Биолог. часы_2012.ppt
- Количество слайдов: 52



БИОЛОГИЧЕСКИЕ ЦИРКАДИАННЫЕ ЧАСЫ

Вопросы 1. 2. 3. 4. Зачем нужны часы живым организмам? Теории внешних и внутренних часов. Суточные и циркадианные ритмы. Требования к механизму биологических часов. 5. Поиск внутриклеточных часов в биохимических колебательных процессах. 6. Открытие генов биологических часов. 7. Генетические модели часов у Drosophila melanogaster и у млекопитающих.

Гибель клеток Спектр поглощения нуклеиновых кислот Спектр поглощения белков 240 260 280 300 320 нм

Зачем живым организмам нужны часы?

1. Для согласования процессов жизнедеятельности со сменой дня и ночи. 2. Для приспособления к смене сезонов. 3. Обитателям приливных зон для учета времени приливов и отливов. 4. Для решения задач ориентации и навигации по Солнцу или звездам. 5. Для согласования разнообразных процессов в клетке и сложном многоклеточном организме.
 Нерест строго увязан с фазами Луны: он происходит через один-два дня")
АТЕРИНА-ГРУНИОН (Leuresthes tenuis) Нерест строго увязан с фазами Луны: он происходит через один-два дня после новолуния или полнолуния, в периоды сигизийных приливов. Живет у побережья Калифорнии.

Многодневная динамика физиологических функций человека

Теория внешних часов Теория внутренних часов
, который способен воспринимать внешние ритмические")
Сторонники внешних часов: • Организм имеет механизм (типа маятника), который способен воспринимать внешние ритмические воздействия и только под их влиянием в часовом устройстве возникают колебания. • Собственных колебаний (автоколебаний) у него нет. Сторонники внутренних часов: • У живых организмов часы устроены по типу генератора, имеющего свои собственные колебания — автоколебания. • Роль внешних датчиков времени заключается в регуляции точности хода этих часов.

Основной аргумент сторонников теории внешних часов Относительная автономность периода суточных ритмов по отношению к обменным процессам : • независимость периода суточных ритмов от температуры. • устойчивость периода суточных ритмов к большинству химических воздействий, даже к ядам.

Доводы сторонников эндогенности биологических часов: 1. При постоянных условиях освещения и температуры происходят небольшие измене- ния периода ритма при повышении температу-ры организма, а также при воздействии на организм тяжелой водой (D 2 O), этиловым спиртом (C 2 H 5 OH) и т. д. Если бы ритм был навязан неконтролируемыми факторами среды, этого бы не наблюдалось. 2. В постоянных условиях освещения и температуры суточный ритм у растений и животных сохраняется. 3. Явление циркадианности.

• Яйца, личинки и куколки дрозофил от момента зачатия содержались в полной темноте и при постоянной температуре. • В этих условиях ритмы куколок не синхронизированы, поэтому время выхода каждой дрозофилы из куколки свое и не привязано ко времени суток. Если бы в термостат проникал неконтролируемый синхронизирующий геофизический фактор, этого не могло бы быть. • Достаточно единственной вспышки света, чтобы синхронизировать «внутренние часы» : через каждые 24 часа после вспышки будет происходить массовое вылупление взрослых мух из куколок (белые точки).
 в подземном свето-")
Эксперименты немецкого физиолога Юргена Ашоффа с изоляцией человека (1962 г. ) в подземном свето- и звуконепроницаемом бункере 1. Биологические часы имеют эндогенную природу. 2. В изоляции от датчиков времени время человека идёт медленнее реального
 ритм - циклическое колебание интенсивности того или иного биологического")
• Циркадианный (околосуточный ) ритм - циклическое колебание интенсивности того или иного биологического процесса с периодом в пределах 20 - 28 часов. • Околосуточные эндогенные ритмы под синхронизирующим воздействием 24 -часовых ритмов среды становятся 24 -часовыми.

Требования к механизму биологических часов 1. 2. Для измерения времени нужен какой-то периодический процесс. Что это за процесс? В основе механизма часов должен быть процесс с периодом колебаний порядка секунд. 3. Ход часов не должен зависеть от температуры. 4. В биологических часах должно быть устройство, преобразующее суточный ритм в низко- и высокочастотные. 5. Механизм часов должен настраиваться по внешним ритмам и по сигналам, идущим от других клеток организма.

Поиск внутриклеточных часов в биохимических колебательных процессах

• Первым исследователем, которому удалось достоверно зарегистрировать биохимические колебания, был Ауфдемгартен. • Он обнаружил (1939 г. ) периодические (с периодом около минуты) колебания скорости газообмена в процессе фотосинтеза.

Выдающийся американский биохимик Бриттен Чанс открыл периодический биохимический процесс в анаэробном гликолизе • Глюкоза + 2 ADP+2 Pi 2 ATP+2 H 2 O 2 Лактат - +2 H+ + • Глюкоза + 2 ADP+2 Pi 2 Этанол + 2 СО 2 +2 ATP+2 H 2 O (брожение) • Фруктозо-6 -фосфат Фосфофруктозокиназа Фруктозо-1, 6 - дифосфат В этой реакции были обнаружены колебания скорости

Если добавить в воду, где живут эвглены и гониаулаксы метаболические яды, останавливающие дыхание и гликолиз, жизнедеятельность этих организмов замирает: эвглены перестают двигаться, гониаулаксы не могут генерировать свет. Но часы у них идут.

• Если добавить в воду яды, отравляющие процессы считывания генетической информации, например, актиномицин-Д, препятствующий функционированию РНК–полимеразы, (обеспечивает синтез м. РНК по матрице ДНК) ход часов нарушается. • Эти наблюдения сделаны американским исследователем Дж. Гастингсом в 70 годы XX века. • Результаты этих опытов послужили толчком для поиска механизма часов в реакциях синтеза белка.

Первые доказательства генетической природы биологических часов Первый в мире часовой ген !!!
 и Сеймур Бензер (справа) открыли ген Per (period) в Х -хромосоме")
Рон Koнoпka (слева) и Сеймур Бензер (справа) открыли ген Per (period) в Х -хромосоме дрозофилы в 1971 г.

Ритм суточной активности дрозофил Локализация гена Per на генетической карте дрозофилы 19 ч 24 ч 29 ч Активность гена циклически меняется с периодом ч. Per ~ 24 Мутации этого гена приводят к наследуемым изменениям хода часов: • аннулируют ритмичность • удлиняют , • укорачивают • циркадианный ритм.

Суточные осцилляции гена Per в легких мыши
 группа исследователей из Национального")
• Двадцать лет спустя ( в начале 1990 -х) группа исследователей из Национального центра биологического времени (Северо. Западный университет, штат Иллинойс, США) под руководством Джозефа Такахаши впервые идентифицировала у мышей • часовой ген Сlock - circadian locomotor output cycles kaput - «циркадный прерыватель циклов двигательной активности» .

• Самцам мышей вводили N-этил-Nнитрозилмочевину — вещество, взаимодействующее с молекулой ДНК и вызывающее случайные мутации. • Под действием этого мутагена у некоторых животных в сперматозоидах возникли мутации, которые проявились у потомства этих животных. • Мутантов распознавали по длительности и устойчивости циркадианного ритма двигательной активности в беличьих колесах при разных условиях освещенности. • • У здоровых мышей среднее значение циркадного ритма составляло 23, 7 0, 17 ч, а у мутантов — 24, 8 ч. • Затем была определена локализация часового гена Clock, отвечающего за длительность и устойчивость циркадного ритма мыши.
. // Cell,")
ЛИТЕРАТУРА • Dunlap J. C. Molecular Bases for Circadian Clocks ( Review). // Cell, 1999. Vol. 96. P. 271 – 290. • Panda S. , Hogenesch J. B. , Kay S. Circadian rhythms from flies to human. // Nature, 2002. V. 417. P. 329 - 335.
 Per 1, Per 2, Per")
Основные часовые гены Дрозофила Мышь Per - Period (период) Per 1, Per 2, Per 3 Сlk – Сlock Сlk - Сlock - circadian locomotor output cycles kaput (циркадный прерыватель циклов двигательной активности) Tim - Timeless (безвременный) Cyc –Cycle (цикл) Bmal 1 (Вrain and Muscle Arnt-like) Cry – Сryptochrom (криптохром, кодирует белок фоторецептора) Cry 1 –Cry 2 - Сryptochrom Dbt – Doubletime (кодирует белок, гомологичный CKl - кодирует казеинкиназу

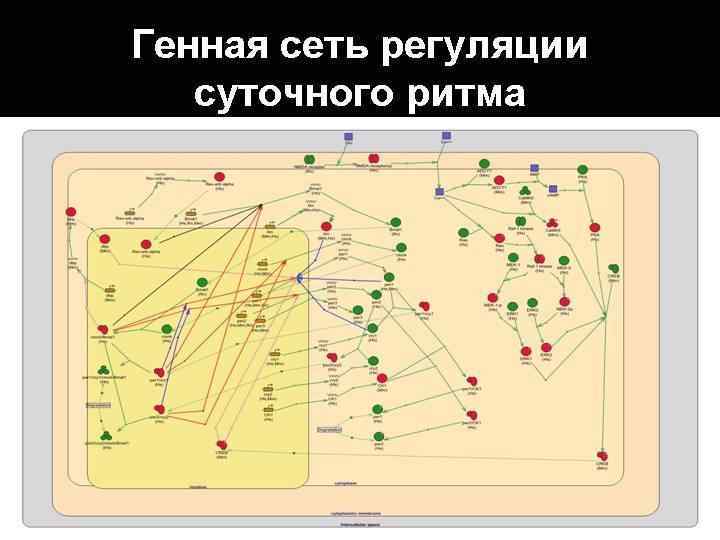
Принцип работы часового механизма у разных организмов от прокариот до человека одинаковый: циркадианные гены организованы в транскрипционно-трансляционную петлю, в которой регуляция осуществляется по принципу обратной связиобразуя цикл с , периодом около 24 ч.

Транскрипционнотрансляционную петлю часового механизма принято делить на два блока: 1. транскрипционный -положительный 2. трансляционный -отрицательный

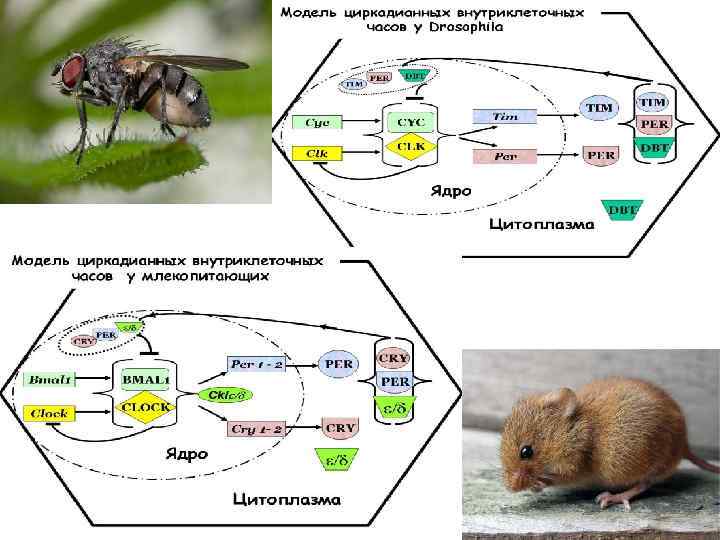
Генетическая модель циркадианных часов у Drosophila melanogaster

Положительный блок • Гены Clk и Cyc являются транскрипционными факторами. • Их белки CLK и CYC образуют в ядре транскрипционный комплекс CYC/CLK который активируют транскрипцию генов Per и Tim. • Уровни матричной РНК генов Per и Tim подниматься и транслируются затем в цитоплазме в протеины.

Отрицательный блок • В цитоплазме белки PER и TIM накапливаются и формируют вместе с белком DBT гетеродимеры (DBT/TIM/PER). Гетеродимеризация служит двум функциям: стабилизации PER, который в отсутствии TIM является нестабильным, и способствует входу комплекса в ядро. • После достижения гетеродимерами в цитоплазме пороговой концентрации, они направляются в ядро. В ядре DBT/TIM/PER ингибирует транскрипционную активность белкового комплекса CYC /CLK, отрицательно воздействуя на транскрипцию генов этих белков и своих собственных. • В ядре оба белка PER/TIM фосфорилируются и постепенно деградируют, в результате блокировка транскрипционного комплекса CYC/CLK снимается. Восстанавливаются функции генов Сyc и Сlk , и вновь запускается синтез м. РНК генов Per и Tim.

Модель циркадианных внутриклеточных часов у Drosophila TIM Cyc Clk PER DBT Tim CYC CLK Per TIM PER Ядро Стрелки показывают активацию или направление процесса, дуга, ограниченная поперечной черточкой – ингибирование. TIM Цитоплазма DBT

Генетическая модель циркадианных часов у млекопитающих

Положительный блок • Два гена Clock и Bmal 1 кодируют транскрипционные факторы CLOCK и BMAL 1. • Эта пара циркадианных белков-активаторов формирует транскрипционный гетеродимерный комплекс CLOCK/BMAL 1. • Комплекс CLOCK/BMAL 1 запускает транскрипцию часовых генов Реr 1/Реr 2 и Cryl/Cry 2, кодирующих соответствующие белки PER и CRY.

• Отрицательный блок • Когда уровень PER- протеинов в цитоплазме достигает определенного уровня, они объединяются с CRY- протеинами и казеинкиназами CK 1 / , и транслоцируются в ядро. • В ядре белки выступает в роли репрессора активности транскрипционного комплекса CLOCK/BMAL 1, в результате блокируется транскрипции генов Per и Cry. • В дальнейшем белки PER и CRY постепенно распадаются под воздействием казеинкиназ и молекулы BMAL 1 и CLOCK высвобождаются, чтобы начать новый суточный цикл в клетке — включить часовые гены Per и Cry.

Модель циркадианных внутриклеточных часов у млекопитающих CRY PER Per 1 - 2 Bmal 1 Clock PER BMAL 1 CLOCK PER Ckl Cry 1 - 2 Ядро CRY Цитоплазма

Время суток Процессы в клетке Дрозофила Мышь Рассвет В ядре запускается транскрипция часовых генов Per, Tim Per 1 - 3, Cry 1 - 2 День Накопление м. РНК часовых генов с последующим синтезом белков в цитоплазме и их накоплением до порогового уровня. PER, TIM PER, CRY Вечер Белки объединяются друг с другом и казеинкиназами, фосфорилируются и возвращаются в ядро. Ночь В ядре они выступают в роли репрессора комплексов транскрипционных факторов. В течение ночи белки CYCLE/CLOCK BMAL 1/CLOCK постепенно распадаются, транскрипционные комплексы освобождаются от блокировки. Рассвет Запускается транскрипция часовых генов PER, TIM, DBT PER, CRY, K Per, Tim Per 1 - 3, Cry 1 - 2

• Гистограмма ритмичности генов из клеток печени • Длина периода транскрипции около трёх с половиной тысяч генов печени (это порядка 4% от всех носителей наследственной информации) примерно сутки, ещё 260 генов – 12 часов, у 63 генов – 8 ч.

• Изменение ритма питания сдвинуло фазу активности 12 часовых генов гепатоцитов печени мышей. • Показан транскрипционный профиль только четырех из восьми исследованных генов.

Температурная независимость молекулярных часов 1. Аминокислотные цепи PER и TIM- белков образуют петли - складываются при взаимодействии аминокислот друг с другом. 2. Кроме того, отдельные молекулы белков PER и TIM образуют димеры, соединяясь друг с другом. 3. Процессы внутримолекулярного и межмолекулярного взаимодействия в точности противоположным образом зависят от температуры. 4. = Повышение температуры приводит к уменьшению активности одного процесса и увеличению активности другого. 5. Таким образом, итоговая «активная поверхность» белка остается постоянной и часы идут независимо от температуры.


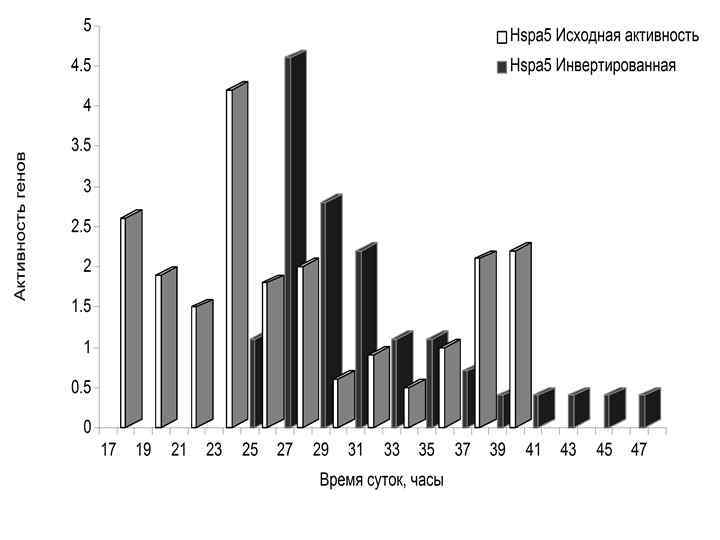
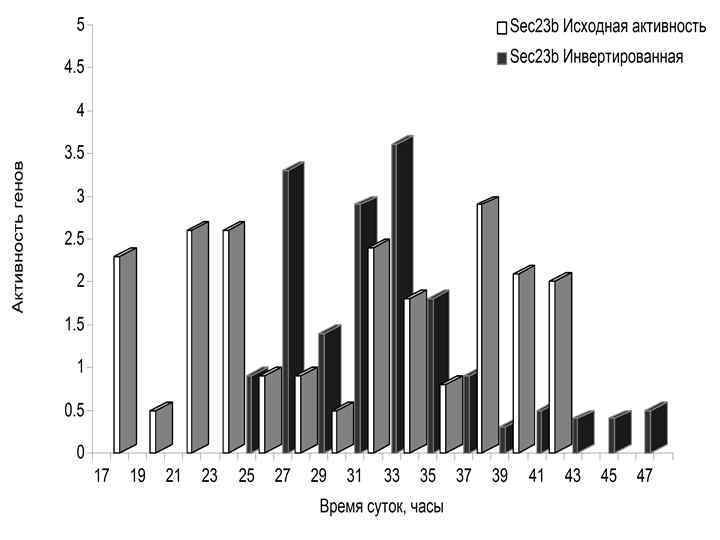
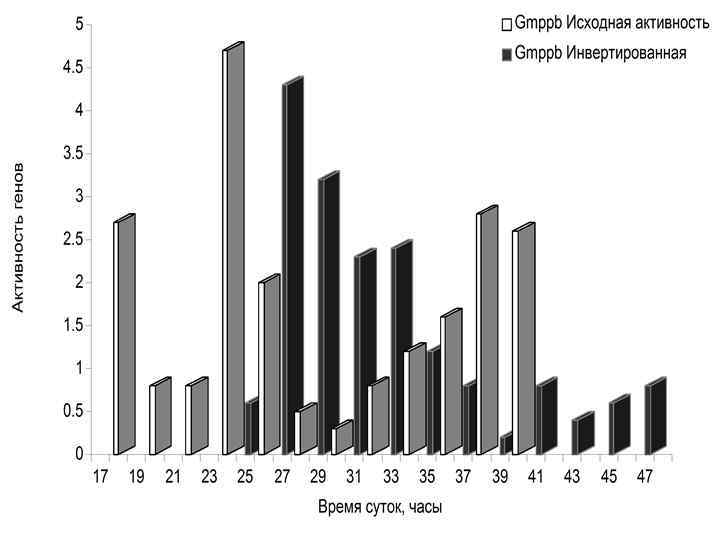
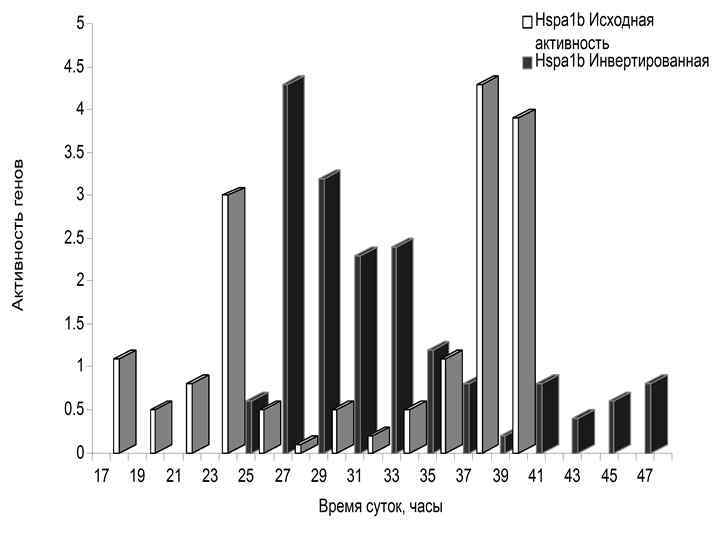

МУТАЦИИ В ЧАСОВЫХ ГЕНАХ (Комаров Ф. И. , Рапопорт С. И. , Малиновская Н. К. , Анисимов В. Н. Мелатонин в норме и патологии. М. , 2004. ) Гены Основные эффекты Per 2 Уменьшение продолжительности жизни, преждевременные нарушения репродуктивной функции, увеличение частоты опухолей Clock Ожирение, метаболический синдром, преждевременные нарушения репродуктивной функции Bmal 1 Уменьшение продолжительности жизни, увеличение перекисного окисления липидов, катаракта, уменьшение мышечной массы Cry 1 -/Cry 2 - Не влияет на продолжительность жизни и развитие опухолей

БИОЛОГИЧЕСКИЕ ЧАСЫ
Биолог. часы_2012.ppt