
Биологическая эволюция Биологическая эволюция — нарушение равновесного






















 13 мая 1707 (Råshult) -")



 Первый закон:")
 Изменение среды Изменение потребностей Выработка")



 Основные труды •")

 открытие гигантских ископаемых")

 1858 г. «О тенденции разновидностей к")
 Thomas Henry Huxley (1825 -1895) С гордостью называет себя бульдогом или")

 В 1863 выступил в защиту")
 – Теория зародышевой плазмы: 1. Эксперименты. 2. Полное отрицание")






: «О некоторых моментах эволюционного процесса с точки зрения современной генетики»")
: «О некоторых моментах эволюционного процесса с точки зрения современной генетики»")


 синтетической теории эволюции (СТЭ) 1. Материалом для эволюции служат,")
 синтетической теории эволюции (СТЭ) 4. Эволюция (в основном) носит дивергентный характер,")








, A (II),")






 Эволюция путем")
 2. Популяционно-экологическое")
 2. Популяционно-экологическое")



Исследования естественного отбора ♂ ♀")






.")








 –")




















 b")



 на мучных")











 называют ту часть популяции, которая эквивалентна")
















 -")




")






: расселение с помощью ветра возможно и для мелких амфибий.")




 Каждый вид должен эволюционировать как можно быстрее для")




 Груз, вносимый оказавшимися не на месте особями (misplaced-individual")












 Manduca sexta, M. quinquemaculata vs Nicotiana")
 Вредитель огурцов, питание стимулируют содержащиеся в тканях")

 vs Dioclea megacarpa (Fabaceae)")











: Гипотеза о расчленяющем характере окраски бабочки Anartia fatima")


 Каждый вид должен эволюционировать как можно быстрее для")

 Основную роль в регуляции")








")
 Номиналистическая концепция вида")



















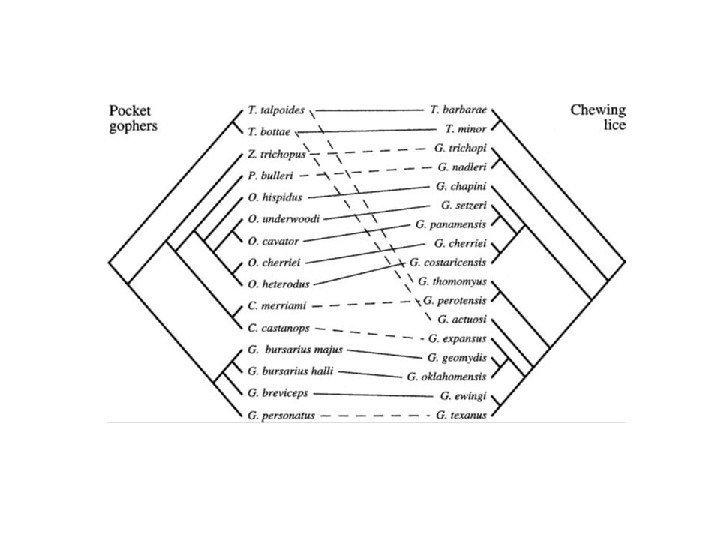
: миграция – изоляция – образование нового вида Ernst Mayr")


 = видообразование путем генетической революции (Mayr, 1954, 1963) =")






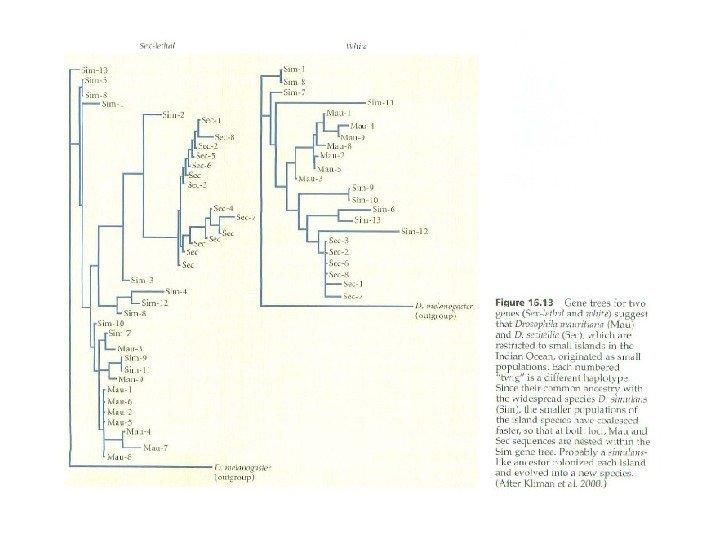



















 –")
 Свои выводы Уайт сделал на основе")
: гомозиготные по разным транслокациям")



Хромосомное видообразование")


")




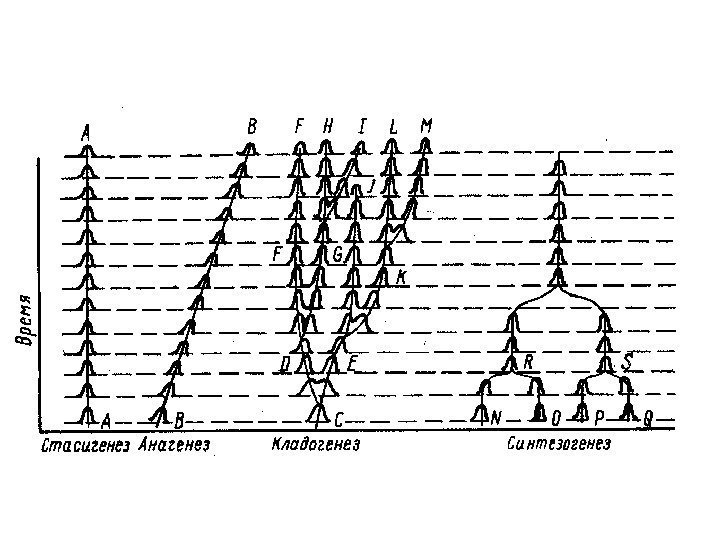









: критерий")








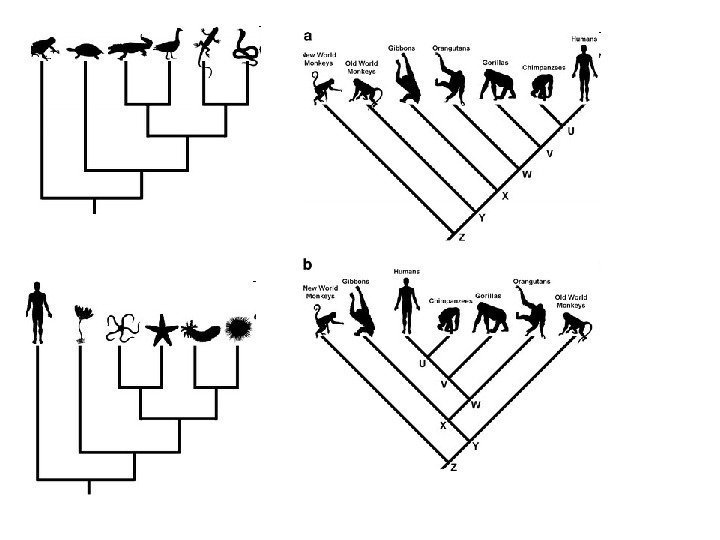




 x=n")





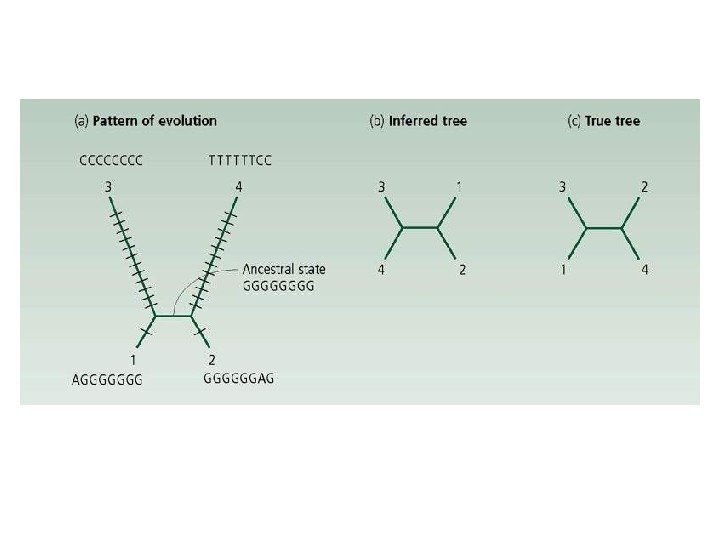









 привел редукцию числа рядов фасеток с")




")

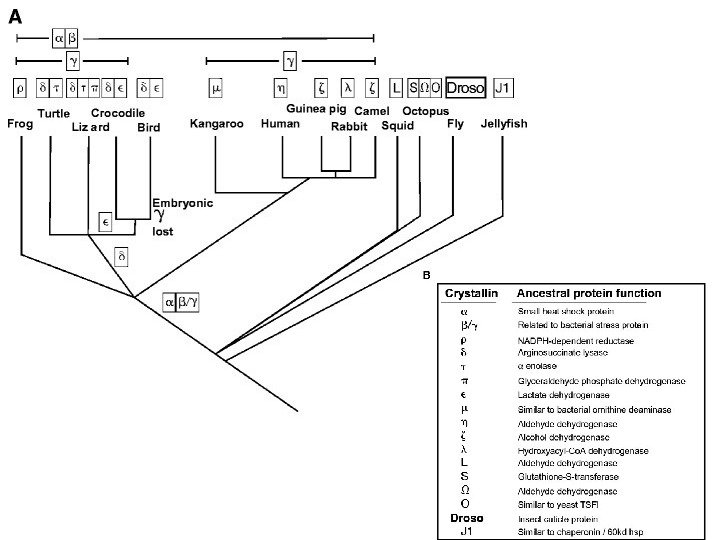
 ген дрозофилы")






 Изменение сроков Название по системе де")


 Маскаренская лысуха (Fulica newtoni)")








evolution2016.pptx
- Размер: 58.2 Мб
- Автор: Елизавета Бойцова
- Количество слайдов: 395
Описание презентации Биологическая эволюция Биологическая эволюция — нарушение равновесного по слайдам
Биологическая эволюция
Биологическая эволюция — нарушение равновесного состояния популяции под действием мутаций и естественного отбора; — приспособление биологических систем к условиям окружающей среды; — историческое развитие группы организмов или развитие жизни на Земле от простейших до высших организмов; — последовательное изменение признака в ряду поколений; Эволюция – серия последовательных изменений в ряду поколений и с исторически значимым результатом.
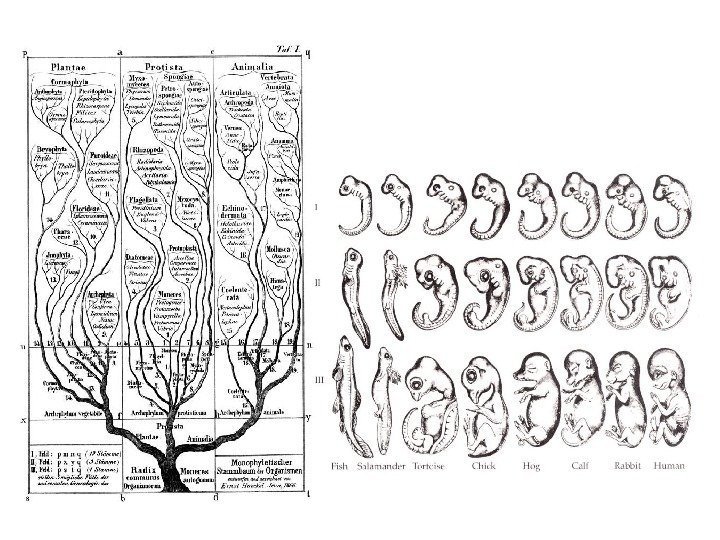
Группы данных, фигурирующие в учебниках как доказательства существования эволюции : Палеонтологические данные Таксономическая структура взаимосвязей между видами Гомология Рудиментарные органы Географическое распространение Прямые наблюдения Экстраполяция наблюдений за эволюционными преобразованиями на популяционном уровне на таксоны более крупного ранга
Палеонтологические данные
Палеонтологические данные
Палеонтологические данные
Таксономическая структура взаимосвязей между видами
Гомологическое сходство
Рудиментарные органы
Географическое распространение
Прямые наблюдения и эксперименты
Прямые наблюдения и эксперименты
Кольцевые ареалы Прямые наблюдения и эксперименты
Экстраполяция наблюдений за эволюционными преобразованиями на популяционном уровне на таксоны более крупного ранга
Десять законов палеонтологии F. I. Pictet, Traité de Paléontologie, 1853 • Различия между вымершими и современными животными тем больше, чем древнее вымершие фауны (третий закон) • Порядок появления типов животных часто напоминает фазы эмбрионального развития (седьмой закон) • С момента, когда какой-либо зоологический тип впервые появился, вплоть до момента, когда он полностью исчезает, нет никакого перерыва в его существовании (восьмой закон) • Вымершие животные построены по тому же плану, что и современные, а их жизнь должна была проявляться в идентичных физиологических актах (десятый закон)
Теория Дарвина – логическое объяснение приспособленности, а также других групп наблюдений, которые теперь приводятся в учебниках как доказательства теории эволюции. Развитие теории эволюции неизбежно, так как сама теория дает толчок развитию и дальнейшему накоплению знаний, которые, в конечном счете, ведут к очередному пересмотру теории. Эволюционная теория — не идеологически выверенный монолит , а богатая альтернативными подходами область знаний, причем различные подходы нередко входят в противоречие другу, предоставляя самокритичную и правдоподобную интерпретацию живого мира. В связи с этим укрепившееся словосочетание «теория эволюции» не слишком удачно, так как по эволюционные представления по сути – не просто теория, а взгляд на мир и одновременно способ объяснения, который используется в абсолютно любой отрасли биологии.
http: //www. icr. org Основан Генри Моррисом в 1972 году. Научный креационизм
1910 – находка отпечатков следов теропод, пересекавшихся с человеческими следами (Paluxy River nr Glen Rose, Texas); 1961 – начало паломничества креационистов в район Пэлюкси. Джон Моррис публикует книгу о загадочных следах; 1968 -1972 – серия ежегодных экспедиций под руководством Тейлора. Снят фильм «Отпечатки ног в камне» ; 1979 – таинственными следами заинтересовался студент биолог Глен Кьюбен
1982 год — бывший баптистский священник Карл Баух начал раскопки в районе реки Пэлюкси. Обнаружил многочисленные новые человеческие следы и другие доказательства сосуществования человека и динозавров, например, человеческие палец и зуб или «след Бердика» и «лондонский молоток». http: //creationevidenc e. org/ The London artifact The Burdick Track Fossilized human finger
http: //www. creationmuseum. org/
Несколько основных подходов изучения эволюции • Популяционно-генетический • Эволюционно-экологический • Молекулярный • Таксономический • Палеонтологический Каждый подход представляет собой особый взгляд на эволюцию, со своими сторонниками, школами и традициями. Нет никаких оснований считать один из подходов более правомочным, нежели другой. Перед эволюционными биологами стоит две основных задачи: 1. В ыявление механизмов эволюционных преобразований 2. Р еконструкция истории эволюционных преобразований
Карл Линней (Carl Linnaeus, позднее Carl von Linné) 13 мая 1707 (Råshult) — 10 января 1778 (Uppsala) Основные биологические труды : • Конспект флоры Лапландии (1733); • Система природы (1735). Последнее, тринадцатое издание вышло посмертно. Десятое издание (1758) впоследствии стало отправной точкой для систематиков; • Флора Швеции (1745); • Фауна Швеции (1746) • Философия ботаники (1751)
Жорж-Луи Леклерк де Бюффон George Louis Leclerc Comte de BUFFON 1707 -1788 В течение почти 50 лет был интендантом, Королевского или Ботанического сада в Париже. Бюффон поднял его на небывалую высоту и превратил в ведущий научный центр Европы. Все это время интенсивно работал над книгой «Естественная история, общая и частная, с описанием Королевского кабинета» . При жизни Бюффона вышло 35 томов, 36 -й том опубликован посмертно. В них вошли человек, четвероногие (млекопитающие), птицы, минералы и история земли.
Жан Батист Пьер Антуан де Ламарк Jean Baptiste Pierre Antoine de Monet de Lamarck (1744 — 1829) Основные биологические труды: Флора Франции, 3 тома (1778) Система беспозвоночных (1801) Мемуары об ископаемых из окрестностей Парижа, 8 томов (1802 -1806) Философия зоологии (1809) Естественная история беспозвоночных животных, 7 томов (1815 -1822)
Теория эволюции Ж. -Б. Ламарка Две формы эволюции: 1. Градация – постепенное повышение организации всех живых существ, основанное на “внутреннем стремлении к совершенствованию”. 2. Приспособительная эволюция Jeanne-Baptiste de Lamarck (1744 -1829) Два механизма приспособительной эволюции: 1. Прямое влияние условий существования 2. Изменение потребностей и привычек
Теория эволюции Ж. -Б. Ламарка Законы Ламарка Jeanne-Baptiste de Lamarck (1744 -1829) Первый закон: «Принцип упражнения-неупражнения органов» Второй закон: «Принцип наследования приобретенных (благоприобретенных) признаков»
Схема эволюции по Ламарку Jeanne-Baptiste de Lamarck (1744 -1829) Изменение среды Изменение потребностей Выработка новых привычек Упражнение органов в соответствии с новыми условиями и привычками Усиление упражняемых и ослабление неупражняемых органов Передача потомкам измененных признаков
Теория эволюции Ж. -Б. Ламарка Теория Ламарка не может объяснить эволюцию: 1. Средств пассивной защиты (ядовитость, мимикрия, покровительственная окраска). 2. Органов и частей, состоящих из мертвых тканей (раковина и т. п. ). 3. Органов и привычек, используемых один раз в жизни (яйцевой зуб цыпленка). 4. Бесплодных рабочих у общественных насекомых. 5. Взаимных адаптаций видов при коэволюции (опыление, мутуализм и т. п. ). 6. Появление принципиально новых признаков (хорда, глаз и т. п. ). 7. Усиление и редукцию многих органов: — Усиление мышц; — Редукция крыльев у насекомых. Jeanne-Baptiste de Lamarck (1744 -1829)
Современные формы ортогенеза и ламаркизма Лима-де-Фариа А. Эволюция без отбора. Автоэволюция формы и функции. Мир, М. , 1991, 455 с. Чайковский Ю. В. Эволюция. Книга для изучающих и преподающих эволюцию. Центр системных исследований, М. , 2003, 472 с.
Жорж Леопольд Кретьен Фредерик Дагобер Кювье Georges Léopold Chrétien Frédéric Dagobert Cuvier 1769 -1832 Основные труды: • Элементарный курс естественной истории животных (1798) • Лекции по сравнительной анатомии (1799 -1800) • Исследование ископаемых костей четвероногих, четыре тома (1812) • Царство животных, 4 тома (1817)
Этьен Жоффруа Сент-Илер Étienne Geoffroy de Saint-Hilaire (1772 -1844) Основные труды • Философия анатомии: 1818 – 1 том, 1822 – 2 том • Принципы философии зоологии, дискутированные в марте 1830 года в лоне Королевской Академии Наук (1830) Отстаивал единство плана строения всех животных. Занимаясь выяснением причин многообразия животного мира пришел к выводу, что постепенно меняющиеся во времени условия жизни вызывают преемственные изменения поколений животных, т. е. изменения видов.
Чарльз Дарвин Charles Robert Darwin 12 февраля 1809 — 19 апреля 1882 Натуралистическое образование получил самостоятельно. В 1825 -1827 годах изучал медицину в Эдинбургском университете. C 1827 по 1831 – теологию в Кембридже. С 1831 по 1836 г. совершает кругосветное путешествие на корабле «Бигл».
В ходе путешествия на Дарвина произвели самое сильное впечатление: « 1) открытие гигантских ископаемых животных, которые были покрыты панцирем, сходным с панцирем современных броненосцев; 2) то обстоятельство, что по мере продвижения по материку Южной Америки близкородственные виды животных замещают одни других; 3) тот факт, что близкородственные виды различных островов Галапагосского архипелага незначительно отличаются друг от друга» . Geochelone elephantopus Bradipus variegatus
В 1839 году публикует описание путешествия на Бигле, а в течение последующих семи лет издает пятитомные зоологические и биологические результаты экспедиции. В 1842 году Дарвин делает черновой набросок в 35 страниц. Через два года расширяет его до 230 страниц В 1856 он начал готовить свою работу к публикации и к 1858 году им написано более 2000 страниц, что составляет лишь половину от запланированного.
Alfred Russell Wallace (1823 — 1913) 1858 г. «О тенденции разновидностей к неограниченному отклонению от первоначального типа» . Выдержки из книги Дарвина публикуются в «Трудах Линнеевского Королевского общества» одновременно со статьей Уоллеса. 24 ноября 1859 – выход в свет книги «Происхождение видов путем естественного отбора или сохранение благоприятных рас в борьбе за жизнь». Альфред Уоллес
Томас Гексли (Хаксли) Thomas Henry Huxley (1825 -1895) С гордостью называет себя бульдогом или цепным псом Дарвина. Своей неутомимой деятельностью во многом сформировал общественное мнение о том, что такое дарвинизм. В 1863 году публикует книгу «Зоологические свидетельства о положении человека в природе»
Среди видных естествоиспытателей – современников Дарвина, его теорию КАТЕГОРИЧЕСКИ НЕ ПРИНЯЛИ: учитель Дарвина геолог Адам Седжвик выдающийся палеонтолог и морфолог Ричард Оуэн Зоолог, палеонтолог и геолог Луи Агассис. ПОДДЕРЖАЛИ : американский ботаник Эйс Грей английский геолог Чарльз Лайель Альфред Уоллес Томас Гексли
Эрнст Геккель Ernst Heinrich Haeckel (1834 — 1919) В 1863 выступил в защиту Дарвина на съезде германских естествоиспытателей. 1866 «Общая морфология. Общий очерк учения об органических формах, механически обоснованного на основе реформированной Чарльзом Дарвином эволюционной теории» . Те же взгляды в популярной форме были изложены в «Естественной истории мироздания» .
Возникновение неодарвинизма: August Weismann (1883) – Теория зародышевой плазмы: 1. Эксперименты. 2. Полное отрицание наследования приобретенных признаков (а также и роли модификационной изменчивости). 3. Тезис о всемогуществе естественного отбора. 4. Теория зародышевого отбора.
Критики дарвинизма сосредоточили внимание на следующих вопросах: 1. Может ли естественный отбор, отсеивающий нежизнеспособных, играть творческую роль, обеспечивать поступательность эволюционного процесса? 2. Каким образом благоприятные признаки выживших особей могут сохраняться в потомстве, не растворяясь при скрещивании носителей этих признаков с особями, несущими иные признаки. Именно в эти годы ( а не после выхода «Философии зоологии» ) возник ламаркизм как антитеза дарвинизму.
В 1900 году законы Менделя были заново открыты в трех разных странах. Цитологическое обоснование менделизма было дано в 1902 году Уолтером Саттоном на основе изучения хромосом кузнечиков. Hugo de Vries (1848 -1935) Carl Erich Correns (1864 -1933) Erich von Tschermak-Seysenegg (1871 -1962)
Вскоре после переоткрытия законов Менделя была сформулирована Мутационная теория де Фриза. Основной объект исследований этого автора — Oenothera lamarkiana.
• Причины неприятия дарвинизма ранними генетиками: – Гены считались практически или совершенно неизменными – Выделяли две формы изменчивости: • Флуктуации – ненаследственные, возникают при влиянии среды • Мутации – очень редкие и крупные изменения – наследуются – Вызвать мутации очень трудно – постоянство видов – Если мутация происходит – это сразу образование нового вида – Мелкие мутации были неизвестны – Отбор – это лишь сито, отделяющее нежизнеспособные формы (виды)
• Формы генетического антидарвинизма в начале XX века: – Мутационизм (Г. Де Фриз, В. Иогансен, У Бэтсон, Т. Морган и др. ) – Преадаптация (Л. Кено, 1901 -1914; Ч. Девенпорт, 1903) – Гибридогенез (И. Лотси, 1916) • Другие формы антидарвинизма в начале XX века: – А. Бергсон – « Творческая эволюция » (1907): эволюцию направляет “жизненный порыв” – Г. Осборн – известный палеонтолог – концепция “аристогенеза”; “аристогены” – гены прогресса – Л. С. Берг – « Номогенез, или эволюция на основе закономерностей » (1922): • число мутаций ограничено и отбору здесь не из чего отбирать • эволюцию движут строгие закономерности • изначальная целесообразность – непостигаемое и «далее неразделимое свойство живого» • «Организмы развились из многих тысяч первичных форм. . . дальнейшее развитие шло преимущественно конвергентно
Четвериков ( 1926): «О некоторых моментах эволюционного процесса с точки зрения современной генетики» мутационный процесс происходит в природных популяциях; большинство вновь возникших мутаций понижает жизнеспособность, хотя изредка возникают мутации, повышающие таковую; в условиях свободного скрещивания вид достаточно устойчив и сохраняет исходное соотношение частот аллелей, а каждая рецессивная мутация «впитывается видом в гетерозиготном состоянии» и при отсутствии отбора может сохраняться неограниченно долго; гетерозиготность по разным аллелям разных генов «пропитывает вид во всех направлениях» и в результате случайных комбинаций постепенно «заражает» большинство индивидов вида; генетическая изменчивость проявляется наиболее сильно, когда многочисленный вид распадается на ряд небольших, изолированных колоний; наиболее обычна пространственная изоляция, хотя может существовать и временнáя экологическая изоляция;
Четвериков ( 1926): «О некоторых моментах эволюционного процесса с точки зрения современной генетики» . изоляция, наряду с наследственной изменчивостью, является основным фактором внутривидовой (а следовательно, и межвидовой) дифференциации; естественный отбор является антагонистом свободного скрещивания; приспособительная эволюция без изоляции ведет к полной трансформации вида, но не может привести к его полному распадению на два вида; отбор, как и свободное скрещивание, ведет к накоплению рецессивных аллелей в гетерозиготном состоянии; свободное скрещивание ведет к повышению полиморфизма вида, а отбор ведет к мономорфизму; отбор отбирает не только отдельный ген, контролирующий отдельный признак, но и, благодаря множественности (плейотропности) действия генов, влияет на все генное окружение.
«Отцы» синтетической теории эволюции
«Отцы» синтетической теории эволюции
Основные идеи (постулаты) синтетической теории эволюции (СТЭ) 1. Материалом для эволюции служат, как правило, очень мелкие, но дискретные изменения наследственности — мутации. Мутационная изменчивость носит случайный характер. 2. Основным или даже единственным движущим фактором эволюции является естественный отбор, основанный на отборе (селекции) случайных и мелких мутаций. 3. Наименьшая эволюционирующая единица эволюции — популяция, а не особь, как это допускалось, исходя из представлений о возможности «наследования благоприобретенных признаков».
Основные идеи (постулаты) синтетической теории эволюции (СТЭ) 4. Эволюция (в основном) носит дивергентный характер, т. е. один таксон может стать предком нескольких дочерних таксонов, но каждый вид имеет единственный предковый тип, а в конечном итоге и единственную предковую популяцию. 5. Эволюция носит постепенный (градуалистический) и длительный характер. Видообразование, признаваемое этапом эволюционного процесса, мыслится в виде поэтапной смены одной временной популяции чередой последующих временных популяций. 6. Вид состоит из множества соподчиненных, морфологически, физиологически и генетически отличных, но репродуктивно не изолированных единиц — подвидов, популяций.
7. Обмен аллелями, «поток генов» возможен лишь внутри вида. Если мутация имеет положительную селективную ценность на всей территории ареала вида, то она может распространиться за счет «потока генов» по всем его популяциям и подвидам. Отсюда следует краткое и достаточно емкое определение биологического вида: вид есть генетически целостная и замкнутая система. 8. Поскольку критерием так называемого биологического вида является его репродуктивная обособленность, эти критерии не применимы к формам без полового процесса, агамным, амфимиктичным, партеногенетическим формам (следствие из п. 7). Основные идеи (постулаты) синтетической теории эволюции (СТЭ)
9. Макроэволюция, или эволюция на уровне выше вида, идет лишь путем микроэволюции. За пределами вида эволюция фактически прекращается. 10. Любой реальный, а не сборный таксой имеет монофилетическое происхождение (следствие из п. 4). 11. Исходя из всех упомянутых постулатов ясно, что эволюция непредсказуема, имеет не направленный к некоей конечной цели, т. е. нефиналистический, характер. Основные идеи (постулаты) синтетической теории эволюции (СТЭ)
Популяция – изолированная совокупность особей данного вида, характеризующихся общностью происхождения, местообитания и образующих целостную генетическую систему. Популяции очень разнообразны по структуре, которая складывается из четырех главных компонентов: пространственной конфигурации; величины; системы размножения; скорости расселения.
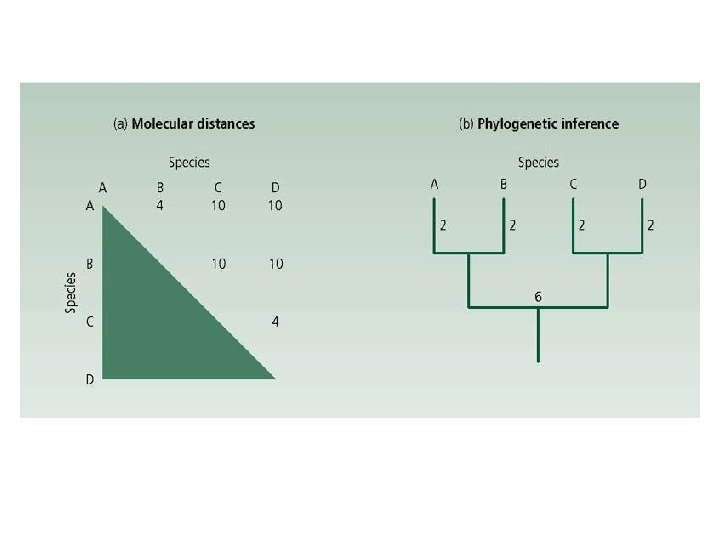
Популяционные генетики, рассматривают микроэволюцию прежде всего как изменения частот гомологичных аллелей, участков хромосом или целых хромосом в локальной популяции. Теорема или закон постоянства частот аллелей (Hardy — Weinberg, 1908): p = P + ½ H; q = Q + ½ H 222 q 2 pq p q)(p pqpppqp Η 1/2 P p 2 q q) q(p pq q 1/2 H Qq
Условия: популяция имеет бесконечно большой размер, так что ошибки выборки не оказывают влияния на частоты в последующих поколениях; популяция изолирована, иммиграция и мутационное давление отсутствуют, то есть приток генетического материала извне отсутствует; все особи вносят равное число функционирующих гамет, то есть особи разных генотипов размножаются одинаково успешно; скрещивание случайное (панмиктическое).
Рассмотренным способом можно определять частоты генотипических классов как в искусственных, так и в природных популяциях Популяции лисиц Западной Сибири исследовались по окраске: рыжие, черные с проседью и полные меланисты (Ромашов, Ильина, 1942). Тихоокеанские тонкоклювые кайры на острове Тюленьем различались по фоновой окраске скорлупы (белые, голубые, зеленые) (Богданов, 1977). Генотипически контролируемые различия в окраске (1 — 3) яиц у тонкоклювой кайры Uria aalge inomata (по Л. В. Богданову, 1977): 1 — зеленая» , 2 — голубая, 3 — белая;
В случае полного доминирования использование формулы Харди-Вайнберга также возможно, но с оговорками, так как частоты аллелей не могут быть выведены непосредственно из частот генотипических классов.
Генотипические частоты в равновесной популяции естественно связаны с соотношением частот аллелей. Графически эта зависимость изображается т. н. параболой Финетти. При p = q = 0. 5 частота гетерозигот максимальна и равна 0. 5 При q > 2/3 → aa>Aa; p>2/3 → AA>Aa.
Три аллеля, шесть генотипических классов: Антигенные группы крови человека — 0 (I), A (II), B(III) и AB (IV) детерминируются тремя аллелями гена I: I A , I B , I 0. Аллели А и В кодоминируют. 1 r 2 qr 2 pr q 2 pq p r) q (p
AABB AABb AAbb Aa. BB Aa. Bb Aabb aa. BB aa. Bb aabb. Два несцепленных двухаллельных аутосомных гена 9 генотипических классов: 4 типа гамет: AB Ab a. B ab При конордантном (согласованном) соотношении частот гамет выдерживается равенство: Ab × a. B = AB × ab
Генотипический состав популяций изменяется в силу: 1. мутационного процесса; 2. колебаний численности особей в популяциях ( «Волны жизни» ); 3. изоляции – возникновения барьеров внутри и между популяциями; 4. естественного отбора. Элементарным эволюционным явлением следует считать длительное, векторизованное преобразование генотипического состава популяции.
Естественный отбор Яйца и только что вылупившиеся предличинки, личинка и взрослая особь атлантической трески ( Gadus callarias ) В теории популяционной генетики естественный отбор рассматривается как важнейший фактор эволюции, вызывающий адаптивные изменения в генетической структуре популяций. Эти изменения – результат различий между вкладами разных генотипов в генофонд следующего поколения, которые возникают за счет их дифференциального воспроизводства и выживаемости. Для отбора по данному признаку необходимы: 1. борьба за существование; 2. изменчивость в популяции по признаку; 3. связь между признаком и приспособленностью; 4. связь между проявлением признака у родителей и потомства, т. е. наследование данного признака.
Основные параметры математического описания отбора. W AA – абсолютная приспособленность. Складывается из дифференциальной выживаемости и дифференциальной плодовитости. w AA – относительная приспособленность ; w = 1 у генотипа с максимальной абсолютной приспособленностью Относительные приспособленности остальных генотипов взвешивают относительно максимальной. S AA – коэффициент отбора. w – мера эволюционного успеха носителей генотипа и, соответственно, она является обратной величиной по отношению к отбору: s AA = 1–w AA s Aa = 1–w Aa s aa = 1–w aa Средняя приспособленность популяции является функцией аллельных частот : В состоянии равновесия: 0 dq wd dp wd 22 2 1211 2 2 wqpqwwpw
В 1930 году Р. Фишер предложил фундаментальную теорему естественного отбора: «Скорость возрастания средней приспособленности популяции равна генетической (аддитивной) дисперсии приспособленности этой популяции» . Естественный отбор – единственный популяционный фактор, который всегда увеличивает среднюю приспособленность популяции. w w dt wd
Согласно концепции С. Райта, окружающую среду можно рассматривать как адаптивный ландшафт , состоящий из холмов и долин. Рельеф местности символизирует распределение адаптивных полей, где вершины холмов – адаптивные пики, а низменные участки между ними – адаптивные долины.
Современные подходы к определению отбора 1. Логическое определение (J. A. Endler, 1986) Эволюция путем естественного отбора происходит при условии, что в популяции существует: (а) изменчивость особей по некоторому признаку; (б) определенная связь между этим признаком и каким-либо компонентом приспособленности; (в) определенная связь по этому признаку между родителями и потомками (наследование признака). • . Условия (а), (б) и (в) – необходимы и достаточны для того, чтобы эволюция путем отбора происходила. • . Однако, это не означает, что адаптивная эволюция обязательно будет происходить.
Современные подходы к определению отбора 1. Логическое определение (J. A. Endler, 1986) 2. Популяционно-экологическое определение «Естественный отбор – это наличие корреляции между индивидуальной изменчивостью признаков и репродуктивным успехом особей» Певчий воробей Melospisa melodia. Галапагосский вьюрок Geospisa fortis
Современные подходы к определению отбора 1. Логическое определение (J. A. Endler, 1986) 2. Популяционно-экологическое определение 3. Популяционно-генетическое определение «Естественный отбор – это дифференциальное и неслучайное выживание и размножение особей с разными генотипами в популяции» (Lerner, 1958)
Естественный отбор: — вероятностный процесс — решает сиюминутные проблемы — не имеет цели и не может предвидеть будущее — не может создать организм заново, начать «с нуля» , а лишь модифицирует то, что уже есть — далеко не всегда находит «идеальное решение»
http: //www. tylervigen. com/spurious-correlations
Полиморфизм в окраске у божьей коровки Adalia bipunctata
Большой кактусовый вьюрок Geospiza conirostris (Isla Genovesa)Исследования естественного отбора ♂ ♀
Лядвенец рогатый Lotus corniculatus Исследования естественного отбора Пустельга Falco tinnunculus
Орхидея Anagraecum sesquepedale отличается удивительно длинным шпорцем длинной 28 -32 см. Дарвин предсказал, что эта орхидея опыляется гигантским бражником с 30 -сантиметровым хоботком. Действительно, 40 лет спустя был описан бражник Xanthopan morgani predicta. Nelson, 1988: укорачивал нектарники орхидей на островах Балтики и отмечал количество снятых насекомыми поллиний и оплодотворенных рылец. Орхидеи с укороченными венчиками обладали сниженной приспособленностью.
Staphylococcus aureus
Staphylococcus aureus Пенициллин G – метицилин – фторхинолон – мупироцин – …
Половой отбор , равно как и любой другой, основан на селективной неравноценности особей, но происходит между особями одного пола (чаще всего самцами) у раздельнополых организмов. Вторичные половые признаки можно разделить на: Признаки, относящиеся к размерам, силе (более крупные размеры самцов морских львов, клыки у самцов тюленей). Появление такого рода признаков трактуется как следствие конкуренции за полового партнера. Украшения и демонстрационное поведение (яркое оперение). Следствие активного выбора партнера противоположным полом.
Две формы полового отбора: 1. Интрасексуальный.
Две формы полового отбора: 1. Интрасексуальный. 2. Интерсексуальный (эпигамный).
Природа полового отбора. Половой отбор – это естественный отбор, основанный на конкуренции за возможность спаривания и выборе полового партнера, при котором успех в спаривании может вступать в противоречие с выживанием, т. е. между этими параметрами возникают компенсации (trade-offs)
Причины интереса к изучению полового отбора Половой диморфизм. Возможность очень быстрой эволюции. Роль в видообразовании.
Фундаментальные различия репродуктивных стратегий полов, лежащие в основе полового отбора. Классические эксперименты Бейтмана (A. J. Bateman, 1948) Принцип Бейтмана: «Вариабельность репродуктивного успеха самцов гораздо больше, чем у самок»
Принцип Бейтмана
Принцип Бейтмана
Принцип Бейтмана Число потомков у самок и самцов льва
Принцип Бейтмана Число потомков у самок и самцов красного оленя
Принцип Бейтмана Число спариваний у самцов болотного козла и белобородого манакина
Неравноценность родительского вклада самок и самцов как причина полового отбора. Robert Trivers (1972) – Концепция родительского вклада Родительский вклад – любое участие родителя в выращивании потомства, повышающее его шансы на выживание: гаметы кормление защита и др. Если родительский вклад одного пола существенно превышает вклад другого, то особи пола, вносящего больший вклад, становятся для особей противоположного пола ограниченным ресурсом , за который они будут конкурировать.
Неравноценность родительского вклада самок и самцов как причина полового отбора.
Неравноценность родительского вклада самок и самцов как причина полового отбора. Важные выводы: 1. При прочих равных условиях отбор должен приводить к стремлению самцов спариваться с возможно большим числом самок – изначальная полигамность самцов. 2. Поэтому между самцами должна возникать конкуренция за самок. 3. При прочих равных условиях отбор должен усиливать стремление самок “относиться серьезнее” к выбору брачного партнера, быть “застенчивыми”, проявлять осторожность и нерешительность. 4. Самки, в отличие от самцов, могут повысить свою приспособленность путем правильного выбора единственного партнера и усилением заботы о потомстве – изначальная моногамность самок.
Редкие случаи ролевой реверсии полов как подтверждение теории родительского вклада. Плавунчики
Редкие случаи ролевой реверсии полов как подтверждение теории родительского вклада. Выбор самцов самками у иглы-рыбы
Редкие случаи ролевой реверсии полов как подтверждение теории родительского вклада. Самец мормонова сверчка со сперматофором
Редкие случаи ролевой реверсии полов как подтверждение теории родительского вклада. Выбор самок самцами мормонова сверчка
Системы спаривания у животных: Моногамия и полигамия Полигиния, полиандрия и полигинандрия 1. Полигиния – наиболее естественная, изначальная система спаривания. 2. Требуют специального объяснения: моногамность самцов полиандричность самок Влияние системы спаривания на интенсивность полового отбора.
http: //www. tylervigen. com/spurious-correlations
Фенотип Приспособленность Частота до отбора Частота после отбора Стабилизирующий, направленный и дизруптивный отбор
Дизруптивный отбор – Streams & Pimentel, 1961: отбор на количество щетинок у Drosophila melanogaster
Помимо дизруптивного отбора, действующего в неоднородной среде, полиморфизм поддерживают: Уравновешивающий отбор = отбор в силу преимущества гетерозигот Частото-зависимый отбор Hovanitz, 1953; Remington, 1954: Полиморфизм в популяции североамериканской желтушки Colias eurytheme Дизруптивный отбор
Еще одна классификация типов отбора основана на уровне отбора. Она подразумевает выделение отбора на видовом уровне; индивидуального отбора; группового и отбора родичей (кин-отбора); субиндивидуального отбора. Нередко выделяют три типа отбора действующих на видовом уровне : 1. Отбор, направленный на развитие экологической дивергенции 2. Замещение одного вида другим 3. Отбор, приводящий к репродуктивной изоляции.
Эволюция альтруизма и полового размножения Два определения понятия “альтруизм”: Широкое Альтруизм – это действия, осуществляемые во вред себе, но на благо другим. Узкое Альтруизм – это поведение, которое приводит к увеличению выживания и/или числа потомков других особей, не являющихся прямыми потомками альтруиста, и уменьшению вероятности выживания альтруиста и/или числа его потомков. Это так называемый “истинный альтруизм”
Примеры истинного альтруизма Кормление своих и чужих щенков у гиеновых собак
Примеры истинного альтруизма Самопожертвование у медоносной пчелы, солдат термитов и тлей
Примеры истинного альтруизма Потеря способности к размножению у общественных насекомых и голых землекопов
Родственный отбор «Это отбор, оперирующий совокупной приспособленностью особей» Название: “kin selection” (Maynard Smith, 1964) Три перевода: “родственный отбор”, “отбор родичей”, или “кин-отбор” Родственный отбор
Родственный отбор Идея Дж. Холдэна: Альтруизм может возникать, если он направлен на родственников Теория В. Гамильтона William Hamilton (1964) – Эволюция социального поведения на основе родственности (“Kinship Theory”) Впервые осмыслил значение для эволюции наличия у родственных особей генов, идентичных по происхождению Его работы положили начало совершенно новым концептуальным подходам в СТЭ и явились основой социобиологии
Способы вычисления коэффициентов родственности 1. Вероятностный метод Коэффициент родственности – это: Вероятность наличия у обеих особей в данном локусе аллеля идентичного по происхождению Доля идентичных по происхождению генов в генотипе (для родителей и потомков – это точная мера, для всех прочих родичей – вероятностная) n – число ”путей” передачи L – число мейозов или поколений. Родственный отбор. L nr)5. 0(
Способы вычисления коэффициентов родственности 1. Вероятностный метод n – число ”путей” передачи L – число мейозов или поколений Родители – Потомки Сибсы Бабушка – Внуки Тетушка – Племянники. Родственный отбор. L nr)5. 0( 5. 0)5. 0(11 r 5. 0)5. 0(22 r 25. 0)5. 0(12 r 25. 0)5. 0(23 r
Альтруистическое поведение можно объяснить, с помощью теории кин-отбора Основные параметры: c (cost) b (benefit) r (relatedness). Доля родства может быть высчитана с помощью генерационного расстояния (числа разделяющих поколений) и составляет (1/2) L Правило Гамильтона: При rb > с средняя приспособленность альтруистов будет выше таковой «эгоистов» и количество копий генов альтруизма должно увеличиваться.
Исследования альтруизма у кустарниковой голубой сойки Aphelocoma coerulescens В среднем, на каждое из гнезд приходилось по 1. 8 помощника, эту величину округлили до двух. Выживаемость птенцов при наличии помощников возрастает от двух до пяти раз, в среднем с 7 до 35 %. Соответственно, b= 28/2= 14%. Даже если бы все помощники создавали свои гнезда в одиночку, утерянная выгода с=7%. Модифицированное правило Гамильтона: r sib × b > r of × c ½ × 14 > ½ ×
Эусоциальные насекомые Рой Polybia occidentalis
Групповой отбор и междемовый отбор Групповой отбор в широком смысле: может действовать на группы особей очень разной природы – популяции, колонии, семьи, социальные группы и т. д. включает разные механизмы, в том числе и родственный отбор является недостаточно четко определенным понятием Групповой отбор в узком смысле – Междемовый отбор: действует на группы популяционной природы – демы, ценопопуляции, локальные популяции и т. п. является четко определенным понятием
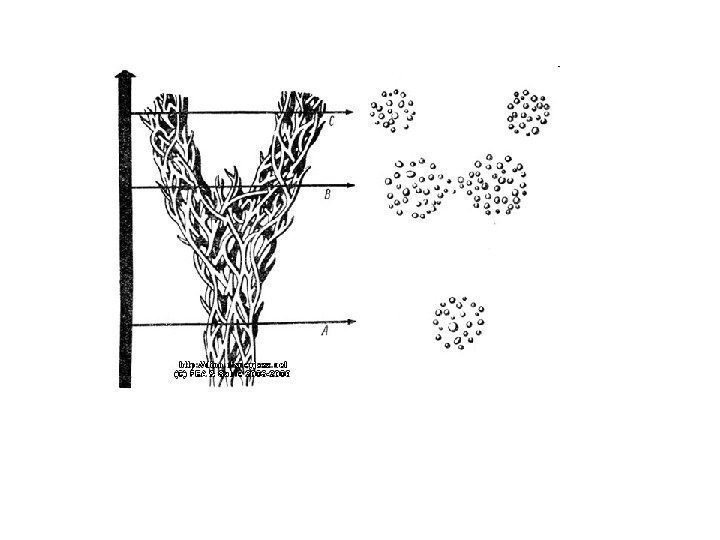
Возможные эффекты группового отбора в некоторых ситуациях: 1. Эксперименты Wade (1977) на мучных хрущаках Tribolium
Междемовый отбор как процесс второго порядка: 1. Плодовитость и смертность особей >> чем групп 2. Генетическая изменчивость особей >> чем групп 3. Корреляция между признаками и репродуктивным успехом особей >> чем у групп Поэтому отбор особей сильнее, чем междемовый отбор Групповой отбор
Биологический альтруизм как генетический эгоизм Следует подчеркнуть: Нет необходимости предполагать осознанность действий альтруиста Причины направленности альтруизма на родичей могут быть различными и необязательно связаны с выбором особи Нельзя говорить, что альтруист “стремится передать свои гены в следующее поколение” Такой результат возникает просто потому, что так действует отбор Родственный отбор
Отбор на уровне генов и хромосом Нарушение сегрегации у эукариот Мейотический драйв Аллели – нарушители сегрегации ( SD – segregation distorter) В гетерозиготе разрушают гомологичную хромосому во время мейоза Механизм требует наличия двух сцепленных локусов; один распознает парную хромосому, второй ее разрушает Результат – преобладание SD гамет – т. е. нарушение законов Менделя Это означает действие отбора на уровне генов в пользу SD аллелей SD аллели найдены у дрозофилы, домовых мышей, комаров, некоторых растений
Нейтральная эволюция
Нейтральная эволюция Схема, иллюстрирующая возможные взаимоотношения генотипической изменчивости и приспособленности.
Термин «нейтральный» может быть применен лишь в сравнительном смысле. Мы называем мутацию нейтральной, если она обладает той же приспособленностью, что и предковая аллель. Поскольку нейтральные аллели по определению не связаны с различиями в приспособленности, их частоты в популяции определяются случайными событиями. Причины отсутствия корреляции между генотипической изменчивостью и приспособленностью различны. Изменение генофонда может не проявляться фенотипически. Изменения в аминокислотной последовательности не всегда влияют на функцию. Канализация развития.
ЧТО ИМЕННО ПОНИМАЕТСЯ ПОД СЛУЧАЙНОСТЬЮ? Генетический дрейф – изменение частот аллелей в популяции в ряду поколений под действием случайных (стохастических) факторов.
Представим множество популяций ограниченного размера численности N и одинаковыми частотами аллелей p и q. Распределение частот аллелей в них будет подчиняться биноминальному закону: (p A + q A ) 2 N Случайное отклонение частот аллелей в каждом отдельном случае равно σ q =q i -q. Дисперсия σ q за одно поколение составляет : Например, при q = p = 0. 5 и N = 50 Фиксация или утрата одной из аллелей при прочих равных равновероятна. Она совершается со скоростью 1/2 N за поколение , из них 1/4 N – доля фиксируемых и 1/4 N – доля теряемых аллелей. N qq q 2 )1(2 05. 0 5 0*2 5. 0*5. 02 q
Экспериментальные исследования дрейфа генов Согласно оценке С. Райта, случайные процессы становятся эффективными при условии 4 Ns<<1.
Atoll of Pingelap The island of color blindness
Коэффициент инбридинга – вероятность того, что при образовании зиготы очередного поколения в одном из ее локусов окажутся аллели, идентичные по происхождению, т. е. появившаяся особь будет аутозиготной по данному локусу гермафродитная популяция обоеполая популяция Эффективная величина популяции неизменна, соотношение полов всегда составляет 1:
ЭФФЕКТИВНАЯ ВЕЛИЧИНА ПОПУЛЯЦИИ Эффективным размером (величиной) называют ту часть популяции, которая эквивалентна доле особей, оставляющих потомство при панмиктическом размножении. Оценки эффективной величины популяции в различных ситуациях: 1. Неравное участие в размножении самок и самцов. Лимитирующим фактором здесь будет доля особей малочисленного пола. 2. Периодические колебания численности популяции. На практике формула применима лишь для небольшого числа поколений. 3. Инбридинг. Коэффициент инбридинга варьирует от 0 до 1. В случае когда F=1, эффективная величина популяции равна половине ее фактической численности. Nf. Nm Nm. Nf Ne 4 Nt. NNTNe 1. . . 2 1 1 111 F Ne
Генетический дрейф Популяционные волны как фактор микроэволюции “ Популяционные волны” – это значительные по амплитуде периодические или апериодические колебания численности популяции «Волны жизни» по С. С. Четверикову (1905)
Генетический дрейф Популяционные волны как фактор микроэволюции “ Популяционные волны” – это значительные по амплитуде периодические или апериодические колебания численности популяции «Волны жизни» по С. С. Четверикову (1905)
Генетический дрейф Популяционные волны как фактор микроэволюции “ Популяционные волны” – это значительные по амплитуде периодические или апериодические колебания численности популяции «Волны жизни» по С. С. Четверикову (1905) Нередко волны жизни связаны не только с изменениями численности, но и пространственным распределением особей
Генетический дрейф Популяционные волны как фактор микроэволюции “ Популяционные волны” – это значительные по амплитуде периодические или апериодические колебания численности популяции «Волны жизни» по С. С. Четверикову (1905) Нередко волны жизни связаны не только с изменениями численности, но и пространственного распределения особей
Генетический дрейф Популяционные волны как фактор микроэволюции “ Популяционные волны” – это значительные по амплитуде периодические или апериодические колебания численности популяции «Волны жизни» по С. С. Четверикову (1905) Нередко волны жизни связаны не только с изменениями численности, но и пространственного распределения особей Они могут быть причиной генетического дрейфа
Генетический дрейф Популяционные волны как фактор микроэволюции Эффект бутылочного горлышка Время. Величина популяции Дрейф и Инбридинг Пониженная гетерозиготность Возможность закрепления новых признаков
Генетический дрейф Популяционные волны как фактор микроэволюции Эффект бутылочного горлышка Возможные примеры эффекта бутылочного горлышка Северный морской слон 20 особей в 1890 – сейчас более 30 тысяч При исследовании аллоферментов не обнаружено гетерозиготности в 24 локусах у 159 животных и в 43 локусах у 67 животных в другом исследовании. В то же время у южного морского слона, численность которого не изменялась так сильно, средняя гетерозиготность по 35 локусам составляет 0. 032. Гепард Не обнаружено гетерозиготности в 52 локусах Может быть, этот вид тоже прошел «бутылочное горлышко» ?
Эффект основателя Образование уникального генофонда при основание новой популяции маленькой группой особей, в которой частоты генов могут заметно отличаться от материнской популяции просто по воле случая (E. Mayr ) Примеры эффекта основателя Некоторые генетические заболевания в популяциях человека В XVI-XVII веках маленькие группы европейских колонистов основывали поселения за океаном. Франция – Квебек (Канада); Дания – Южная Африка. Некоторые генетические болезни, редкие в Европе, встречаются в этих популяциях гораздо чаще. Большинство из 10– 20 тысяч носителей заболевания “porphyria variegata”, вызывающего нарушения метаболизма гемоглобина, являются потомками одной пары, прибывшей в Кейптаун в 1688 г. Майна Расселилась по многим океаническим островам Каждую популяцию основывали немногие птицы Не обнаружено гетерозиготности при исследовании изоферментных локусов Генетический дрейф
Acridotheres tristis
Эффект основателя Генетический дрейф
Эффект основателя Генетический дрейф
Мутационное давление Мутация – ненаправленное, случайное изменение генетического материала, происходящее спонтанно или под воздействием особых физических, химических или биологических факторов. По характеру воздействия на генотип выделяют: Геномные мутации : анеуплоидия и полиплоидия. Хромосомные мутации Инверсии Реципрокные транслокации Делеции Дупликации (инсерционные транслокации) Точковые (генные) мутации
Пара- и перицентрические инверсии Реципрокные транслокации Метацентрическая хромосома может разделиться на две акроцентрические за счет транслокации с небольшой донорской хромосомой.
По характеру изменения в гене точковые мутации можно разделить на несколько типов: 1. Миссенс-мутация (missense mutation). 2. Мутации со сдвигом рамки считывания (frame shift mutation). 3. Нонсенс-мутация (nonsense mutation). 4. Синонимическая сеймсенс-мутация (samesense mutation).
Организм и ген Частота мутаций I. Многоклеточные организмы Частота на гамету Homo sapiens Альбинизм 3*10 -5 Гемофилия А 1, 3*10 -5 Гемофилия В 5, 5*10 -7 Хорея Гентингтона 2*10 -6 Drosophila melanogaster Желтая окраска тела 12*10 -5 Окраска глаз 4*10 -5 Черная окраска тела 2*10 -5 Zea mays Окраска растения 4, 9*10 -4 Ингибитор окраски 11*10 -5 Сахаристый эндосперм 2*10 -6 Сморщенные семена 1*10 -6 Восковидно-крахмалистый эндосперм 0 на 1, 5 млн. проверенных гамет II. Бактерии Частота на 1 клетку на одно деление Escherichia coli Сбраживание лактозы 2*10 -7 Потребность в гистидине 4*10 -8 Устойчивость к фагу Т 1 2*10 -8 Чувствительность к стрептомицину 10 -8 – 10 —
Частота возникновения мутаций у двух близкородственных видов Drosophila Частота мутаций Вид хромосома II хромосома Ш D. willistoni D. prosaltans 2, 2 * 10 -5 1, 1 * 10 -5 3, 0 * 10 -5 2, 1 * 10 —
• Вероятность сохранения и фиксации единичных и повторных мутаций R. Fisher (1930) — Вероятность сохранения единичной мутации: Таким образом, только повторяющиеся мутации могут иметь значение для эволюции Поколения Нейтральная мутация Мутация с 1% преимуществом 1 0. 6321 0. 6358 3 0. 3741 0. 3803 31 0. 0589 0. 0687 127 0. 0153 0. 0271 В пределе 0. 0000 0.
• Равновесие между прямыми и обратными мутациями
• Влияние потока генов на генофонд популяций: – Привнесение новой генетической изменчивости – Нивелирование различий между популяциями – Противодействие возникновению локальных адаптаций Водяные змеи из озера Эри
Расстояние, м Частота, % все ящерицы самцы 0— 5 60, 1 61, 2 6— 10 25, 9 19, 7 11— 30 10, 0 13, 7 31 —60 2, 5 3, 3 61— 100 1, 1 101— 200 0, 4 0, 7 201— 400 0, 07 0, 2 401— 700 0, 03 0, 1 Практическое исследование расселения Расселение Lacerta agilis (Яблоков и др. , 1980)
Расстояние от места выпуска, м Число меченых мух В месте выпуска 123 40 115 80 116 120 91 160 67 200 65 240 67 280 55 320 45 360 29 400 35 440 19 480 11 520 9 560 2 600 4 640 3 680 11 720 6 760 6 800 2 Практическое исследование расселения Расселение Drosophila pseudoobscura , (Powell et al. , 1976 )
Практическое исследование расселения Расселение Scleroporus olivaceus (Blair, 1960)
Расстояние, на которое происходит расселение, м Интенсивнос ть миграции, m Место выпуска 0, 517 19— 30 0. 168 50— 60 0, 084 90— 100 0, 039 128— 158 0, 041 234— 274 0, 028 378 — 408 0. 029 443— 512 0, 036 638 — 768 0, 052 871— 932 0, 075 Средний радиус индивидуальной активности – среднеарифметическое между местом выпуска и поимки. Интенсивность миграции обозначается показателем m и варьирует от 0 (полное отсутствие иммиграционных аллелей) до 1 (полное затопление популяции иммигрантами за одно поколение). Интенсивность иммиграции у Drosophila nigrospiracula (Johnson & Heed, 1976)
Изменение частоты аллеля за одно поколение под действием потока генов составит: ∆ q = m (q m – q n ) Новая частота аллеля в популяции с учетом иммигрантов одного поколения составит: q n +1 = m (q m -q n ) + q n – исходная частота аллеля в популяции q m – частота аллеля среди мигрантов q n +1 — частота аллеля в смешанной популяции
Поток генов способен достаточно быстро с эволюционной точки зрения уравнять частоты аллелей в смежных популяциях.
Какую роль он играет в более широком масштабе? Генетически эффективный поток генов будет значительно более медленным и менее эффективным по сравнению с процессом расселения. Исследования расселения и потока генов в большинстве случаев касаются изменений происходящих на протяжении жизни одного поколения. Корректная аппроксимация этих данных на длинный ряд поколений едва ли возможна.
Исследования тлей • Økland, 1955: Сосновая тля Cinara piceae Panz. была в большом количестве найдена на Шпицбергене, в 1 300 километрах от предположительного места расселения.
Расселение дрозда рябинника Turdus pilaris с помощью ветра Может ли генетически эффективный поток генов распространить вариации по обширной популяционной системе?
• Inger (1954): расселение с помощью ветра возможно и для мелких амфибий. • Oreophryne annulata Взрослые особи достигают 25 мм. Откладывают яйца во влажный мох на стволах деревьев.
Рекомбинация как фактор микроэволюции
Вследствие рекомбинации между пятью генами, каждый из которых имеет по 10 аллелей, может возникнуть полмиллиона генотипов. n rr g
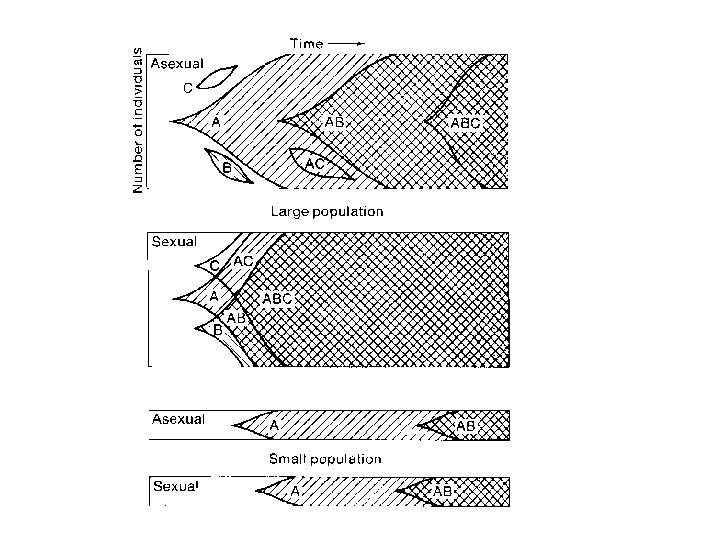
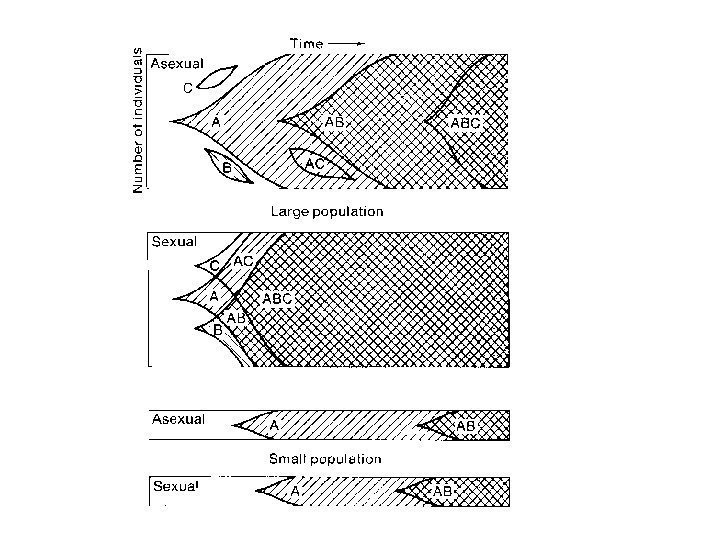
Двукратное преимущество бесполого размножения
Агамные формы — большей частью относительно молодые таксоны – мелкие веточки на филогенетическом дереве, недавно возникшие от обоеполых предков
Принцип красной королевы (Van Valen, 1973) Каждый вид должен эволюционировать как можно быстрее для того, чтобы успеть за изменениями среды В этом соревновании должны побеждать преимущественно виды, размножающиеся половым путем
Проблема сохранения полового размножения Гипотезы, основанные на индивидуальном отборе 1. Гипотеза устранения вредных мутаций Идея, высказанная Мёллером (1964) была развита популяционными генетиками (Crow & Kimura, 1979; Kondrashov, 1983) Половое размножение существует потому, что оно усиливает действие отбора против вредных мутаций (Kondrashov, 1988) Иногда эту гипотезу называют «мутационной теорией полового размножения» Основные идеи: Преимущество полового размножения состоит в том, что оно увеличивает число вредных мутаций, устраняемых в результате одной смерти Если особь размножается путем клонирования, то одна смерть среди ее потомков устраняет только один плохой ген Если же она размножается половым путем, то одна смерть может устранять два и даже много плохих генов Поэтому приспособленность выживающих потомков может в среднем повышаться
• Полиморфизм и полифенизм Генетический полиморфизм популяций
• Различные понимания термина “полиморфизм” в биологии • Популяционно-генетическое определение понятия “генетический полиморфизм” Ford (1940): “ Генетический полиморфизм – это наличие в популяции одновременно двух или более различающихся генотипов (генетических форм) в таком соотношении, что самая редкая из них не может поддерживаться только путем периодических мутаций” Cavalli-Sforza & Bodmer (1971): “ Генетический полиморфизм – это нахождение в популяции двух или более аллелей одного локуса с частотами, поддающимися оценке” Генетический полиморфизм популяций
Генетический груз популяций соответствует той доле, на которую приспособленность популяции оказывается меньше приспособленности оптимального генотипа (Crow, 1958). maxmax W WW L
Мутационный груз (mutation load) Груз, вносимый оказавшимися не на месте особями (misplaced-individual load) Сегрегационный груз (segregation load) При p=q=0. 5 и S=0. 2 в 1000 локусов вероятность дожить до репродуктивного возраста для отдельной особи составляет 4. 3 * 10 -6 Субституционный груз или груз замещения (substitution load)
Haldane, 1957, 1960 : понятие плата за отбор – общее число случаев генетической гибели в процессе полного замещения одного гена. Число гибнущих особей не связано с интенсивностью отбора, а определяется исходной частотой аллеля. От интенсивности отбора напрямую зависит время, необходимое для замещения одного аллеля другим.
Суммарный субституционный груз При p=q=0. 5 зависимость между интенсивностью отбора и числом поколений n, необходимых для замещения аллеля, приблизительно описывается равенством s = 30/n , где 30 – «цена» единичного генного замещения. Таким образом, если s=0. 1, то n=300. С точки зрения Холдейна, для возникновения нового вида достаточно аллельных замещений в 1000 локусах, на что потребуется не менее 300 тысяч поколений, причем одновременно в популяции могут идти замены не более чем по 12 локусам.
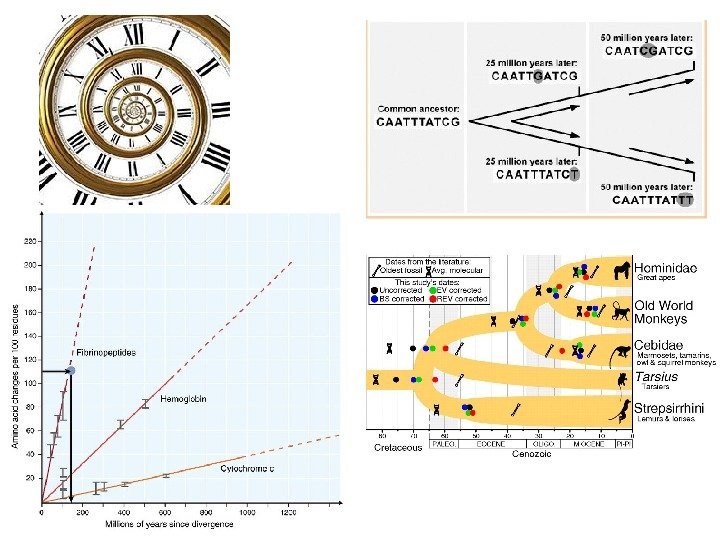
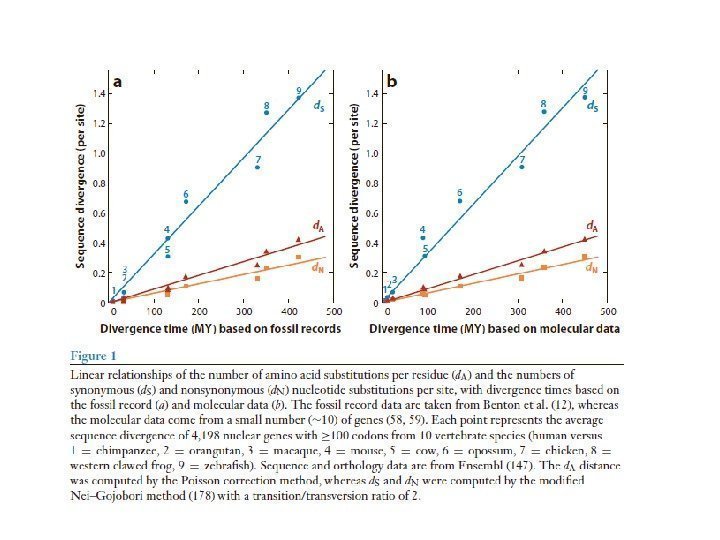
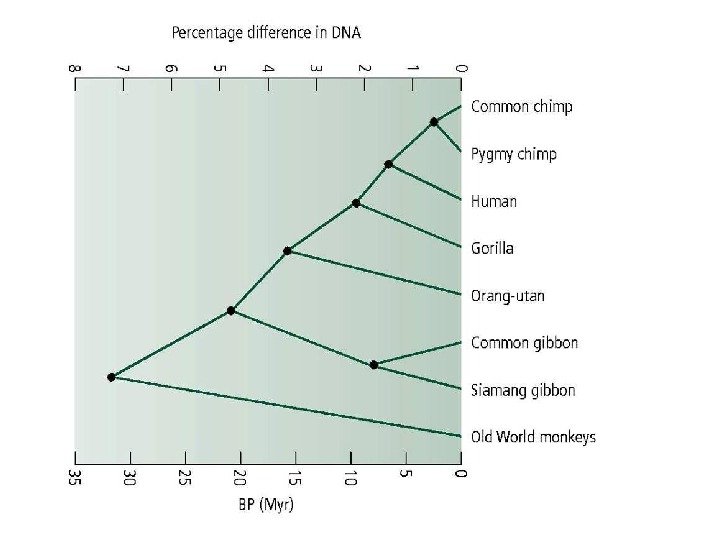
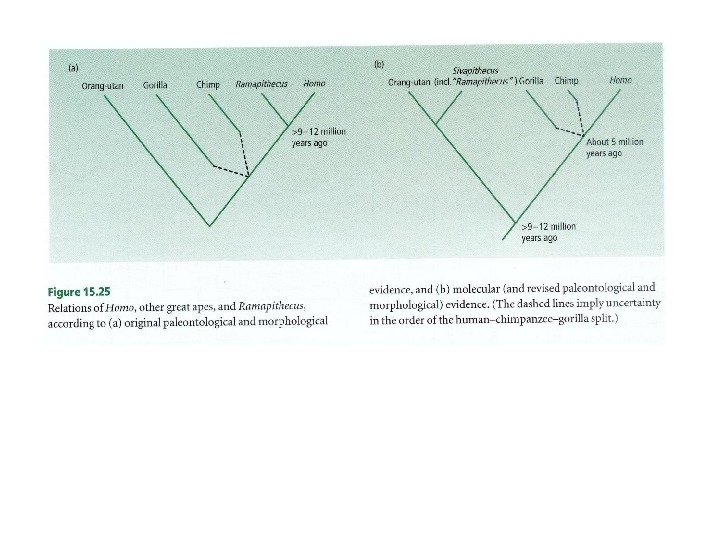
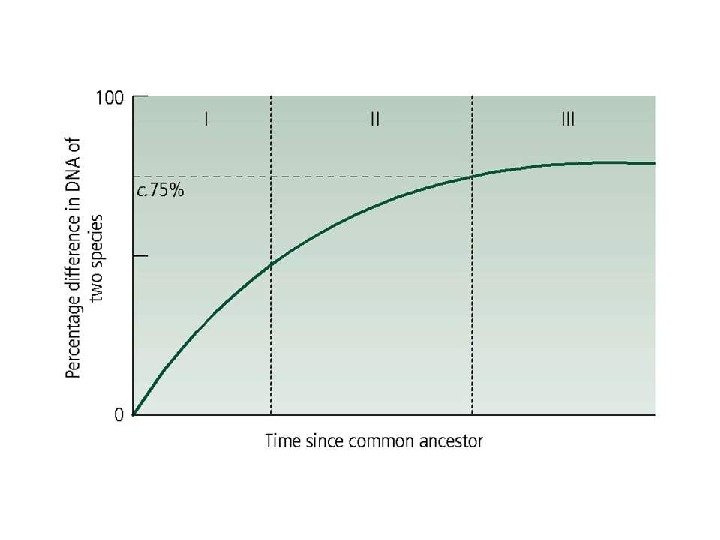
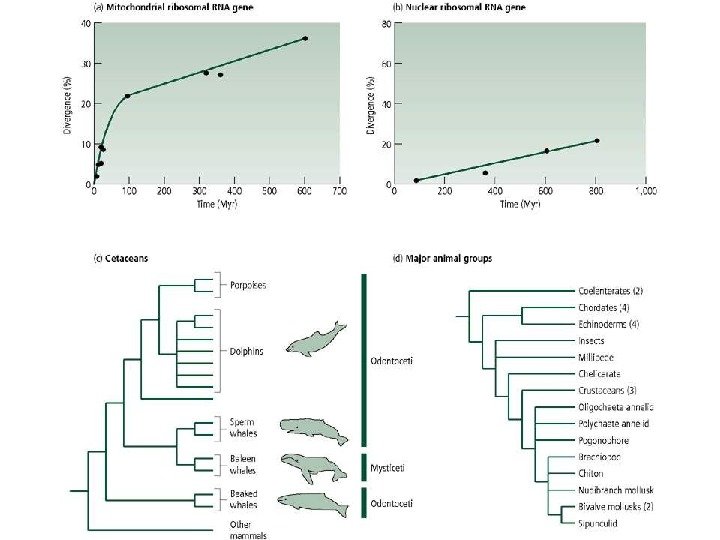
Вскрытие огромных пластов генотипической изменчивости в 60 -х годах XX века стали предпосылкой для формирования концепции нейтральной эволюции Одно из следствий признания селективной нейтральности основной массы мутаций — представление о равных скоростях аминокислотных замен в гомологичных белках разных организмов (молекулярные часы эволюции).
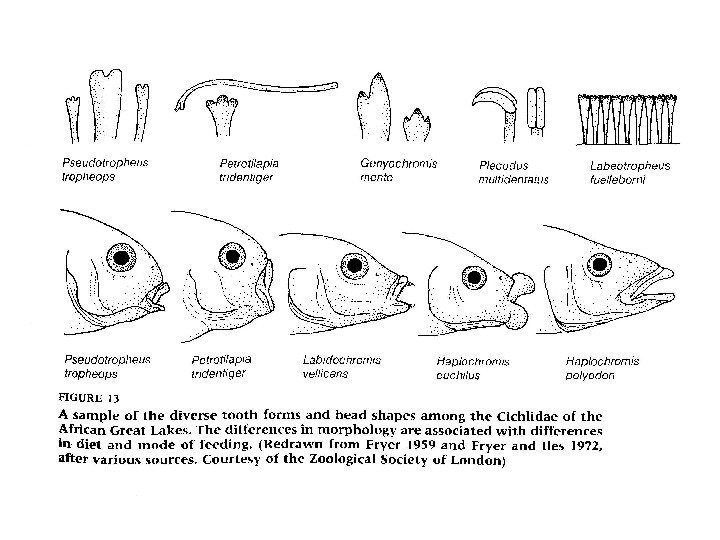
Соотношение генотипа и фенотипа Возможность изменения генотипов в ответ на отбор в значительной степени ограничена общими для родственных организмов механизмами развития. Характер экспрессии генетической изменчивости может быть обусловлен двумя причинами: эволюционно обусловленными и специфическими для изучаемого таксона механизмами развития; взаимодействием организмов с окружающей средой.
Рисунок на крыльях бабочек семейства Nymphalidae Мутации в определяющих рисунок локусах приводят к изменениям в пределах накладываемых рамок, но не к чему-либо абсолютно новому.
Один генотип может формировать несколько фенотипов адекватно влиянию среды Растворенные в воде молекулы, свидетельствующие о присутствии хищников, таких как Notonecta и личинки Chaoborus , вызывают развитие у Daphnia хвостовых шипов, панциря и шейных зубцов. Морские желуди рода Chthamalus реагируют на присутствие хищных моллюсков Acanthina изменениями в развитии, образуя склоненную форму (с расположенным практически перпендикулярно отверстием).
Какие методы анализа характера экспрессии генов используются для корректной оценки роли генетической составляющей, механизмов развития и влияния среды? Выявление нормы реакции. Выявление степени канализированн ости признака Оценить, насколько канализированным является тот или иной признак можно лишь в сравнении.
Насколько адаптивна фенотипическая пластичность? Один из путей определения – произвести взаимный обмен сравниваемых популяций. Исследование двух изолированных популяций лягушек, одна из которых адаптирована к низкой, а другая – к высокой температуре. Лягушки первой популяции развивались дольше и вступали в репродуктивный период достигая в среднем больших размеров, по сравнению с высокотемпературной популяцией. Мы сможем назвать сроки развития адаптивными, если сроки развития местных лягушек приведут к увеличению приспособленности по сравнению с привнесенными из второй популяции лягушками.
Фенотипически сходные организмы могут различаться генотипически. Инфузории комплекса видов Tetrahymena pyriformis неотличимы друг от друга морфологически. Генетические различия между видами свидетельствуют о том, что отдельные виды комплекса эволюционировали независимо в течение более чем 100 миллионов лет. Напротив, генотипически сходные организмы могут различаться фенотипически. Если трохофора эхиуриды Bonellia оседает на нормальный субстрат, она развивается в самку, однако, при оседании личинки на самку, метаморфоз приводит к формированию сильно редуцированного самца.
Какова роль регуляторных генов в природных популяциях? . Появление глазчатых элементов рисунка контролируется геном distal-less. Ген есть у всех бабочек, а его экспрессия в данной ячейке крыла приводит к формированию глазчатого элемента рисунка. Такие регуляторные гены создают обширные возможности для формирования ошеломляющего разнообразия.
Сезонный полифенизм бархатницы Bicyclus anynana При развитии в теплый сезон дождей формируются крупные глазки, а в холодный сухой сезон – маленькие. Критическим фактором для экспрессии distal-less оказалась температура содержания куколки.
Глюкозинолаты: Phyllotreta cruciferae, P. striolata vs Capparidaceae, Tropaeolaceae & Limnanthaceae
Алкалоиды (соланин, никотин…) Manduca sexta, M. quinquemaculata vs Nicotiana spp.
Acalymma vittatum (Fabricius) Вредитель огурцов, питание стимулируют содержащиеся в тканях кукурбитацины (тетрациклические тритерпеноиды)
Abies balsamea : juvabione
Caryedes brasiliensis (Bruchidae) vs Dioclea megacarpa (Fabaceae)
Апосематическая окраска видов, питающихся на Asclepias sp.
Talking trees: Populus, Acer, & Salix spp. vs Malacosoma californicum pluviale
Monostegia abdominalis vs Lysimachia terrestris
Chrysolina quadrigemina vs Hypericum perforatum
Концепция адаптации: основные подходы и проблемы Три значения понятия «адаптация»: 1. Адаптация как эволюционный процесс 2. Адаптация как состояние = “Адаптированность” 3. Адаптация как приспособление в онтогенезе = Фенотипическая пластичность
Концепция адаптации: основные подходы и проблемы Основополагающая роль адаптации в эволюции История представлений о соотношении эволюции, адаптации и отбора ЭВОЛЮЦИЯ = АДАПТАЦИЯ ?
«Front legs a puzzle: how Tyrannosaurus used its tiny front legs is a scientific puzzle; they were too short even to reach the mouth. They may have been used to help the animal rise from a lying position. «
Cepaea nemoralis
Основные методы изучения адаптаций Три этапа исследования: 1. Выделение признаков, генетических или фенотипических вариантов 2. Создание гипотезы об их адаптивности 3. Проверка гипотезы
Основные методы изучения адаптаций Три метода изучения адаптаций: 1. Корреляционный метод 2. Экспериментальный метод 3. Сравнительный метод
Silberglied et al. (1980): Гипотеза о расчленяющем характере окраски бабочки Anartia fatima
Двукратное преимущество бесполого размножения
Агамные формы — большей частью относительно молодые таксоны – мелкие веточки на филогенетическом дереве, недавно возникшие от обоеполых предков
Принцип красной королевы (Van Valen, 1973) Каждый вид должен эволюционировать как можно быстрее для того, чтобы успеть за изменениями среды В этом соревновании должны побеждать преимущественно виды, размножающиеся половым путем
Почему соотношение полов обычно близко к 50: 50% ? Многие биологи уверены в том, что ответ на этот вопрос дает хромосомное определение пола Однако это не так по следующим причинам 1. У многих видов с хромосомным отношением пола соотношение полов сильно отклоняется от 1: 1 2. У видов с иными механизмами определения пола соотношение полов в популяциях обычно бывает близко к 1: 1 Можно утверждать, что естественный отбор мог бы создать практически любое соотношение полов, если бы это было необходимо, даже при хромосомном определении пола
Естественный отбор и соотношение полов Теория Р. Фишера (Fisher, 1930) Основную роль в регуляции соотношения полов играет отбор на уровне особей Суммарная репродуктивная ценность самцов всегда равна таковой у самок в каждом поколении вне зависимости от их численного соотношения Поэтому репродуктивный успех самок и самцов зависит от их численности в популяции: чем больше особей какого-то пола, тем меньше их индивидуальная приспособленность Это типичный частото-зависимый отбор Точка равновесия при соотношении полов 50: 50% В этой ситуации приспособленность самок в точности равна приспособленности самцов 100% самцов 50: 50% 100% самок. W самцов W самок. Точка равновесияw
Physogastric female of the mite Pyemotes ventricosus
Естественный отбор и соотношение полов Влияние числа яиц, отложенных первой самкой, на долю самцов в потомстве второй самки у наездника Nasonia vitripennis. Необычные соотношения полов
Естественный отбор и соотношение полов Необычные соотношения полов Локальная конкуренция при спаривании Если братья конкурируют за оплодотворение сестер то соотношение полов сдвигается в сторону преобладания самок Количество ресурсов и конкуренция Социальный ранг У видов с полигинией, самки с высоким социальным статусом должны производить больше сыновей, поскольку они достигну высокого социального положения и оставят больше потомства Самки с низким социальным статусом должны производить больше дочерей, поскольку репродуктивный успех самок почти не связан с их статусом
Влияние социального ранга самок красного оленя на долю сыновей и дочерей в их потомстве
Виды и видообразование На практике виды различают потому, что внутривидовая изменчивость по ряду признаков не перекрывается межвидовой. Систематики используют все типы признаков, будь то морфологические, этологические или генетические.
Современные представления о виде Критерии вида: морфологический физиологический биохимический экологический этологический молекулярно-генетический репродуктивный Относительность всех критериев. Отсутствие абсолютного и универсального критерия.
Вершина гипопигия D. melanogaster D. simulans Фаллус D. orena D. erecta D. yakuba. Сложности возникают в тех случаях, когда границы между видами не слишком хорошо очерчены.
# # # ## # # # ## # # 2 n = 4 62 n = 5 4# A v a i l a b l e n a m e s Виды-двойники в группе полевок Microtus arvalis. Виды-двойники (sibling species)
Corvus corone и ее два подвида: С. corone (Западная Европа) и C. cornix (Восточная и Северная Европа) Гибридная зона, в пределах которой можно встретить все переходные формы, cоставляет от 20 до 200 километров в ширину. Она, по-видимому, стабильна и сохранилась со времени последнего оледенения.
Краткая история представлений о виде Типологическая концепция вида (приписывается К. Линнею) Номиналистическая концепция вида (приписывается Ж. -Б. Ламарку) Противоречивость взглядов Ч. Дарвина на понятие «вид» Концепция вида в конце XIX – начале XX вв. : «жорданоны» и «линнеоны» Формирование концепции политипического вида
БИОЛОГИЧЕСКАЯ КОНЦЕПЦИЯ ВИДА Виды – группы природных популяций, которые реально или потенциально способны к скрещиванию и репродуктивно изолированы от других таких групп (Mayr, 1963). Основные положения – Виды определяются не различиями, а репродуктивной обособленностью от других видов – Виды состоят из популяций, а не из особей – Виды можно определять более адекватно, исходя из их отношений к популяциям других видов (изоляция), чем на основании отношений между особями в пределах вида Вид = репродуктивное сообщество. “Вид –это защищенный генофонд”(Майр)
Сложности : 1. Организмы, не размножающиеся половым путем Aspergillus niger Taraxacum officinale 2. Виды размножаются половым путем, но репродуктивные барьеры не очень сильны. 3. Биологическая концепция может быть применена лишь к современным организмам. 4. Критерий изоляции очень трудно использовать на практике и его не применить к географическим изолятам
ФИЛОГЕНЕТИЧЕСКАЯ КОНЦЕПЦИЯ ВИДА Вид – монофилетическая группа, состоящая из наименьшей распознаваемой группы организмов, внутри которой наблюдается преемственность поколений (Cracraft, 1983). Каждая такая группа должна характеризоваться набором продвинутых черт, отличающих ее от других подобных групп. Сложности: Остается не вполне ясным, какими продвинутыми чертами должна обладать группа, чтобы называться видом.
ВИДООБРАЗОВАНИЕ Два вопроса относительно механизмов видообразования вызывают наибольшие дискуссии: возникновение репродуктивной изоляции роль географической изоляции в видообразовании
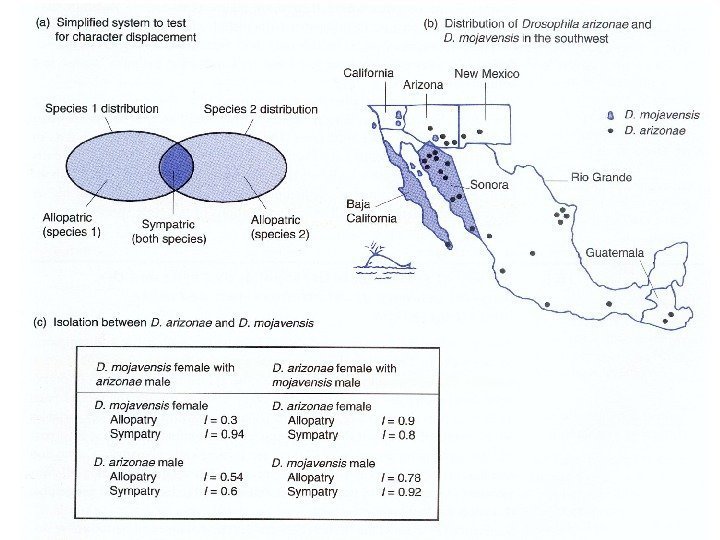
Пространственные взаимоотношения между локальными популяциями в значительной степени определяют степень скрещивания. 1. Аллопатрическими называют популяции, обитающие в разных местах. 2. Симпатрическими называют две или несколько популяций, обитающих в одной и той же области. Смежная симпатрия. Термин применяется к популяциям или экологическим расам одного вида, которые населяют различающиеся, хотя и примыкающие друг к другу местообитания. Биотическая (истинная) симпатрия. Популяции, живущие в одном и том же местообитании. 3. Парапатрическими называют популяции смежные, но не перекрывающиеся и не скрещивающиеся между собой.
Аллопатрическое Парапатрическое Симпатрическое. Основные типы видообразования
Основные формы репродуктивной изоляции. ПРЕЗИГОТИЧЕСКИЕ Экологическая изоляция Этологическая изоляция Механическая изоляция Несовместимость гамет ПОСТЗИГОТИЧЕСКИЕ Нежизнеспособность гибридов Стерильность гибридов Разрушение гибридов Изолирующие механизмы, препятствующие обмену генами между популяциями: пространственные репродуктивные
Концепция биологического вида Географическая и биологическая изоляция между популяциями и видами.
Механизмы биологической изоляции Презиготическая изоляция: экологическая Quercus coccinea предпочитает сырые, плохо дренируемые почвы Quercus velutina предпочитает сухие, хорошо дренируемые почвы
Bufo americanus размножается весной раньше, чем Bufo fowleri. Механизмы биологической изоляции Презиготическая изоляция: экологическая
Anas plathyrhynchus. Anas acuta Свободно скрещиваются в неволе, гибриды всех поколений фертильны. Однако, в природе гибриды очень редки. Особи предпочитают партнеров своего вида. Гибридные особи при попытках спаривания проявляют промежуточное поведение, и поэтому их отвергают партнеры обоих видов. Механизмы биологической изоляции Презиготическая изоляция: экологическая этологическая
Механизмы биологической изоляции Презиготическая изоляция: экологическая этологическая механическая Xenopsylla cheopis Stenoponia vlasovi
Мужские и женские гаметы разных видов морских ежей не вступают в половой процесс, поскольку белковые рецепторы на мембране женских гамет чувствительны только к белкам сперматозоидов своего вида. Механизмы биологической изоляции Презиготическая: экологическая этологическая механическая гаметическая
При скрещивании разных видов леопардовых лягушек из комплекса Rana pipiens зиготы погибают, вероятно, потому что хромосомы не образуют пары в первом же мейозе XМеханизмы биологической изоляции Постзиготическая изоляция: нежизнеспособность гибридов.
При скрещивании Rana pipiens и Rana sylvatica гибриды погибают на стадии ранней гаструлы. ? X Механизмы биологической изоляции Постзиготическая изоляция: нежизнеспособность гибридов.
Equus caballus X Equus asinus = Мул. X Механизмы биологической изоляции Постзиготическая изоляция: нежизнеспособность гибридов. стерильность гибридов.
Лютики и их гибриды. Ranunculus millanii растет во влажных местообитаниях, а Ranunculus dissectifolius – в сухих. Их гибриды не могут конкурировать с родительскими видами в их местообитаниях и поэтому занимают промежуточные, нарушенные стации. X Ranunculus millanii Ranunculus dissectifolius. Механизмы биологической изоляции Постзиготическая изоляция: нежизнеспособность гибридов. стерильность гибридов. разрушение гибридов.
Drosophila pseudoobscura и Drosophila persimilis могут скрещиваться в лаборатории. 1. Гибриды F 1 вполне жизнеспособны. Самки полностью фертильны, но самцы стерильны. 2. При скрещивании самок F 1 с самцами родительских видов потомство имеет пониженную жизнеспособность и фертильность. X Drosophila pseudoobscura Drosophila persimilis. Механизмы биологической изоляции Постзиготическая изоляция: нежизнеспособность гибридов. стерильность гибридов. разрушение гибридов.
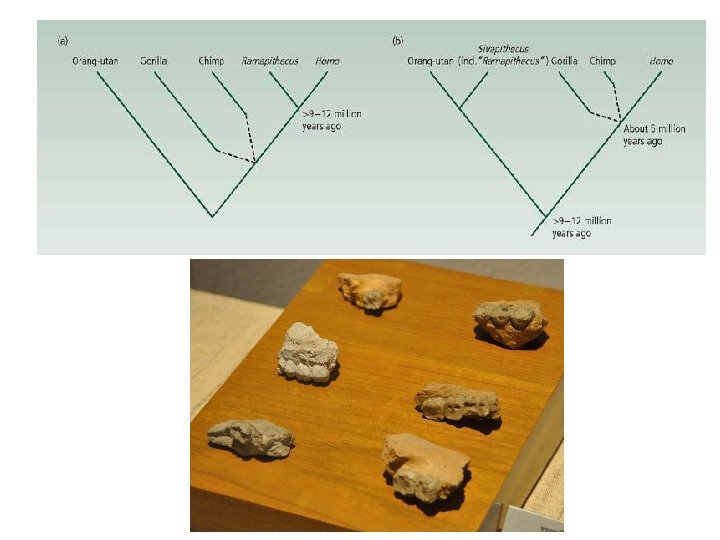
Аллопатрическое видообразование Moritz Wagner (1868): миграция – изоляция – образование нового вида Ernst Mayr (1942 и последующие работы) – сыграл основную роль в разработке теории. Последовательность нарастания различий при дивергенции: Возникновение биологической изоляции между новыми видами – это побочный результат дивергенции в условиях аллопатрии. Локальная раса Географическая раса Аллопатрический полувид Вид
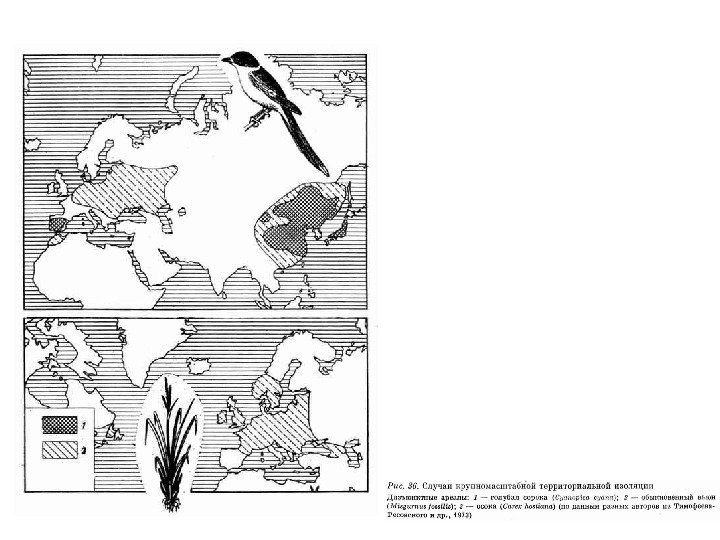
Аллопатрическое видообразование Отправной точкой является разделение единого вида на две или более группы популяций. Подобная ситуация может возникнуть в результате миграции, локального вымирания промежуточных популяций или в силу геологических событий. Барьеры могут быть как географического, так и экологического характера. Кольцевые ареалы
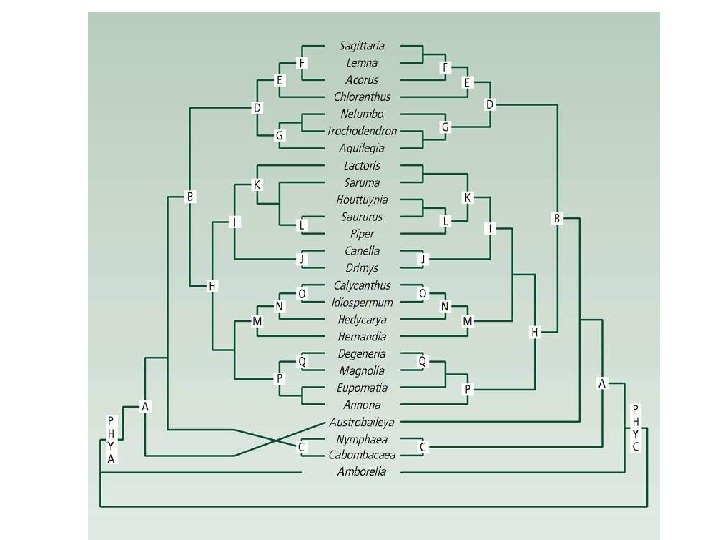
Кольцевые ареалы комплексов видов большой синицы Parus major и саламандры Ensatina eschscholtzii
Перипатрическое видообразование ( Mayr, 1982) = видообразование путем генетической революции (Mayr, 1954, 1963) = квантовое (Grant, 1963) Новый вид формируется из изолированной островной или периферической популяции родительского вида. Небольшие изоляты на периферии ареала, представленные лишь несколькими особями родительской популяции подвергаются жесткому давлению отбора в сочетании с сильным действием дрейфа генов и инбридинга, снимающими гомеостатические буферы популяции. Подавляющее большинство подобных колоний погибнут или сольются с предковой популяцией.
Перипатрическое видообразование – это специальный случай аллопатрической модели. Это быстрое видообразование в условиях небольшой изолированной популяции. Ареал вида
Перипатрическое видообразование – это специальный случай аллопатрической модели. Это быстрое видообразование в условиях небольшой изолированной популяции. Ареал вида Перипатрические изолированные популяции
Перипатрическое видообразование – это специальный случай аллопатрической модели. Это быстрое видообразование в условиях небольшой изолированной популяции. Ареал вида Перипатрические изолированные популяции
Перипатрическое видообразование – это специальный случай аллопатрической модели. Это быстрое видообразование в условиях небольшой изолированной популяции. Ареал вида Перипатрические изолированные популяции Аллопатрические виды
Вероятные примеры перипатрического видообразования. Шмель Bombus agrorum
Островные расы, происходящие от небольшого числа мигрантов из популяции главного острова, заметно отличаются. Степень их дифференциации значительно выше обычной расовой изменчивости на главном острове, а одна из островных популяций дивергировала до видового уровня. Изменчивость у новогвинейских зимородков группы Tanysiptera hydrocharis-galatea :
Симпатрическое видообразование Ареал материнского вида Симпатрические популяции, изолированные от материнского вида Новые симпатрические виды
Симпатрическое видообразование Два возможных способа симпатрического видообразования. 1. Быстрое недивергентное видообразование: гибридизация полиплоидия хромосомные мутации и т. п. 2. Медленное видообразование путем постепенной дивергенции симпатрических популяций. наиболее дискуссионный до сих пор способ видообразования. Два типа симпатрии: Смежная симпатрия дивергентное видообразование в принципе возможно. Биотическая симпатрия дивергентное видообразование крайне маловероятно, поскольку возможно только на базе ассортативного скрещивания.
Возможные механизмы смежно-симпатрического видообразования. 1. Отбор, возникающий в силу использования различных ресурсов. Например, паразитические формы могут переключиться на другого хозяина, если он недавно появился или его численность сильно возросла. Подобная ситуация исследовалась на мухах пестрокрылках Rhagoletis polmonella, личинки которых поедают плоды боярышника.
Дивергентное симпатрическое видообразование Расы по хозяину и по кормовому растению у насекомых как возможные примеры симпатрического видообразования. Боярышниковая пестрокрылка Rhagoletis pominella в Северной Америке Исходная форма – на диком боярышнике. С 1866 г. стала заражать завезенные в США яблони. Образовалась яблоневая форма, а затем и вишневая. Все три формы спариваются на своем кормовом растении и откладывают яйца на него. Кроме того, они заметно изолированы сезонно.
Дивергентное симпатрическое видообразование Фенология трех форм боярышниковой пестрокрылки Rhagoletis pominella в Северной Америке
Дивергентное симпатрическое видообразование Различия пищевой специализации в группах близких видов: Аллопатричные виды нередко используют одни и те же кормовые растения. Симпатричные виды очень часто отличаются кормовыми растениями, или используют для питания различные части одного растения. Филогенетический анализ некоторых групп монофагов растений позволяет считать симпатрический способ видообразования наиболее вероятным. Например, известны сотни видов фиговых ос, причем каждый питается на своем виде фиговых деревьев.
Возможные механизмы смежно-симпатрического видообразования. 2. Дивергенция сроков цветения у растений. Репродуктивная изоляция у растений может быть следствием дифференциации по времени цветения. Расы большого погремка При различных сроках покоса
• Виды Agrostis и Anthoxanthum могут произрастать около отвалов из шахт несмотря на высокую концентрацию меди, свинца и цинка в почве. На отвалах могут выживать только толерантные формы, в то время как нормальные формы доминируют вокруг копей. Возможность биотически-симпатрического видообразования сопряжена со значительно большими трудностями. Для дивергенции в этом случае необходим сильный дизруптивный отбор, сопровождающийся высокой платой за отбор.
Симпатрическое видообразование пальм на острове Лорд Хау
Сроки цветения Howea belmoreana и H. forsteriana
Видообразование путем полиплоидии играет особую роль у цветковых растений, где, как полагают специалисты, 70 -80 % видов возникли путем полиплоидии. Частота полиплоидных видов составляет 47% у цветковых, только 1, 5% у голосемянных и 95% у папоротников. Полиплоидизация происходит прежде всего у организмов, у которых возможно самооплодотворение и вегетативное размножение. В местах с более жесткими условиями существования процент полиплоидов возрастает и явно коррелирует с географической широтой и высотой обитания над уровнем моря. Внезапное симпатрическое видообразование путем полиплоидизации
Полиплоидия у животных менее распространена , поскольку у животных самооплодотворения и бесполого размножения обычно не бывает. Ряд полиплоидных видов среди животных представлен партеногенетическими самками (жуки, бабочки-чехлоноски и мокрицы). Установлено полиплоидное происхождение многих видов и целых семейств рыб, таких как лососевые, хариусы, некоторые сельдеобразные. Автополиплоидами являются южно-американские лягушки семейства Ceratophrididae. Путем гибридизации произошли многие виды родов Lacerta и Cnemidophorus , они партеногенетические и различаются по числу хромосом. Гибридогенез теснейшим образом связан с полиплоидией. У растений межвидовые гибриды имеются во всех крупных группах. Особо много видов гибридогенного происхождения среди ив, дубов, эвкалиптов, фиалок, водосборов, папоротников рода Pteris. У животных естественная гибридизация выявлена у насекомых, рыб, амфибий и рептилий. У птиц и млекопитающих гибридизация не играет существенной роли.
Фертильный аллотетраплоидный вид Primula kewensis = P. floribunda × P. verticillata
Фертильные тетраплоиные гибриды, репродуктивно изолированные от исходно интродуцированных в Северную Америку родительских пар видов: Tragopogon murus = T. dubius × T. porrifolius Tragopogon miscellus = T. dubius × T. pratensis
Хромосомное определение пола – невозможность полиплоидии. Но возможно образование гибридных партеногенетических видов, состоящих из одних самок. Такие виды известны у некоторых ящериц и лягушек. Аллополиплоидия потенциально возможна у рыб, имеющих гормональное определение пола. Действительно, такие виды были найдены в конце XX века у: Рыбок-шиповок рода Gobitis в Японии и Подмосковье (Васильев, 1985). У лососевых (Кирпичников, 1979). В природе отмечены плодовитые гибриды между разными видами сусликов (Воронцов)Возможности видообразования путем гибридизации и полиплоидии у животных
Видообразование путем интрогрессии.
Примеры видообразования путем интрогрессии Ирисы Iris hexagona, Iris fulva и Iris brevicaulis из штата Луизиана, США
Гибридизация ирисов Iris hexagona, I. fulva и I. brevicaulis в местах контакта рукавов реки с болотами. Примеры видообразования путем интрогрессии
Гибридный вид ириса Iris nelsonii в лесах штата Луизиана, СШАПримеры видообразования путем интрогрессии
Гиногенез и кредитогенез у карпообразных рыб Живородящая рыба Poecilia formosa (ЮВ Техас) – гиногенетический вид, состоящий только из самок, но сперма других видов самцов того же рода стимулирует его икру к развитию. Род Poeciliopsis содержит помимо диплоидных видов (2 n=48) однополые диплоидные (2 n=48) и триплоидные (2 n=72) виды гибридного происхождения. Самки диплоидных однополых видов развиваются из гибридных зигот, но все отцовские хромосомы избирательно элиминируются в ходе овогенеза и зрелые яйцеклетки содержат только гаплоидный материнский геном. Во всех случаях формирование и поддержание гибридных видов у животных возможно лишь благодаря нерегулярным формам полового размножения – партеногенезу, гиногенезу и кредитогенезу.
Парапатрическое видообразование = смежно-симпатрическое или стасипатрическое White (1978) Свои выводы Уайт сделал на основе комплексного исследования 160 видов и подвидов австралийских кузнечиков подсемейства Morabinae. Было найдено 34 транслокации хромосом, закрепившихся в ходе эволюции группы. Основная идея парапатрического видообразования – дивергенция в соответствии с градиентом воздействия среды с последующим усилением изоляции и снижением плодовитости гибридов.
Комплексы видов Vandiemenella viatica и V. scurra (Eumastacidae): гомозиготные по разным транслокациям подвиды занимают смежные территории. На их границах в узкой зоне образуются гетерозиготные гибриды, которые уступают родительским формам по плодовитости и приспособленности. Подобные узкие гибридные зоны известны и для других видов животных, исследовались у жуков долгоносиков Eremochrous oppositus и E. elongatus Алтая, сверчков рода Gryllus Коннектикута. Ширина гибридной зоны должна определяться силой отбора против гетерозигот, хотя в большинстве случаев точные причины узости гибридных зон неизвестны.
Kawakami et al. , 2011: Chromosomal speciation revisited: models of diversification in Australian Morabine grasshoppers (Vandamiella viatica species group)
Транслокации. Хромосомное видообразование Видообразование у растения Clarkia в Калифорнии, США
Хромосомные расы у Peromyscus goldmani. Хромосомное видообразование
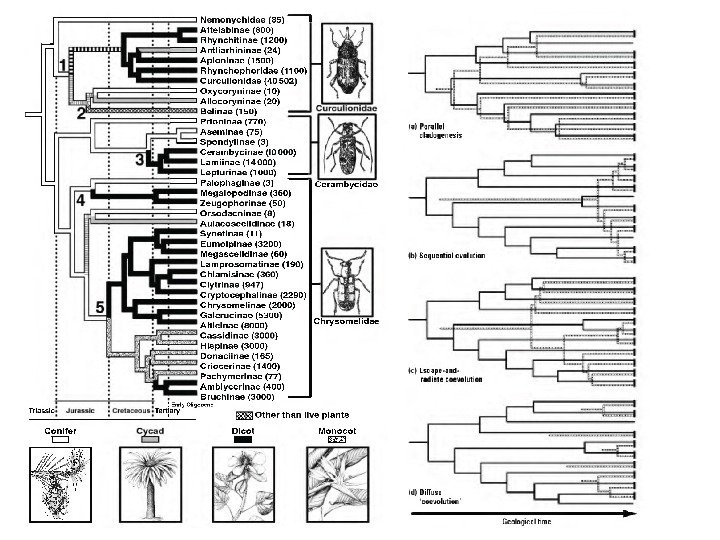
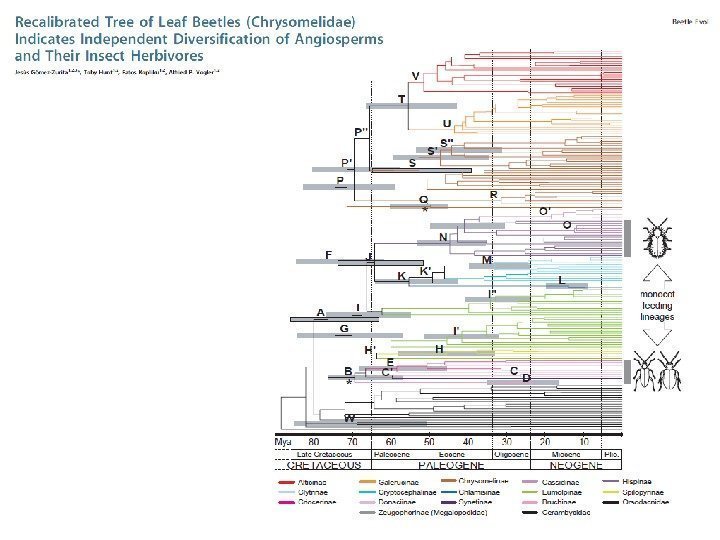
Быстрая кариотипическая эволюция и видообразование у голубянок рода Agrodiaetus (Лухтанов, 2001)Хромосомное видообразование
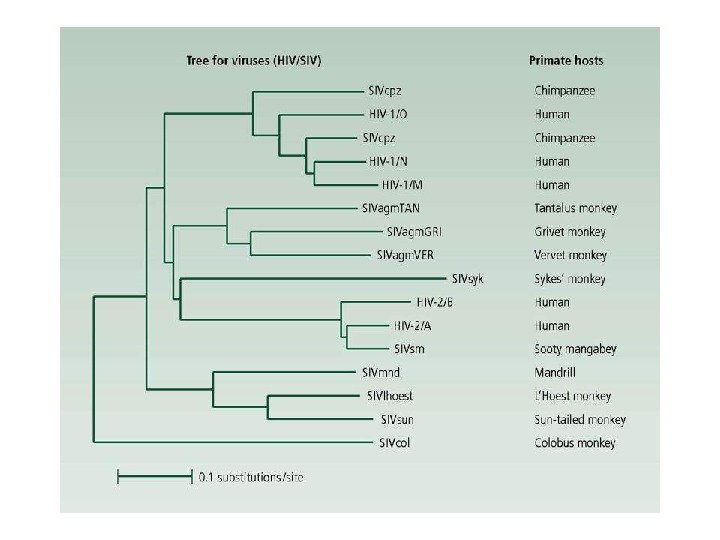
Phylogeny of seven species and three incipient species related to A. gambiae Ayala F. J. , Coluzzi M. PNAS 2005; 102: 6535 -6542 © 2005 by National Academy of Sciences
Geographic distribution of A. gambiae and six other closely related species Ayala F. J. , Coluzzi M. PNAS 2005; 102: 6535 -6542 © 2005 by National Academy of Sciences
Возникновение репродуктивной изоляции Что происходит, если исходно аллопатрические популяции встречаются (вторичная симпатрия) достигнув лишь частичной репродуктивной изоляции? Какой уровень переноса генов приведет к остановке водообразования? Дивергенция последовательностей может оказаться барьером для рекомбинации. Усиление изоляции (Reinforcement ) Поскольку постзиготическая изоляция обходится популяции дорого, Добжанский предположил, что отбор должен благоприятствовать формированию презиготических механизмов изоляции.
Усиление изоляции у бабочек голубянок рода Agrodiaetus
Роль полового отбора Видообразование у дрозофил Гавайских островов тщательно исследовалось с 60 -х годов. Гавайские острова населяют около 800 эндемичных дрозофил. Вероятнее всего, эти острова заселили несколько мух с последующими аллопатрическим видообразованием и вторичной симпатрией. Самцы в этой группе часто сильно различаются, но самки обычно крайне схожи. Генетически эти виды также крайне схожи. Полагают, что отличительные черты самцов – побочные продукты контролирующих брачное поведение генов. Презиготическая репродуктивная изоляция возникла в результате аллопатрических изменений брачного поведения, возможно благодаря эффекту основателя.
Взрывное видообразование цихлид рода Haplochromis в озере Виктория В Виктории обитает порядка 500 -1000 видов цихлид этого рода, возникших менее чем за 13000 лет, то есть последнего оледенения, когда озеро совершенно полностью пересыхало. Показано, что репродуктивную изоляцию между симпатричными видами обеспечивает активный выбор самками самцов соответствующего цвета.
Эксперименты Г. X. Шапошникова с тлями рода Dysaphis Купырь Anthriscus (основное кормо- вое растение) Бутень Chaerophyllum maculatum Бутень Chaerophyllum bulbosum
Эволюционные сценарии
• Эволюционная систематика Классическая эволюционная таксономия призывает к вкладыванию максимального количества эволюционной информации в биологические классификации Ernst Mayr, Gorton Linsley & Robert Usinger (1953) Methods and principles of Systematic Zoology
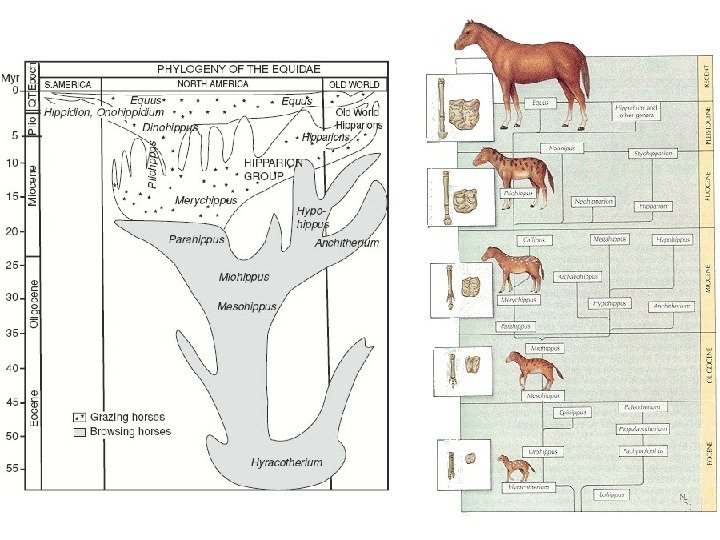
GINA C. GOULD and BRUCE J. Mac. FADDEN,
Matthew, 1903: Mac. Fadden, 1992:
ФЕНЕТИКА Основные идеи: По возможности полностью исключить эволюционные суждения, поскольку филогения для подавляющего большинства групп неизвестна; Методы эволюционистов недостаточно точны и количественны; Отражающие филогению классификации по своей природе предназначены для очень специальных целей.
Фенетика Эволюционная систематика Кладистика Тип данных Матрица дистанций между таксонами Дискретные признаки Метод группировки Общее сходство Специфическое сходство Тип диаграммы Фенограмма Эволюционное дерево Кладограмма Иерархический уровень определяется Количеством различий Наличием уникальных черт Учитывается ли степень различий Да да нет
КЛАДИСТИЧЕСКИЙ АНАЛИЗ Ядро кладистического анализа составляет представление о том, что все таксоны должны быть только монофилетическими. Основой филогенетической систематики или кладистики является использование апоморфных или продвинутых (прогрессивных) состояний признаков для группировки таксонов. Термин апоморфия или апоморфный признак противопоставляется плезиоморфии или исходному, примитивному состоянию признака.
Монофилетическим является таксон, который включает общего предка и все филогенетические ветви, идущие от этого предка к каждому члену этого таксона, т. е. включает всех потомков данного предка. Парафилетическим является таксон, включающий в себя предка, общего для всех членов таксона, но не всех потомков этого предка. Полифилетическим называется таксон, не включающий в себя предка, общего для всех членов данного таксона.
Гомология – синапоморфия – гомоплазия. Современное понимание критериев гомологии формализовал Remane (1952): критерий положения, критерий особого строения, переходные формы.
Параллелизмы – конвергенции. Принято считать, что если наблюдается прямая зависимость приобретаемого признака от функции, то мы имеем дело с конвергенцией. Если же отчетливо проявляется зависимость приобретаемого признака от общих унаследованных особенностей организма, то это параллелизм. Присоски на пальцах у древесных форм бесхвостых амфибий Расширения на концах пальцев у различных древесных ящериц
Параллельные и конвергентные изменения зачастую теснейшим образом переплетаются В качестве отграничивающего критерия используют: ранг группы (отряд); развиваются ли сходные структуры после того как родственные филумы заняли каждый свою адаптивную зону (параллелизм) или когда группы независимо друг от друга вселяются в одну адаптивную зону (конвергенция); признаки, не изменяющие степени сходства или повышающие степень сходства; гомологичны ли изменяющиеся признаки или нет.
Изучение сходства в строении = предполагаемых гомологий у ряда изучаемых таксонов может привести к следующим трем заключениям: 1. Признаки, считающиеся гомологичными, инвариантны в пределах изучаемой группы. Они не подтверждают и не опровергают любой вариант группирования таксонов. 2. Признаки, считающиеся гомологичными, однозначным образом диагностируют таксоны разного ранга. В данном случае гомологичные признаки являются синапоморфиями. 3. Признаки, считающиеся гомологичными варьируют таким образом, что подтверждают различные, несовместимые группы. Противоречащие другу признаки именуются гомоплазиями.
Признаки Tаксоны 1 2 3 4 5 6 7 X
Признаки Таксоны 1 2 3 4 5 6 7 X 0 0 0 0 M 1 1 0 0 1 1 1 N 1 1 1 1 O 1 1 0 0 0 ПРИНЦИП ПАРСИМОНИИ
Таксоны Признаки 1 2 3 4 5 6 7 8 9 10 11 12 Внешняя группа 0 0 0 A 1 0 1 1 1 0 0 0 B 1 1 1 0 0 0 1 0 0 C 1 0 1 1 0 0 0 0 D 1 1 0 0 1 0 E 1 1 0 0 0 1 Таксоны 1 2 3 4 5 6 7 8 9 10 11 Внешняя группа 0 0 0 F 1 1 0 1 1 1 G 1 1 0 0 1 1 H 1 1 0 0 1 1 I 0 1 1 1 0 0 0 J 1 1 0 0 K 1 1 0 0 Таксоны 1 2 3 4 5 6 7 8 9 10 Внешняя группа 0 0 0 0 0 L 1 0 1 1 0 0 M 1 0 1 0 0 0 N 1 1 1 0 0 0 1 O 1 1 0 0 0 1 0 0 P
Прогресс и регресс Обычно прогрессом называют направленное развитие при котором более поздние стадии превосходят более ранние по каким-либо критериям. Одна из довольно популярных точек зрения заключается в том, что запечатленный геологической летописью ход событий — это и есть прогресс. А. Н. Северцов, 1939: Разграничение биологического и морфофизиологического прогресса Критерии биологического прогресса: 1. Увеличение численности 2. Расширение ареала 3. Увеличение числа систематических групп, составляющих данный таксон: видов в роде, родов в семействе и т. д.
Способы достижения биологического прогресса по Северцову: 1. Морфофизиологический прогресс или ароморфоз. Организмы получают в борьбе за существование преимущества общего характера, не ограниченные какой-либо определенной средой. 2. Идиоадаптации , т. е. ограниченные приспособления к среде, при которых организмы не испытывают не значительного усложнения, ни их упрощения. Эволюция направлена в сторону специализации и приспособления к конкретной экологической обстановке. 3. Морфофизиологический регресс , когда организмы переходят к более простым взаимоотношениям со средой, а лишние органы претерпевают редукцию.
Прогресс — это длинный крутой подъем, который ведет ко мне. Ж Сартр. Слова.
Hines et al, 2007: Multigene phylogeny reveals eusociality evolved twice in vespid wasps Exemplar method vs. Groundplan estimation
Алгоритм построения кладограммы методом полного перебора x=3 N = P (2 n-5) x=n
Разнообразие иглокожих на протяжении геологического времени
Четыре представителя иглокожих из нижнего палеозоя А. Геликоплакоидея. Б. Паракриноидея. В. Стилофора. Г. Примитивная эхиноидея. Все эти формы были построены из кальцитных пластинок и обладали амбулакральной системой. В остальном они сильно различались как по типу симметрии, так и по общему строению.
Вероятные причины неполноты геологической летописи Разрывы вызваны полным уничтожением ископаемых остатков в результате эрозии и других процессов или же несохранением когда-то существовавших промежуточных форм. Интерградирующим формам иногда присваивают видовые названия, затемняя таким образом их переходный характер. Эволюционные превращения происходили в небольших географически ограниченных популяциях, после чего новая форма быстро распространялась по более обширному ареалу предкового вида. Предсказания, сделанные Симпсоном на основе «бумажного эксперимента» 1. Ископаемые остатки, относящиеся к крупному разрыву, должны обнаруживаться лишь в очень редких случаях. 2. Ряды последовательных родов должны встречаться чаще, чем ряды последовательных видов. 3. Некоторые крупные таксоны будут вновь обнаруживаться после долгого отсутствия.
Скорости эволюции : филогенетическая — скорость эволюции отдельного признака или комплекса признаков в пределах ветви; таксономическая — скорость, с которой таксоны сменяют друга в эволюции. Три крупные категории скоростей эволюции: горотелические, брадителические и тахителические. Latimeria chalumnae сходна с представителями позднего Карбона, 250 млн. лет. Triops cancriformis – неотличим от известных из триаса ископаемых форм (180 млн. лет) и отнесен к тому же виду.
Различия по последовательностям белков между видами саламандр рода Plethodon свидетельствуют о том, что наиболее древние виды дивергировали от общего ствола более чем 60 миллионов лет назад. Тем не менее, эти виды морфологически отличаются крайне незначительно, размерами, цветом и относительными размерами костей скелета.
Квантовая эволюция — быстрые и значительные эволюционные изменения, сопровождающихся занятием новой адаптивной зоны. В ранний период радиации новых групп, распространяющихся в незанятые адаптивные зоны, возможна тахителия. Во время такой быстрой радиации все линии относительно плохо приспособлены и между ними нет конкуренции. В результате возникают разнообразные линии; одни из них быстро вымирают, тогда как другие линии укрепляют свои позиции в данной адаптивной зоне. Скорости эволюции могут резко изменяться в пределах одной линии.
Скорость изменения отдельного признака. Холдейн предложил термин «дарвин» для обозначения единицы эволюционного изменения размера, равного изменению в е (2, 3) раз за 10 6 лет. В практических целях 1 дарвин можно примерно приравнять к изменению размеров на 0, 001 за тысячу лет, что дает изменение размеров вдвое за 10 6 лет. Средняя скорость увеличения в размерах от самого маленького к самому крупному цератопсидному динозавру (например Triceratops) – 60 миллидарвинов. В тоже время, искусственный отбор в лабораторных экспериментах способен изменять значения признаков за короткий период при скоростях в 60000 дарвинов. В случаях, когда геологическая летопись более полна, видно, что сравнительно небольшие средние скорости маскируют периоды быстрых изменений Изменения размеров радиолярии Pseudocubus vema
Cо времен Дарвина большинство эволюционистов, в том числе сторонники синтетической теории, рассматривают эволюцию как состояние медленного и равномерного изменения, как случай «филетического градуализма» ; подлинные макроэволюционные изменения носят характер вспышек, так что длительные периоды постоянства (или «застоя» ) чередуются с короткими эпизодами быстрого изменения; эпизоды существенного изменения совпадают с видообразовательными событиями. Прерывистое равновесие Punctuated equilibrium (Eldredge & Gould, 1972)
В качестве оригинального примера прерывистого равновесия Eldredge (1971) привел редукцию числа рядов фасеток с 18 до 15 у трилобитов рода Phacops. Представления о генетических процессах в популяциях не поддерживают идею о том, что для эволюционных изменений признаков необходимо видообразование. Более того, для того чтобы показать, что быстрые изменения в геологической летописи сопряжены с видообразованием, необходимо показать, что наряду с новым видом симпатрически обитает и предковая форма. Однако хорошо задокументированных примеров такого рода практически нет.
Трактовка палеонтологических данных может быть различной, в том числе и при оценке того, имеем мы дела с градуальными или прерывистыми изменениями. Реконструкция эволюции первого верхнего моляра у Hyopsodus из раннего Эоцена рассматривалась ее автором (Gingerich, 1976) как пример прежде всего филетического градуализма, однако впоследствии использован Элдриджем и Гулдом для иллюстрации прерывистого равновесия.
Эволюция глаз А – светочувствительная клетка к убомедузы Tripedalia cystophora; B – камерный глаз этого же вида (есть роговица, хрусталик, сетчатка, пигментный слой и диафрагма); С – простой глаз трематоды Multicotyle purvisi ; D — более сложная конструкция, включающая одну пигментную клетку и несколько рецепторных, турбеллярии Bdellocephala brunnea.
Распределение типов глаз в основных таксонах животных
Теоретическая модель оценки времени, необходимого для возникновения сложного камерного глаза путем серии постепенных изменений (Nilsson & Pelger, 1994)
Некоторые варианты строения глаза моллюсков (Conn, 1900)
Рабдомерные и цилиарные фоторецепторы животных
Eyeless (Pax 6) ген дрозофилы
Часто встречающиеся заблуждения о эволюции сложных органов 1. Переходные стадии эволюции сложных органов не могут функционировать. Не может быть половины глаза или половины крыла. 2. Переходных форм нет. 3. Биологи считают, что сложные органы возникают по воле случая. 4. Все особенности сложного органа сконструированы наилучшим образом. 5. Неспособность объяснить возникновение сложного органа подрывает фундамент эволюционной биологии. 6. Гипотеза о сложных органах не может быть протестирована.
Основные формы эволюционного процесса • Прямое Формирование адаптаций под действием естественного отбора (последовательная Al —» А 2—АЗ —> • А 4 и параллельная А 1/В 1 —► А 2/В 2 —> АЗ/ВЗ А 4/В 4. ) • Экзаптация = коаптация = преадаптация • Gene sharing • Дупликация Гомотипия Гомодинамия или сериальная гомология Гомономия
Ambystoma mexicanum
Таблица 6 -1. Типы гетерохронии (Gould, 1977) Изменение сроков Название по системе де Бера Морфологический результат появления соматического признака созревания репродуктивных органов Ускорение Без изменения Акселерация Рекапитуляция (путем ускорения) Без изменения Ускорение Педоморфоз (прогенез) Педоморфоз (путем урезывания) Задержка Без изменения Неотения Педоморфоз (путем задержки) Без изменения Задержка Гиперморфоз Рекапитуляция (путем продления)
Меgаloceros giganteus
Ambystoma mexicanum
Маврикийский пастушок ( Aphanapteryx bonasia ) Маскаренская лысуха (Fulica newtoni)
Трилобиты рода Arthricocephalus , ранний Кембрий
Stropiurichthys robustus
Microdipodops Dipodomys Orthogeomys
Массовые вымирания
Астероидная гипотеза Как свидетельствуют косвенные данные, примерно 65 млн. лет назад на землю упал астероид диаметром 10 км. При его входе в верхние слои атмосферы образовалось облако пыли, сохранявшееся в течение 2 или 3 лет. В морских осадках на границе между мелом и третичным периодом образовался прослой, обогащенный иридием — металлом, относящимся к семейству платины, который редок в земной коре, но содержится в больших количествах в астероидах.
Слабые места: 1. Исчезновение какого-либо вида в одном или нескольких местонахождениях ископаемых не означает, что данный вид вымер. В других местах популяции этого вида могли выжить. 2. Остается неясным, мог ли постулируемый слой пыли подавить фотосинтез в такой степени, чтобы вызвать голод в широких масштабах. 3. Астероидная гипотеза не позволяет объяснить, почему некоторые группы вымерли, а другие выжили. 4. Астероидная гипотеза требует допустить, что все вымирания произошли за короткое время, измеряемое годами или десятилетиями, непосредственно следовавшими за катастрофой.
Палеонтологическая летопись позволяет предполагать эпизодическое возникновение периодов массового вымирания, во время которых многие прежние группы гибнут, после чего происходит развитие и адаптивная радиация новых групп. Существенная черта эпизодического вымирания — высокая скорость этого процесса, наблюдаемая в совершенно не связанных между собой группах примерно в одно и то же время. От поздней перми и до конца плиоцена кривая вымирания семейств образует 12 максимумов. Эти максимумы вымирания расположены со средним интервалом 26 млн. лет. Периодичность геологических кризисов—около 180 и около 30 млн. лет— наводит на мысль о том, что геологические кризисы возникают в результате взаимодействия Земли с другими космическими телами. Первый период соответствует галактическому году — периоду обращения Солнечной системы вокруг центра Галактики, второй — периоду ее вертикальных колебаний около галактической плоскости.