциал=+0, 82 В (конечный акцептор цепи), остальные компоненты дыхательной цепи занимают промежуточное положение. Перепад энергии используется для генерирования АТФ. Процесс образования АТФ сопряжён с транспортом ẽ по цепи переносчиков от НАДН·Н или ФАДН 2 к О 2, называется «окислительное фосфорилирование» .
 • Палладин.")
• БАХ. В окислительных процессах принимает участие активный кислород (О 2) • Палладин. Окисление субстратов идёт за счёт дегидрирования, т. е. без непосредственного участия О 2, который присоединяется только на последнем этапе • Механизм протекания процесса окислительного фосфорилирования предложил П. Митчел, сформулировав его в виде хемиосмотической теории

МЕЖМЕБРАННОЕ ПРОСТРАНСТВО МАТРИКС КРИСТЫ ВНУТРЕННЯЯ МЕМБРАНА

Основная идея процесса: 1. Окислительное фосфорилирование идёт на внутренней мембране митохондрии. 2. ЦТК протекает в матриксе митохондрий и поставляет большую часть НАДН·Н и ФАДН 2. 3. Окисление и фосфорилирование – процессы сопряжённые, характеризуются отношением Р/О (это число молекул неорганического фосфата, включающегося в органические формы в расчёте на 1 атом потребляемого О 2). При окислении НАДН·Н индекс Р/О=3, т. е. НАДН·Н даёт 3 молекулы АТФ. При окислении ФАДН 2 индекс Р/О=3, т. е. ФАДН 2 даёт 2 молекулы АТФ.

4. Работают дыхательные ансамбли, включающие в себя многочисленные переносчики ẽ. Многоступенчатый транспорт ẽ от НАДН·Н и ФАДН 2 к О 2 способствует выбросу H+ из матрикса митохондрии в межмембранное пространство и генерированию мембранного потенциала (протон-движущей силы). H+ перекачиваются 3 -мя видами ẽ-переносящих комплексов. В процессе обратного тока H+ в митохондриальный матрикс через трансмембранный ферментный комплекс АТФ-синтетазу происходит синтез АТФ.
: I – НАДН·Н - коэнзим Q")
В дыхательной цепи митохондрий работает 4 комплекса (ансамбля): I – НАДН·Н - коэнзим Q II – сукцинат(ФАДН 2) - коэнзим Q III – коэнзим Q - цитохром с - редуктазный IV – цитохром с - оксидазный АТФ-синтазную систему считают V комплексом.
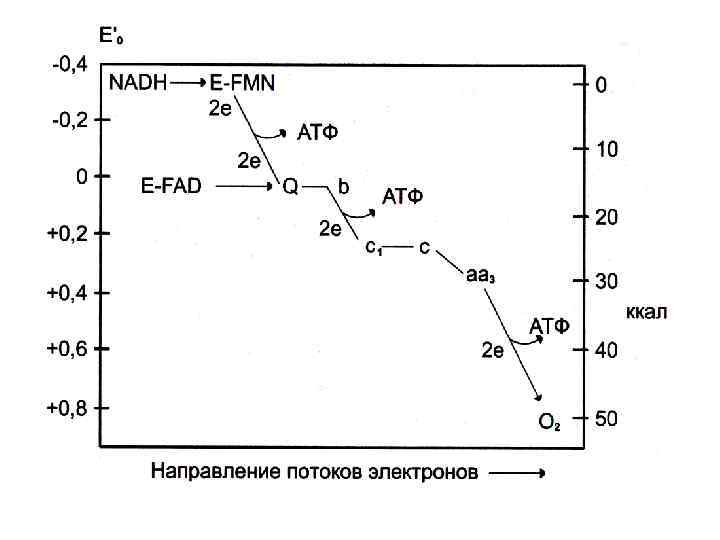

матрикс митохондрии межмембранное пространство Схема дыхательной цепи митохондрий Далее механизмы её работы более подробно
 С НАДН протоны и электроны поступают на ФМН.")
I комплекс (НАДН·Н - коэнзим Q) С НАДН протоны и электроны поступают на ФМН. На ФМН. H 2 происходит разделение H+ и ẽ: перенос H+ на другую сторону мембраны, а ẽ идут через железосерные комплексы (железо-серные белки – это негемовые железопротеины) на коэнзим Q II комплекс (сукцинат(ФАДН 2) - коэнзим Q) • H+ H+ • • I • ФМН. H 2 ẽẽ ФМН Fe. S II Q---QH 2 ẽẽ • • НАДН·Н ФАДН 2 В итоге работы первых двух комплексов ẽ через флавопротеины поступают на Ко. Q и следуют далее на комплексы цитохромов

• Электроны при своём окислительном движении вдоль внутренней мембраны митохондрий трижды (в 3 -х комплексах: I, III, IV) пересекают мембрану от внутренней поверхности к наружной и при этом в межмембранное пространство выбрасывается по 2 H+. Комплексы цитохромов переносят только ẽ, используя Fe. S-белки. • H+ H+ III H+ H+ IV

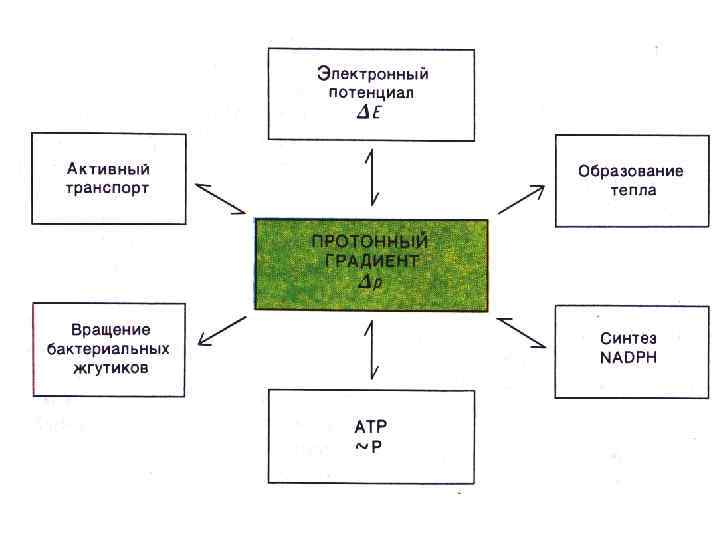
В результате переноса ẽ по дыхательной цепи идёт выброс H+ из матрикса в межмембранное пространство, где концентрация Н+ возрастает. • Протонный градиент, образованный окислительным транспортом электронов, представляет собой резервуар свободной энергии. Теперь, если обеспечить поток Н+ обратно сквозь мембрану, можно получить энергию.

Энергия накапливается в протонном градиенте за счет 2 компонентов: • 1 -й компонент – разница в концентрации ионов H+ на противоположных сторонах мембраны. Концентрационный градиент протонов обозначают как дельта р. Н (Δ р. Н). Энергия концентрационного градиента не зависит от абсолютной величины р. Н. Она обусловлена разницей р. Н по обе стороны мембраны и составляет около 60 мэ. В. (Δ р. Н = 60 мэ. В).

• 2 компонент Протон Н+ является заряженной частицей. Поэтому 2 компонент энергии протонного градиента - это градиент электрического заряда Н+. Движение зарядов через мембрану создает разницу в электрическом потенциале. В итоге возникает электростатическое поле, которое действует на все заряженные частицы. Этот компонент обозначают как мембранный потенциал - дельта пси (Δ ), он составляет 160 -180 мэ. В.

Общая энергия протонного градиента, т. е. разность электрохимического потенциала протонов – это сумма концентрационного (или осмотического) и электрического компонентов: Δ Н = Δр. Н + Δ = 230 -240 мэ. В • Таким образом, основной смысл хемиосмотического механизма в том, что первичным актом, запасающим энергию, является перенос протонов через внутреннюю мембрану митохондрии

• Сопряжение тканевого дыхания и окислительного фосфорилирования определяется уникальными свойствами внутренней мембраны митохондрии. Она плотнее наружной мембраны, поэтому 1) имеет высокое электрическое сопротивление 2) выдерживает высокое электрическое напряжение • Мембрана находится под огромным
 - подаче")
• Повреждение мембраны по любой причине - при гипоксии (недостатке кислорода) - подаче большого количества О 2 - активации перекисного окисления липидов - набухании митохондрий ведет к пробою мембраны. В этом случае Н+ не могут накапливаться в межмембранном пространстве, и митохондрия не выполняет своей главной функции – производства энергии и снабжения этой энергией всей клетки.
 на механическую работу (вращение жгутиков, движение сперматозоидов, диапедез")
Пути расходования энергии Δ Н 1) на механическую работу (вращение жгутиков, движение сперматозоидов, диапедез лейкоцитов) 2) на осмотическую работу (перенос частиц через мембраны – например ионов Са 2+ в матрикс митохондрий или секреция ионов Н+ в полость желудка при образовании соляной кислоты) 3) на теплообразование (механизм см. в конце лекции) 4) на образование АТФ. В виде АТФ энергия используется для внутриклеточных процессов. Всего в 3 -х точках дыхательной цепи создается достаточный уровень энергии ΔμН+, поэтому только за счёт этих 3 -х точек происходит образование АТФ
. • АТФ-синтетаза состоит из 2 -х")
V комплекс Синтез АТФ осуществляет митохондриальная АТФ-аза (АТФ-синтетаза). • АТФ-синтетаза состоит из 2 -х частей: 1 часть – сопрягающий фактор 1 (F 1). F 1 - это белок с массой в 360 к. Да, содержит 5 видов полипептидных цепей. Физиологическая роль этого компонента состоит в катализе синтеза АТФ. 2 часть – протонный канал (F 0) АТФ-синтезирующего механизма
 Н+ Н+ Н+ ТРАНСЛОКАЗА Н+ Н+ стебелек АДФ +")
протонный канал комплекса (F 0) Н+ Н+ Н+ ТРАНСЛОКАЗА Н+ Н+ стебелек АДФ + Рi = АТФ АДФ сопрягающий фактор (F 1)

Гипотеза П. Митчелла: ток Н+ непосредственно действует на реакцию синтеза АТФ. Сначала свободная фосфатная группа связывается с ферментом в активном участке F 1 -компонента комплекса, (вблизи конца протонного канала через Fо). По каналу под действием мембранного потенциала и градиента р. Н переходят два Н+. Эти два Н+ воздействуют на один из кислородов фосфата и соединяются с ним, образуя молекулу воды. Фосфатная группа при этом становится активной и легко связывается непосредственно с АДФ, образуя АТФ.

протонный канал комплекса или F 0 Н+ Н+ Н+ ТРАНСЛОКАЗА Н+ Н+ стебелёк ТРАНСЛОКАЗА АДФ + Рi = АТФ АДФ сопрягающий фактор 1 или F 1
 на осмотическую работу; • 2) на механическую работу; •")
Пути расходования АТФ • 1) на осмотическую работу; • 2) на механическую работу; • 3) на химическую работу: а) аллостерический регулятор, б) пластический материал для синтеза нуклеиновых кислот, коферментов, в) источник ц. АМФ, г) источник фосфатных групп для образования фосфорных эфиров, д) источник энергии для процессов синтеза.

ДЫХАТЕЛЬНЫЙ КОНТРОЛЬ. • Скорость дыхания митохондрий, а, следовательно, и скорость образования энергии – регулируемый процесс. • Скорость может контролироваться концентрацией АДФ. Если в митохондриях весь АДФ→АТФ, то это ведёт к торможению переноса ẽ в дыхательной цепи. Это обусловлено тем, что окисление и фосфорилирование жестко сопряжены, т. е. функционирование дыхательной цепи по окислению Н+ и ẽ не может осуществляться, если оно не сопровождается фосфорилированием АДФ.

Окисление и фосфорилирование могут быть не только сопряжены, но и разобщены или заингибированы ГРУППЫ ИНГИБИТОРОВ ДЫХАТЕЛЬНОЙ ЦЕПИ И ОКИСЛИТЕЛЬНОГО ОСФОРИЛИРОВАНИЯ 3 группы: 1) Ингибиторы собственно дыхательной цепи 2) Ингибиторы окислительного фосфорилирования 3) Разобщители окислительного фосфорилирования
 ИНГИБИТОРЫ, БЛОКИРУЮЩИЕ ДЫХАТЕЛЬНУЮ ЦЕПЬ действуют в 3 местах, соответствующих трем комплексам дыхательной цепи:")
1) ИНГИБИТОРЫ, БЛОКИРУЮЩИЕ ДЫХАТЕЛЬНУЮ ЦЕПЬ действуют в 3 местах, соответствующих трем комплексам дыхательной цепи: • Первый участок блокирования – в 1 -ом комплексе дыхательной цепи, где ингибиторы препятствуют НАД-зависимому окислению субстратов. Ингибиторы: барбитураты (амобарбитал), ротенон (очень токсичен; американские индейцы добывали его из растений и использовали в качестве яда при ловле рыб).

• Второй участок блокирования дыхательной цепи локализован в 3 -м комплексе между цитохромами b и с. Ингибитором является антибиотик антимицин А • Третий участок блокирования – 4 -й комплекс, т. е. цитохромоксидаза, локализованная в конце цепи. Ингибируется классическими ядами – сернистым водородом окисью углерода цианидом
 ИНГИБИТОРЫ, ПОДАВЛЯЮЩИЕ СТАДИЮ ФОСФОРИЛИРОВАНИЯ: • антибиотик олигомицин, который полностью блокирует окисление и фосфорилирование")
2) ИНГИБИТОРЫ, ПОДАВЛЯЮЩИЕ СТАДИЮ ФОСФОРИЛИРОВАНИЯ: • антибиотик олигомицин, который полностью блокирует окисление и фосфорилирование в интактных митохондриях за счет того, что не пропускает протоны через канал F 0. • Атрактилозид блокирует окислительное фосфорилирование за счёт ингибирования фермента транслоказы, производящей поступление АДФ в митохондрию и выход АТФ из митохондрии.
 РАЗОБЩИТЕЛИ отличаются тем, что нарушают в дыхательной цепи систему сопряжения процессов окисления и")
3) РАЗОБЩИТЕЛИ отличаются тем, что нарушают в дыхательной цепи систему сопряжения процессов окисления и фосфорилирования. • В этих условиях процесс дыхания происходит неконтролируемым образом, т. е. концентрации АДФ и Фн в этом случае не являются лимитирующими. • Разобщители обычно легко растворимы в липидном слое мембран, этим объясняется их разобщающее действие
 Разобщители увеличивают проницаемость митохондриальной мембраны для протонов (протонофоры), тогда не может")
ТИПЫ РАЗОБЩЕНИЯ 1) Разобщители увеличивают проницаемость митохондриальной мембраны для протонов (протонофоры), тогда не может набраться достаточно водородных ионов для создания необходимой ΔμН+, и энергия рассеивается. К этому типу относят динитрофенол и другие нитро- и галогенные производные фенола (связывают Н+ и переносят через мембрану), некоторые кислые продукты метаболизма, продукты ПОЛ. 2) Разобщители сами проникают через мембрану митохондрии и нейтрализуют заряд протонов, чем снижают ΔμН+. К ним относят многие антибиотики.

РАЗОБЩИТЕЛИ БЫВАЮТ ЭКЗОГЕННЫМИ И ЭНДОГЕННЫМИ • лекарственные вещества – строфантин, папаверин, салицилаты • естественные метаболиты организма: - белок термогенин бурой жировой ткани (у страдающих ожирением бурого жира мало) - тиреоидные гормоны - жирные кислоты Эндогенные разобщители важны для согревания организма на холоде (см. далее механизм согревания)

• Снижение температуры тела возбуждает терморецепторы, что ведёт к повышению выделения гормона норадреналина. В результате увеличивается липолиз и повышается содержание жирных кислот. • Активируется бета-окисление жирных кислот и увеличивается количество ацетил-Ко. А, что способствует возрастанию активности цикла Кребса, повышению содержания НАДН и ФАДН 2, поступающих в дыхательную цепь, где растёт активность тканевого дыхания и увеличивается электрохимический потенциал

• Однако сами жирные кислоты снижают ΔμН+ за счёт сброса Н+ в обход V комплекса, создающего АТФ, поэтому энергия ΔμН+ рассеивается, расходуясь на согревание. При этом на холоде человек дрожит, дополнительно идёт рефлекторное возбуждение терморецепторов. • При дрожании снижается содержание АТФ и увеличивается АДФ, которые регулируют активность цитратсинтазы, запускающей ЦТК. В ЦТК растёт концентрация НАДН и ФАДН 2, что повышает разность электрохимического потенциала, расходуемого на согревание. • Итог: человек согревается
Презентация лекции 5_БО_Мх и ОФ.ppt