2fb55a413edc35f9abb6c940741a9e92.ppt
- Количество слайдов: 84



BIOINFORMATIKA BIOLÓGIA “A valaha élt kutatók 99%-a kortársunk” Az adatokra is igaz információs forradalom

INFORMATIKA - információk megfejtése új információk produkálása - adatok feldolgozása, csoportosítása, megjelenítése - adatok harmonizálása Adatbevitel, adatrendezés adatbankok Adatfeldolgozás, adatmegjelenítés, kiértékelés újabb információk újabb adatbankok Adatbankok: - adatok gyors cseréje - interaktív kapcsolat az és a kutatók között adatbankok automatizálás, speciális szoftverek speciális szaktudás

Pre-bioinformatika, az informácó hordozó megfejtése 1866 Mendel: borsó keresztezési kísérlek 1869 Miescher: lazac sperma DNS tisztítás öröklődés egységekben DNS az örökítő anyag 1903 WS Sutton az öröklődési mintázat a kromoszóma sajátságaihoz kapcsolt az osztódás során citokémia: a kromoszóma DNS-ből és fehérjéből épül fel

Pre-bioinformatika, az informácó hordozó megfejtése F. Griffith 1925 -1928 Avery 1944 Streptococcus pneumoniae egér meghal virulens baktérium egér túlél nem-virulens baktérium egér túlél hőkezelt baktérium a transzformáló anyag DNS proteáz RNáz DNáz nincs hatás inaktivált egér meghal nem-virulens baktérium proteáz szennyeződés? + hőkezelt baktérium virulens baktérium transzformálási elv

Hershey és Chase 1952 T 2 fág DNS fehérje burok DNS-ben nincs S, fehérjében nincs P 32 Pjelölt 35 S jelölt fág DNS fágok a baktériumhoz tapadva bakteriofág fehérje burok fág DNS turmixolás fehérje burok leválik új fágok képződnek baktérium lizál a fágok kiszabadulnak 70% 32 P 20% 35 S

Út a kettős hélixhez, Crick és Chargaff E. nukleotid arányok humán sejt E. coli baktérium 1952 -1953 Biofizikai adatok, víztartalom Pauling triple hélix DNS tisztítás gyenge savas kezelés foszfodiészter kötés hidrolízis Röntgen diffrakciós adatok Rosalind Franklin és Maurice Wilkins kromatográfia és a nukleotidok kvantitálása Bázis arány Watson Bázis arány A: T 1. 00 A: T 1. 09 G: C 1. 00 G: C 0, 99 fehérje alfa hélix már ismert

A DNS kettős hélix

Centrális dogma és a bioinformatika főbb területei a molekuláris biológiában Gén DNS transzkripció, RNS szerkesztés transzkriptomika RNS degradáció transzláció, poszttranszlációs módosítás fehérje proteomika degradáció biokémiai aktivitás metabolikus útvonalak metabolomika

A BIOLÓGIAI INFORMÁCIÓ HORDOZÓ MEGFEJTÉSE GENOMIKA A teljes genetikai állomány szekvenciájának meghatározása, A szekvenciákon elhelyezkedő funkcionális régiók számítógépes jóslása: annotálás 9

Funkcionális genomika RNS szinten TRANSZKIPTOMIKA 10

Egy DNS chip kísérlet folyamatábrája

A chipek kiértékelése, eredménye 12

Funkcionális genomika fehérje szinten PROTEOMIKA 13

Proteomika Egy. Tipikus protokol Izoelektromos fókuszálás SDS PAGE Minta elő Protein azonosítás tömegspektrometria Láthatóvá tétel Protein pötty kivágás Kép analízis

15

Proteomika: az elválasztástól az azonosításig 16
 Approximate number of genes References Arabidopsis thaliana (plant) 125")
Species Size of genome (Mb) Approximate number of genes References Arabidopsis thaliana (plant) 125 25 500 AGI (2000) Caenorhabditis elegans (nematode worm) 97 19 000 CESC (1998) Drosophila melanogaster (fruit fly) 180 13 600 Adams et al. (2000) Homo sapiens (human) 3200 40 000 IHGSC (2001); Venter et al. (2001) Saccharomyces cerevisiae (yeast) 12. 1 5800 Goffeau et al. (1996) Escherichia coli K 12 4. 64 4400 Blattner et al. (1997) Mycobacterium tuberculosis H 37 Rv 4. 41 4000 Cole et al. (1998) Mycoplasma genitalium 0. 58 500 Fraser et al. (1995) Pseudomonas aeruginosa PA 01 6. 26 5700 Stover et al. (2000) Streptococcus pneumoniae 2. 16 2300 Tettelin et al. (2001) Vibrio cholerae El Tor N 16961 4. 03 4000 Heidelberg et al. (2000) Yersinia pestis CO 92 4. 65 4100 Parkhill et al. (2001) Archaeoglobus fulgidus 2. 18 2500 Klenk et al. (1997) Methanococcus jannaschii 1. 66 1750 Bult et al. (1996) Eukarióta Eukaryotes Bacteria Archaeacteria

A BIOLÓGIAI INFORMÁCIÓ HORDOZÓ MEGFEJTÉSE GENOMIKA A teljes genetikai állomány szekvenciájának meghatározása, A szekvenciákon elhelyezkedő funkcionális régiók számítógépes jóslása: annotálás 18

Az emberi genetikai állomány emberi család emberi sejtmag genom mitokondriális genom 22 autoszóma 2 szex kromoszóma

A humán és élesztő mitokondriális genom respirációs komplex génjei riboszómális RNS gének intronok riboszómális protein gének egyéb RNS gén transfer RNS gén

Másik extrakromoszómális elem növényekben: kloroplaszt A rizs kloroplasztjának genomja 136 kb fotoszintézis gének riboszómális protein gének riboszómális RNS gének transzfer RNS gének RNS polimeráz gén
 Mitochondrial genomes Plasmodium falciparum Protozoan (malaria parasite)")
Species Type of organism Genome size (kb) Mitochondrial genomes Plasmodium falciparum Protozoan (malaria parasite) 6 Chlamydomonas reinhardtii Green alga 16 Mus musculus Vertebrate (mouse) 16 Homo sapiens Vertebrate (human) 17 Metridium senile Invertebrate (sea anemone) 17 Drosophila melanogaster Invertebrate (fruit fly) 19 Chondrus crispus Red alga 26 Aspergillus nidulans Ascomycete fungus 33 Reclinomonas americana Protozoa 69 Saccharomyces cerevisiae Yeast 75 Suillus grisellus Basidiomycete fungus 121 Brassica oleracea Flowering plant (cabbage) 160 Arabidopsis thaliana Flowering plant (vetch) 367 Zea mays Flowering plant (maize) 570 Cucumis melo Flowering plant (melon) 2500 Chloroplast genomes Pisum sativum Flowering plant (pea) 120 Marchantia polymorpha Liverwort 121 Oryza sativa Flowering plant (rice) 136 Nicotiana tabacum Flowering plant (tobacco) 156 Chlamydomonas reinhardtii Green alga 195
 kezdete")
Eukarióta gének szerkezete exon upstream intron exon szabályozó elemek biológiai információ (kódoló régió) kezdete downstream biológiai információ (kódoló régió) vége altenatív splicing egymásba ágyazott gének neurofibromatosis type I gene exons introns OGMP EVI 2 B EVI 2 A

“Abnormális” genetikai elemek Pszeudogének keletkezése A. Processzált pszeudogén B. funkcionális gén transzkripció reverz transzkripció RNS DNS funkcionális gén új integráció csonka gén pszeudogén génfragment a kódoló régió is sérült nincs szabályozó régió konvencionális pszeudogén: funkcióvesztéses mutáció
 13 bp repeat,")
ISMÉTLŐDŐ SZEKVENCIÁK A GENOMOKBAN 1 kromoszóma mikroszatellitek (short tandem repeat, STR) 13 bp repeat, interspersed repeats 150 bp hossz: pl. CACACA 2 kromoszóma átlagosan minden 2 kb tartalmaz miniszatellitek tandem repeated DNA Long Interspersed Nuclear Elements: LINE Short Interspersed Nuclear Elements: SINE 25 bp repeat, 20 kbp hossz Genetikai profil analízisére alkalmasak

Retroelemek és retrotranszpozíció retrotransposon transzkripció reverz transzkripció RNS DNS retrotransposon új integráció retrotransposon kópia

DNS transpozonok replikatív konzervatív eukariótákban a retrotranszpozon a jellemzőbb

A HUMÁN GENOM EGY SZEGMENSE

A prokarióta genom szerkezete cirkuláris kétszálú DNS Az E. coli nucleoidjának modell szerkezete néhány fordulat kettős hélix megbomlása negatív szupertekeredett struktúra

Prokarióta gének felépítése, policisztronos struktúra

A laterális géntranszfer szerepe különböző porkariótákban

Evolúciós törzsfa Archaeák: Carl Woese: - 1977. - 16 S r. RNS szekvenciák univerzális filogenetikai törzsfa Archaebaktériumok: - Eubaktérium-szerű tulajdonságok: - sejtszerveződés - sejtciklus - fő metabolikus utak - cirkuláris kromoszóma, replikáció - policisztronos operonok - Shine- Dalgarno szekvenciák (SD) - transzkripció és transzláció öszekapcsolt - génexpresszió szabályozás (regulátor fehérjék) Eukarióta-szerű tulajdonságok: transzkripció, transzláció: - promóter elemek: TATA-box (-30) - transzkripciós faktorok: TBP és TFB - RNS polimeráz sok alegységes (~12) - riboszómák: 70 S 16 S, 23 S, 5 S de: eukariótákéhoz hasonló riboszómális fehérjék - transzlációs faktorok - intronok, - kis nukleoláris RNS szerű molekulák (sno. RNS) - hiszton fehérjék (erősen bázikusak) nukleoszómák (kis árok) - hősokk fehérjék (Hsp 60) - citoplazmában chaperonok

DNS MANIPULÁCIÓ számítógéppel

Clone Manager 6
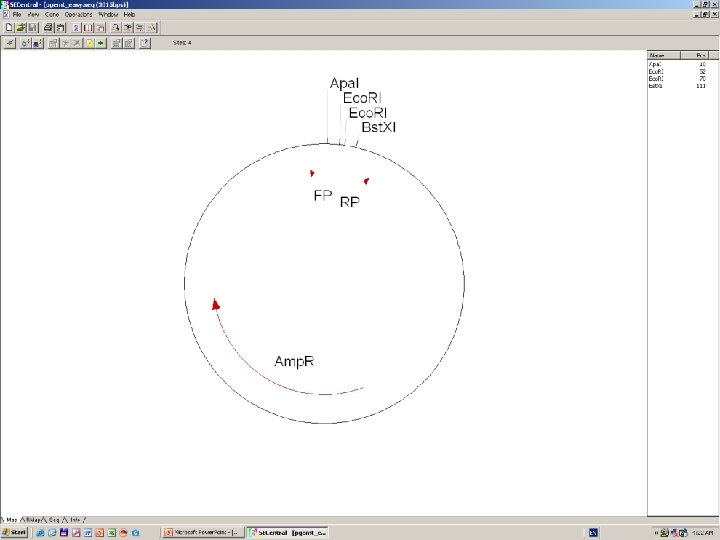
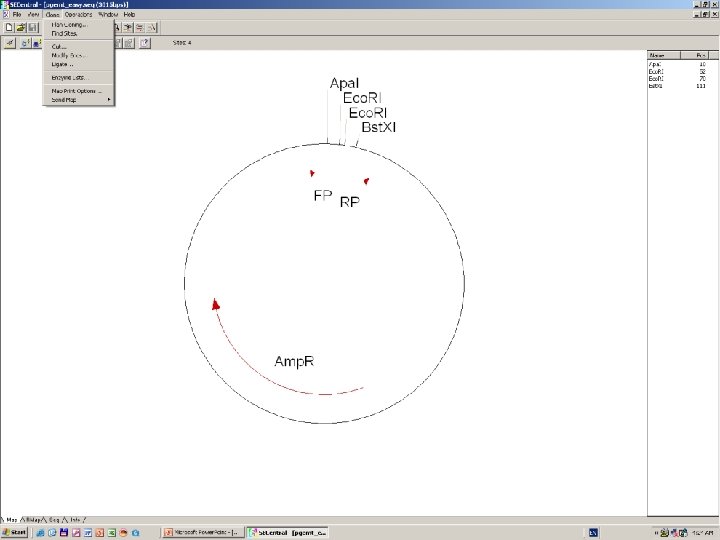
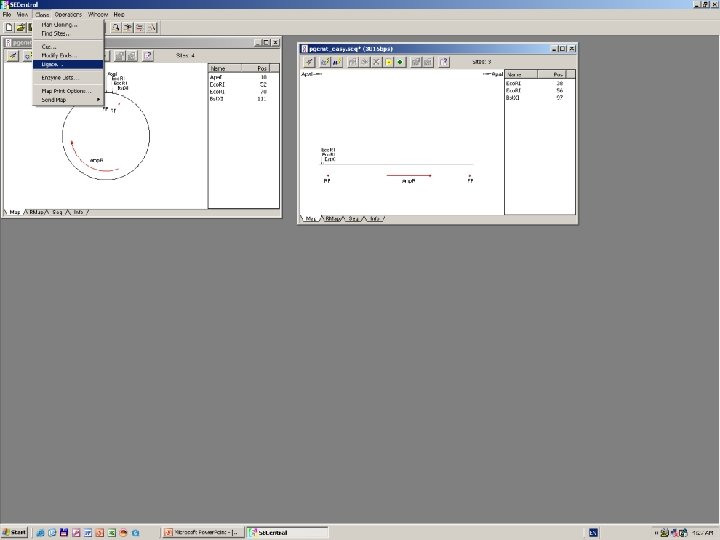
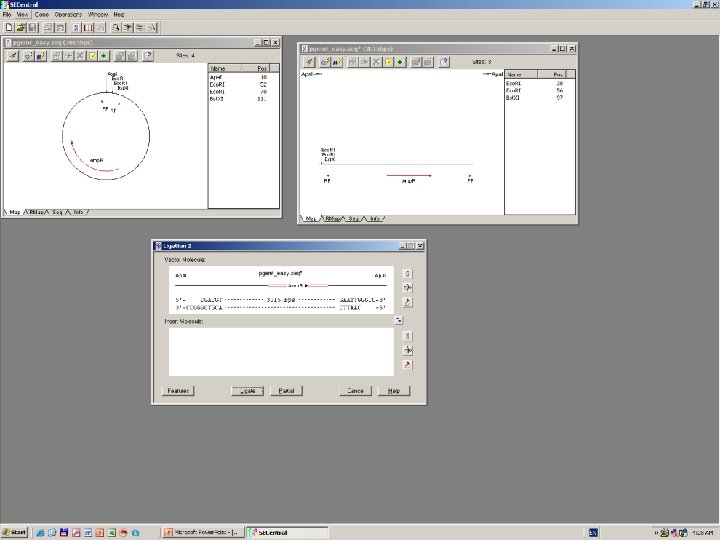
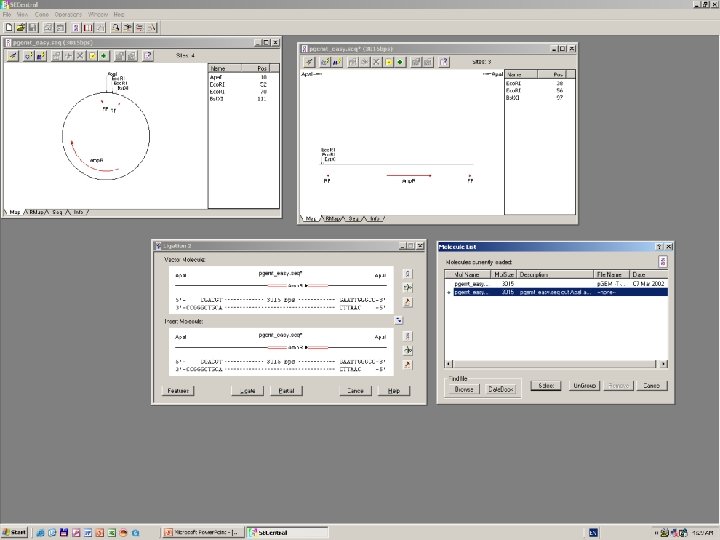
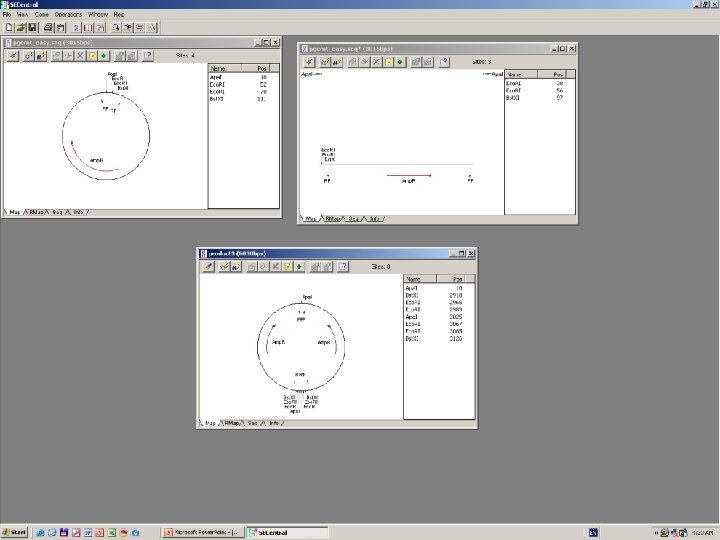

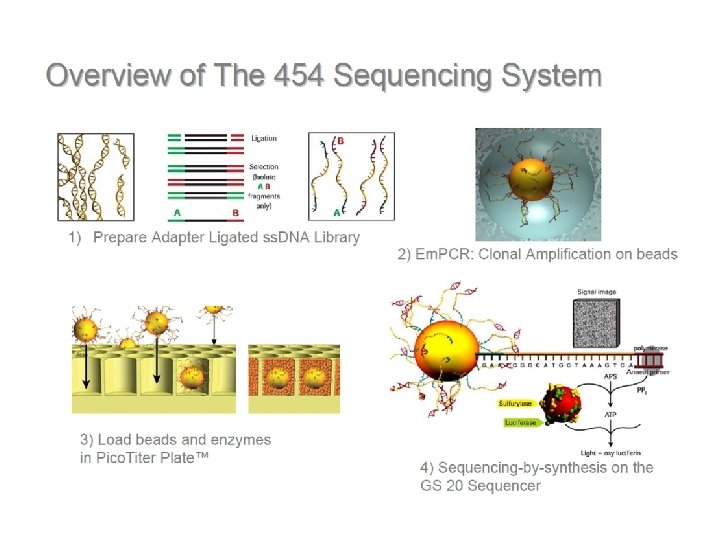
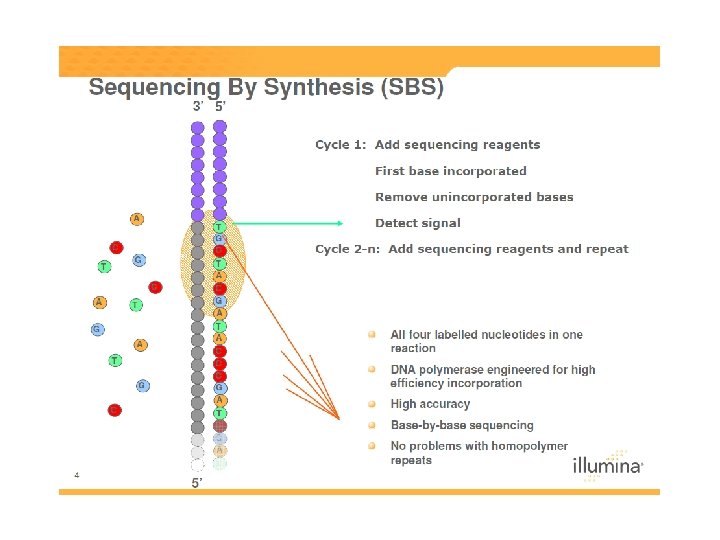
DNS szekvenálás SANGER szerint +

KLASSZIKUS DNS SZEKVENÁLÁS PCR TERMÉKEN VAGY KLÓNOZÓ VEKTORBAN

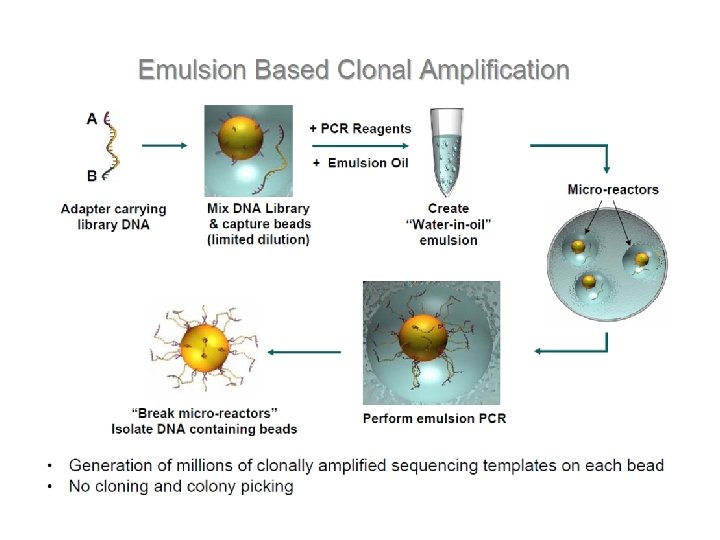
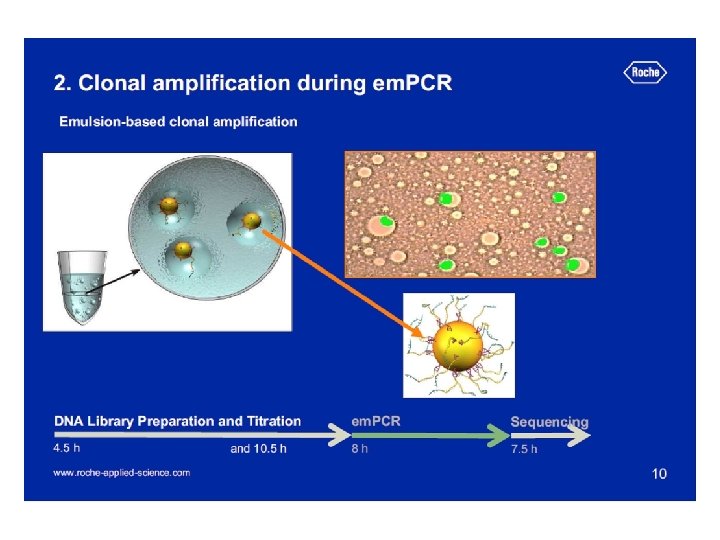
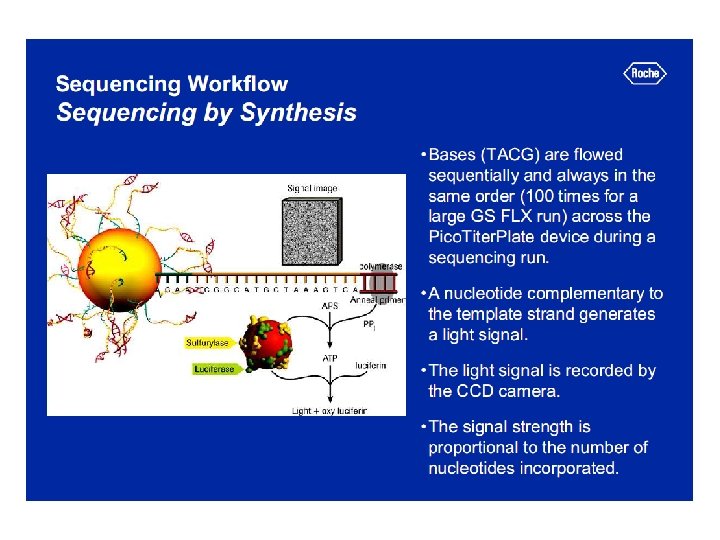
Szekvenálási stratégiák Automata, Sanger-alapú piroszekvenálás Chip-alapú

A PCR ALAPÚ SZEKVENÁLÁS SÉMÁJA TEMPLÁT DNS PCR egy primerrel a terminálódott láncok száma a ciklus számmal növekszik a hiba nem amplifikálódik

AZ AUTOMATA DNS SZEKVENÁLÁS ELVE

AZ AUTOMATA SZEKVÁNÁLÓ A SZEKVENCIÁK MANUÁLIS ELLENŐRZÉSE

Genom szekvenálási stratégiák Shot gun Primer séta

Alternatív shot gun stratégiák térképezés: - genetikai: gének, tulajdonságok pozícionálása - fizikai: szekvenciák, gének rendeződése

Bakteriális shot gun könyvtár készítése

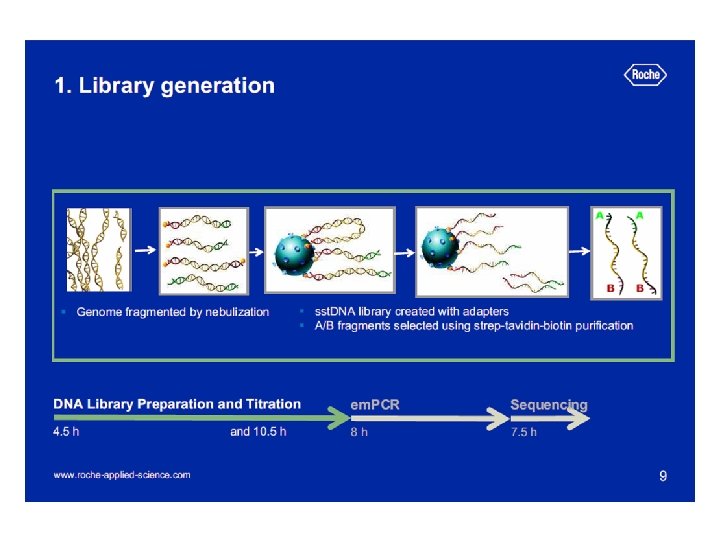
Preparation of shotgun library E. coli chromosomal DNA transformation electroporation 2 -3, 5 kb fragments blunting the ends broken DNA fragments dephosphorylation Preparative gel electrophoresis

A szekvenciák feldolgozása szekvenciaanalízis ellenőrzés, validáció Phrap Seq. Man/DNASTAR STADEN programcsomag vektor és egyéb szennyező szekvenciák eltávolítása gyenge minőségű szekvenciák eltávolítása Vector_clipping Phrap átfedő fragmentumokból kontigok összerakása Phred

Szekvenciák kontigokba rendezése, Példa S 19 T 7 S 12 SK S 19 SK S 148 O 20 S 11 T 7 S 17 SK S 148 O 22 S 148 O 15 S 148019 S 148 O 13 S 148 O 8 S 17 T 7 S 148 O 17 S 13 SK S 148 O 14 S 148 O 18 S 12 T 7 S 13 T 7 S 148 O 21 mac. A orf 2 pca. B 2000 p. SC 1/1 S 148 SK S 148 O 11 S 148 O 9 orf 1 S 148 O 12 S 148 O 10 S 11 SK S 148 T 7 SC 110 SK SC 110 T 7 orf-3 p. SC 1/2 PSC 148 6000 p. SC 1/3 (7405 bps) pca. G pca. H 4000 S 14 SK S 18 SK S 16 SK p. SC 1/8 p. SC 1/10 p. SC 1/4 p. SC 1/6









File management

Szekvenciák, kontiggá való összerakása I. Tévedések vígjátéka

A scaffold fogalma Szekvenálási lyukak Kontig 1 Kontig 2 Scaffold 1 Nincs kapcsolat Kontig 3 Kontig 4 Kontig 5 Scaffold 2 összekapcsolás: kozmidkönyvtár (BAC, YAC) klónok végeinek szekvenciái két küldönböző kontigra esnek Scaffold: láncszerűen lineáris sorrendbe elhelyezett nem összeérő kontigok sora.

Kontigok szuperkontigokká való összerakása
 Buzgó (greedy) algoritmus Átfedési-szerkezet-konszenzus")
Összerakó algoritmusok (Assemblers) Buzgó (greedy) algoritmus Átfedési-szerkezet-konszenzus

De ha sikerül, és van szekvenciánk Mi van rajta, van-e gén? Honnan tudjuk, hogy Valamit találtunk, találtunk-e gént? CTCGAGACGCTGTTTCTGGGGTCATTCTTGGCGGGCTGCAACTGCTGGTGTGACCGACGCGACCTGGCAGGCCGCGGTGCGCAACTGGCCGGGCGGACTAATGGTGGAGCAAAAGA TCGGCATGTCCAGCGCACCTGAAGCTTGGGTGGTTGCTGCAATAGCAGCCTTATTGGCATGGCGAAGGGCGGTTTGGCCAATGTGGGGGTTATCGCCGTTCCCTTGATGTCCCTGG TCAAGCCGCCGCTTACCGCTGCCGGATTGCTGCTCCCGATCTATGTCGTTTCTGATGCATTCGGCGTCTGGCTTTATCGGCACCGGTATTCTGCCTCCAATCTGCGCATCCTGATTCCTT CGGGATTTTTTGGGGTCCTGATTGGCTGGTTATTGGCCGGGCAGATCTCCGACGCGATTGCCAGTGTCATTGTTGGTTTCACCGGCTGCGGCTTCGTGGCTGTGCTGCTGGCACGACGAG GGGTGCCATCGGTGCCGCGTCAAGCCAACGTGCCCAAAGGATGGTTTCTGGGGGTGGCCACCGGCTTTACCAGCTTTTTGACTCATTCCGGTGCGGCGACCTTCCAGATGTTCGTGCTGC CGCAACGGCTGGACAAGACCATGTTCGCGGGCACATCAACGCTTACCTTTGCTGCCATAAACCTATTCAAGATTCCGTCCTACTGGGCATTGGGACAGCTTTCGACTTCCTCGGTCATGT CCGCGCTAGTGTTGATTCCGGTGGCCGGGACGTTCGCAGGTGTTTTTGCGACGCGCAGGCTATCGACATCCTGGTTCTTCATTCTGGTCCAGGCGATGTTGCTGGTGGTCTCCA TTCAGCTTCTGTGGAGGGGAATGTCGGATATCCTGAACTAGCTGGAGATCGCAATGTCAGAACGCTCAATCAGAATGTAATCTTGACATAGAATACCGTTCCGATTTATTGCTTCG AGTGAAGCTGCCCGTCCGCTGAGATGTCATGACATTTTCCCCGCTTGATTCCGCCCTGCTTGGACCGTTGTTCGCGACCGATGAAATGCGCACGGTCTTCTCCGAACGGCGTTTTTTGGC GGGAATGCTTCGTGTTGAAGTGGCCCTGGCGCGCAGGCGGCAGAGGGCCTTGTCAGTTCGGAATTGGCCGACGCGATCGAGGTTGTTGGTACTGCCGGGTTGGACCCCGAGGCGAT GGCGGCGACTACTCGCATGACAGGAGTGCCCGCAATATCGTCCGTGCGGTGCAATCGGCCCTGCCGCCCTCACTGGCGGGTGGATTTCGGCGCCACCAGTCAAGACATCGT GGATACGGCCCACGCGCTCCAGCTGGCCGAGGCACTCGATATTATAGAAGTCGATTTACACGCCACTGTCAGCGCAATGATGAATCTGGCCGCTGCTCACTGCAATACACCCTGTATCGG GCGCACGGCCTTGCAGCACGCAGCGCCAGTTACGTTCGGCTACAAGGCGTCCGGCTGGTGCGTTGCCCTGGCGGAGCATCTGGTGCAGCTTCCCGCGCTGCGAAAGCGGGTTCTGGTGGC GTCGCTAGGGGGGCCGGTTGGTACCCTTGCCGCGATGGAGGAGCGGGCCGACGCTGTACTGGAGGGTTTCGCTGCGGACCTGGGGTTGGCCATTCCCGCCCTGGCACACGCAGCG GGCCCGGATCGTCGAGGTGGCCAGTTGGCCATATTGCTGGGAATTCTGGCAAAAATGGCCACCGATGTCGTTCACTTGTCCTCCACGGAAGTGCGCGAGCTTTCCGAACCTGTAGC GCCGGGCAGGGGGGGCTCCTCGGCGATGCCTCACAAGCGGAACCCGATTTCCTCGATTACCATCCTGTCCCAGCATGCTGCGGCAGGGGCCCAGCTCTCCATTCTCGTGAACGGCATGGC CAGTCTGCACGAACGTCCGGTGGGGGCGTGGCATTCGGAATGGTTGGCTCTGCCGACGCTGTTCGGCCTTGCCGGCGGTGCCGTGCGCGAGGGCAGGTTTCTGGCCGAGGGGCTGCTGGT CGATGCCGACCAGATGGGTCGCAATCTACAATTGACCAATGGCCTGATTTTCAGCGACGCGGTAGCCGGCCAGTTGGCAAAGCACTTGGGTCGGGCCGAGGCTTATGCCGCTGTCGAGGA TGCCGCCGCCGAGGTGTTGCGTTCAGGCGGCAGCTTTCAGGGTCAGCTGAACCAGCGCCTGCCCGATCACCGCGACGCTATTGCTTTTGATACGACGCCGGCGATCCAGGCCGG GGCCGCTGCCGTAGTGCGCTGGATCATGTGGCTCGTATTCTTGGACCCGCCTCTACCATCGGATTTCAAGGAGGCTAATGACGACACTGTTTGAGGCGACGACCATCCCGA TTTGCGAGGGCCCGCGCGACCAGACCGCCGAGATCCTTTTCGAGATGCCGCCGGGTGCGTGGGATACCCATTTTCATGTTTTTGGCCCAGTTTCATCGTTTCCATACGCAGAACACAGGC TCTATTCCCCACCGGAGTCGCCACTTGAGGATTATCTGGTGTTGATGGAGGCTTTGGGGATCGAGCGCGGCGTTTGTGTCCATCCGAATGTTCATGGTGCCGACAATTCGGTGACGCTCG ACGCAGTTGCGCGGTCCGATGGTCGTCTGCTGGCGGTGATCAAGCCACATCACGAGATGACTTTTGTTCAGCTGCGGGACATGAAGGCGCAGGGGGTCTGCGGGGTACGTTTTGCCTTCA ATCCGCAGCATGGCTCGGGCGAGTTGGATACTCGTTTGTTCGAGCGTATGTTGGACTGGTGCCGCGACCTAGGCTGGTGCGTAAAATTGCATTTCGCGCCCGCTGGACGGTCTGG CTGAACGTTTGGCGCGCGTCGATATTCCGATCATCATCGATCATTTCGGGTGGACACCGCGCAAGGTGTGGATCAGCCGCACTTCCTGCGTTTGCTCGATCTGGCCAAACTGGACC

Hasonlóság CTCGAGACGCTGTTTCTGGGGTCATTCATTCTTGGCGGGC TGCAACTGCTGGTGTGACCGACGCGACCTGGCAGGCCGCG GTGCGCAACTGGCCGGGCGGACTAATGGTGGAGCAAAAG ATCGGCATGTCCAGCGCACCTGAAGCTTGGGTGGTTGCTG CAATAGCAGCCTTCCTTATTGGCATGGCGAAGGGCGGTT TGGCCAATGTGGGGGTTATCGCCGTTCCCTTGATGTCCCT GGTCAAGCCGCCGCTTACCGCTGCCGGATTGCTGCTCCCG ATCTATGTCGTTTCTGATGCATTCGGCGTCTGGCTTTATC GGCACCGGTATTCTGCCTCCAATCTGCGCATCCTGATTCC TTCGGGATTTTTTGGGGTCCTGATTGGCTGGTTATTGGCC GGGCAGATCTCCGACGCGATTGCCAGTGTCATTGTTGGTT TCACCGGCTGCGGCTTCGTGGCTGTGCTGCTGGCACGACG AGGGGTGCCATCGGTGCCGCGTCAAGCCAACGTGCCCAAA GGATGGTTTCTGGGGGTGGCCACCGGCTTTACCAGCTTTT TGACTCATTCCGGTGCGGCGACCTTCCAGATGTTCGTGCT GCCGCAACGGCTGGACAAGACCATGTTCGCGGGCACATCA ACGCTTACCTTTGCTGCCATAAACCTATTCAAGATTCCG TCCTACTGGGCATTGGGACAGCTTTCGACTTCCTCGGTCA TGTCCGCGCTAGTGTTGATTCCGGTGGCCGTGGCCGGGAC GTTCGCAGGTGTTTTTGCGACGCGCAGGCTATCGACATCC TGGTTCTTCATTCTGGTCCAGGCGATGTTGCTGGTGGTCT CCATTCAGCTTCTGTGGAGGGGAATGTCGGATATCCTGA ACTAGCTGGAGATCGCAATGTCAGAACGCTCAATCAATC AGAATGTAATCTTGACATAGAATACCGTTCCGATTTATT GCTTCGAGTGAAGCTGCCCGTCCGCTGAGATGTCATGACA TTTTCCCCGCTTGATTCCGCCCTGCTTGGACCGTTGTTCGC GACCGATGAAATGCGCACGGTCTTCTCCGAACGGCGTTTT TTGGC a két szekvencia teljesen ugyanaz

Hasonlóság Most is - majdnem CTCGAGACGCTGTTTCTGGGGTCATTCTTGGCGGGC TGCAACTGCTGGTGTGACCGACGCGACCTGGCAGGCCGCG GTGCGCAACTGGCCGGGCGGACTAATGGTGGAGCAAAAG ATCGGCATGTCCAGCGCACCTGAAGCTTGGGTGGTTGCTG CAATAGCAGCCTTATTGGCATGGCGAAGGGCGGTT TGGCCAATGTGGGGGTTATCGCCGTTCCCTTGATGTCCCT GGTCAAGCCGCCGCTTACCGCTGCCGGATTGCTGCTCCCG ATCTATGTCGTTTCTGATGCATTCGGCGTCTGGCTTTATC GGCACCGGTATTCTGCCTCCAATCTGCGCATCCTGATTCC TTCGGGATTTTTTGGGGTCCTGATTGGCTGGTTATTGGCC GGGCAGATCTCCGACGCGATTGCCAGTGTCATTGTTGGTT TCACCGGCTGCGGCTTCGTGGCTGTGCTGCTGGCACGACG AGGGGTGCCATCGGTGCCGCGTCAAGCCAACGTGCCCAAA GGATGGTTTCTGGGGGTGGCCACCGGCTTTACCAGCTTTT TGACTCATTCCGGTGCGGCGACCTTCCAGATGTTCGTGCT GCCGCAACGGCTGGACAAGACCATGTTCGCGGGCACATCA ACGCTTACCTTTGCTGCCATAAACCTATTCAAGATTCCG TCCTACTGGGCATTGGGACAGCTTTCGACTTCCTCGGTCA TGTCCGCGCTAGTGTTGATTCCGGTGGCCGGGAC GTTCGCAGGTGTTTTTGCGACGCGCAGGCTATCGACATCC TGGTTCTTCATTCTGGTCCAGGCGATGTTGCTGGTGGTCT CCATTCAGCTTCTGTGGAGGGGAATGTCGGATATCCTGA ACTAGCTGGAGATCGCAATGTCAGAACGCTCAATC AGAATGTAATCTTGACATAGAATACCGTTCCGATTTATT GCTTCGAGTGAAGCTGCCCGTCCGCTGAGATGTCATGACA TTTTCCCCGCTTGATTCCGCCCTGCTTGGACCGTTGTTCGC GACCGATGAAATGCGCACGGTCTTCTCCGAACGGCGTTTT TTGGC GLOBÁLIS, LOKÁLIS AAACTCGAGACGCTGTTTCTGGGGTCATTCTTGGC GGGCTGCAACTGCTGGTGTGACCGACGCGACCTGGCAGGC CGCGGTGCGCAACTGGCCGGGCGGACTAATGGTGGAGCA AAAGATCGGCATGTCCAGCGCACCTGAAGCTTGGGTGGT TGCTGCAATAGCAGCCTTATTGGCATGGCGAAGGG CGGTTTGGCCAATGTGGGGGTTATCGCCGTTCCCTTGATG TCCCTGGTCAAGCCGCCGCTTACCGCTGCCGGATTGCTGC TCCCGATCTATGTCGTTTCTGATGCATTCGGCGTCTGGCT TTATCGGCACCGGTATTCTGCCTCCAATCTGCGCATCCTG ATTCCTTCGGGATTTTTTGGGGTCCTGATTGGCTGGTTA TTGGCCGGGCAGATCTCCGACGCGATTGCCAGTGTCATTG TTGGTTTCACCGGCTGCGGCTTCGTGGCTGTGCTGCTGGC ACGACGAGGGGTGCCATCGGTGCCGCGTCAAGCCAACGTG CCCAAAGGATGGTTTCTGGGGGTGGCCACCGGCTTTACCA GCTTTTTGACTCATTCCGGTGCGGCGACCTTCCAGATGTT CGTGCTGCCGCAACGGCTGGACAAGACCATGTTCGCGGGC ACATCAACGCTTACCTTTGCTGCCATAAACCTATTCAAG ATTCCGTCCTACTGGGCATTGGGACAGCTTTCGACTTCCT CGGTCATGTCCGCGCTAGTGTTGATTCCGGTGGCCGTGGC CGGGACGTTCGCAGGTGTTTTTGCGACGCGCAGGCTATCG ACATCCTGGTTCTTCATTCTGGTCCAGGCGATGTTGCTGG TGGTCTCCATTCAGCTTCTGTGGAGGGGAATGTCGGATA TCCTGAACTAGCTGGAGATCGCAATGTCAGAACGCTCAATCAGAATGTAATCTTGACATAGAATACCGTTCCGA TTTATTGCTTCGAGTGAAGCTGCCCGTCCGCTGAGATGTC ATGACATTTTCCCCGCTTGATTCCGCCCTGCTTGGACCGT TGTTCGCGACCGATGAAATGCGCACGGTCTTCTCCGAACG GCGTTTTTTGGC BLASTN, FASTA

ANALÓGIÁK - ADATBANKOK Összahasonlítás már ismert elemekkel

… és kódol-e fehérjét? Open reading frames: nyitott leolvasási keretek Áltában ATG-vel kezdődik, de opció Hossz: ajánlás 100 aminosav, de opció Az eredmény hipotetikus, össze kell vetni a valósággal Hipotetikus fehérje lista hasonlóság BLASTP Információból információ generálása Problémák: frameshift mutáció, a globál hasonlóság csődje Hol kezdődik? Mi a start?

Egy régió nyitott leolvasási keret térképe

Hol kezdődik? Ki tudja? Egyéb elemek azonosítása, genomi elrendeződés Kísérletes ellenőrzés

FRAME SHIFT MUTÁCIÓ - MEGOLDÁS Minden leolvasásái keretben transzláció Stop kodon nem számít Mindent mindennel összehasonlít fehérje szinten BLASTX

In silico transzláció mind a 6 leolvasási keretben

Kodon felhasználás, codon usage Az élőlényekre jellemző a kodon felhasználási gyakoriság Kodon felhasználási táblázatok, adatbankok

Kodon felhasználás, codon usage

Kodon felhasználás, eltérések
2fb55a413edc35f9abb6c940741a9e92.ppt