Презентация по биофизике для ДИТИ МИФИ Ч3.ppt
- Количество слайдов: 166



БИОФИЗИКА МЕМБРАННЫХ ПРОЦЕССОВ клетка

Клетка - открытая система Элементарная живая система: Самостоятельное существование Развитие Размножение Автономна по отношению к окружающей среде Взаимодействует с окружающей средой

Схематическое изображение клетки животного и растения

БМ в клетке Плазматическая мембрана - образует границу клетки Органеллы • Ядро • Эндоплазматический ретикулум • Аппарат Гольджи • Лизосомы • Митохондрии • Хлоропласты • …. •
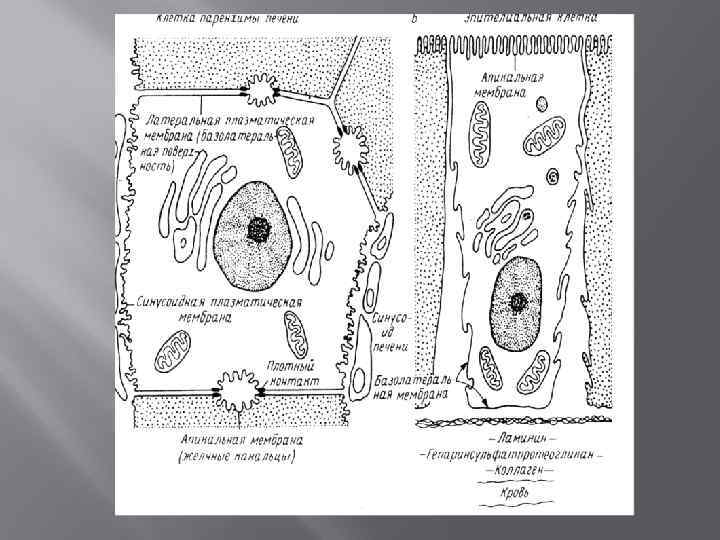

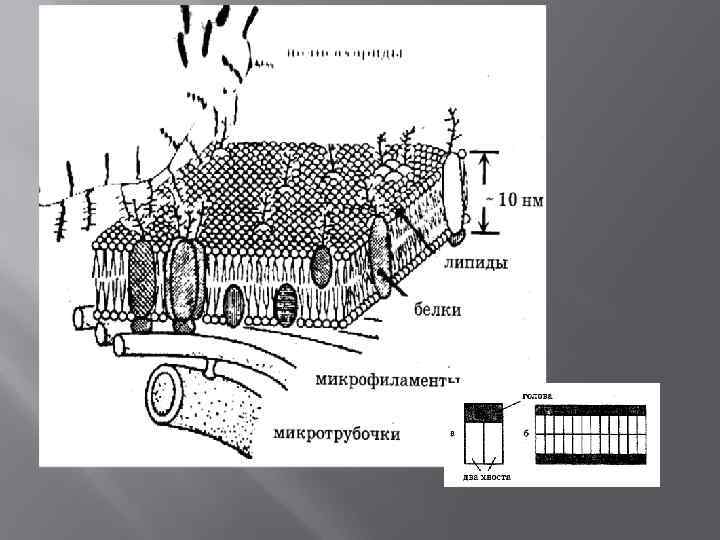
Основные функции БМ Барьерная – селективный, регулируемый пассивный транспорт, активный обмен веществом с окружающей средой Матричная – определенное взаимное расположение и ориентация мембранных белков Механическая – прочность и автономность клетки, внутриклеточных структур

Дополнительные функции БМ Энергетическая – синтез АТФ и фотосинтез Генерация и проведение потенциалов Рецепторная – механическая, акустическая, обонятельная, зрительная, химическая, терморецепция ….

Развитие представлений о строении биологических мембран XIX в – ПМ - определенная структура к XIX в – Овертон: БМ состоят из молекул, которые похожи на молекулы масла (липиды) 1925 г – Гардер и Грендел БМ образована 2 -ым слоем липидных молекул (липидный бислой) кювета Ленгмюра

Развитие представлений о строении биологических мембран 1935 г Коул и Кертис – подтвердили гипотезу БМ двойной липидный слой электрическое сопротивление 107 Ом▪м 2 удельная электрическая емкость 0, 5*10 -2 Ф/м 2 БМ – электрический конденсатор Толщина неполярной части

Развитие представлений о строении биологических мембран 1935 г Дэвинсон и Даниелли – модель БМ «сендвича» • Рентегоструктурный анализ – упорядоченность в расположении липидных молекул в БМ • Электронная микроскопия – в БМ встроены глобулярные частицы • 1972 г – Спингер и Николсон – жидко-мозаичная модель БМ: БМ текучий фосфолипидный слой, в который погружены свободно диффундирующие белки •
")
Развитие представлений о молекулярной организации БМ (по Ю. А. Владимирову)

Образование мембранных структур • Основные молекулярные компоненты БМ – биополимеры, функционирующие в водной среде
 Электронная микроскопия а) оптический микроскоп: отдельные части клетки")
Методы изучения БМ (состав, структура, строение) Электронная микроскопия а) оптический микроскоп: отдельные части клетки б) электронный микроскоп внутреннее строение, клеточные органеллы, детали строения БМ «Замораживание-скол-травление» • ЭПР, ЯМР, флуоресцентные методы: динамические характеристики БМ •

ЭПР Различия в спектрах ЭПР в зависимости от способа прикрепления спиновой метки к фосфолипидной молекуле
")
Изменение спектров ЭПР при увеличении подвижности (уменьшении микровязкости)

БМ Функциональные структуры клетки, которые ограничивают цитоплазму и большинство внутриклеточных структур, образуют единую систему канальцев, складок и замкнутых полостей Толщина 10 нм масса 1/2 массы сухой клетки Состав: липиды, белки, углеводы
 Связанная вода • а) внутренняя связанная вода • б) вода")
Вода в БМ 1) Связанная вода • а) внутренняя связанная вода • б) вода гитратных оболочек • в) слабосвязанная вода 2) Свободная вода 3) Захваченная вода

Физическое состояние липидных мембран Газ Жидкость Твердое тело Плазма БМ – жидкокристаллическое агрегатное состояние

Жидко - кристаллические структуры

Фазовые переходы в липидных мембранах
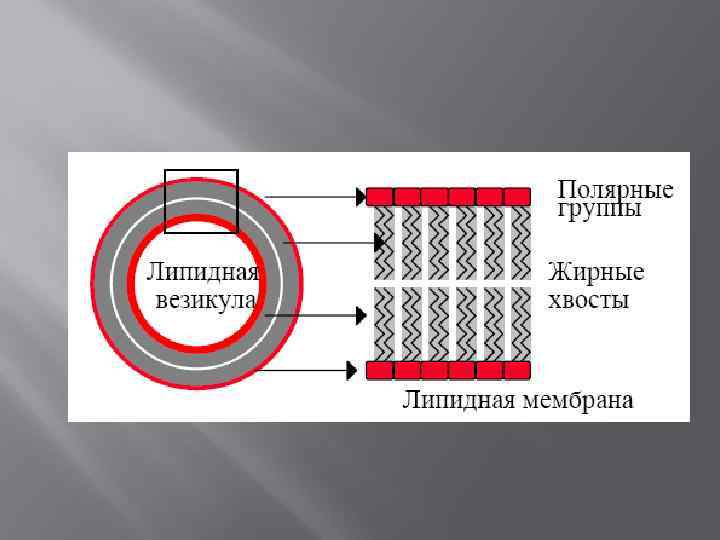

Модельные липидные мембраны Схема строения однослойной липосомы

Модельные липидные мембраны Образование плоской бислойной липидной мембраны

ФИЗИЧЕСКИЕ СВОЙСТВА МЕМБРАН

Подвижность молекулярных компонентов в мембранах Вращательная подвижность. Время поворота липида вокруг своей оси на 1 рад 10 -9 с Латеральная диффузия - перемещение молекул липидов вдоль слоя. Коэффициент латеральной диффузии D 10 -7 - 10 -8 см 2/с Среднеквадратичное расстояние

Передвижение иона в липидном слое мембран

Подвижность молекулярных компонентов в мембранах

Подвижность молекулярных компонентов в мембранах Частота перескоков Площадь, занимаемая одной молекулой f Характерное время перескока

ЭПР Различия в спектрах ЭПР в зависимости от способа прикрепления спиновой метки к фосфолипидной молекуле (подвижность)

Трансбислойное движение: флип-флоп-переход Время движения в БМ: • -БМ электрического органа угря 3 -7 мин • -БМ эритроцитов 20 -30 мин • Модельные визикулярные мембраны 10 - 20 ч и более

Константы латеральной диффузии мембранных белков

Микровязкость углеводородной области липидного бислоя в искусственных и природных мембранах, маслах и жидкостях
 Свободная энергия Гельмгольца F=U-TS (d. F)T=d. U-Td.")
Упругие свойства мембран В равновесном состоянии (T=const) Свободная энергия Гельмгольца F=U-TS (d. F)T=d. U-Td. S=d. W [F]=[W]=Дж/м 2=Н/м Деформации - относительное изменение площади: растяжение (сжатие) = S|S - Растяжение (сдвиг) поверхности

• Изотропное растяжение поверхности БМ относительно равновесного состояния, натяжение Т 0 • Модуль поверхностного изотермического сжатия (модуль Юнга) • Поверхностный модуль упругости при сдвиге

Силы, действующие на мембрану • • • Р –сжимающие f - растягивающие h – толщина мембраны

Сравнение свойств искусственных липидных и биологических мембран Свойство Биомембран БЛМ ы Электронно-микроскопический снимок поперечного среза Трехслойная структура Трехслойн ая структура Толщина, нм 6, 0 -10, 0 2, 5 -8, 0 Сопротивление, Ом*см 2 Электрическая емкость, мк. Ф/см 2 Показатель преломления 102 -105 0, 5 -1, 3 106 -109 0, 2 -1, 0 1, 6 1, 56 -1, 66 Проницаемость для воды, мкм/с 0, 5 -400 31, 7
 • Гемолиз (разрушение эритроцитов с выделением гемоглобина)")
Разрушение БМ Слияние клеток • Лизис (разрушение) • Гемолиз (разрушение эритроцитов с выделением гемоглобина) Липидные мембраны – метастабильные системы Значительные отклонения параметров БМ от равновесных, приводят к возникновению и накоплению дефектов в структуре •

Сквозная пора – критический дефект Тепловые флуктуации Электрический пробой Замораживание Действие ПАВ Осмотическое давление Др

Поры в БМ
 R>R* (КРИТИЧЕСКИЙ РАДИУС ПОРЫ) Пора залечивается Необратимое разрушение")
Размер поры R<R* (КРИТИЧЕСКИЙ РАДИУС ПОРЫ) R>R* (КРИТИЧЕСКИЙ РАДИУС ПОРЫ) Пора залечивается Необратимое разрушение мембраны - гибель
 – расширение поры h р(r")
Модель критической поры р h/2 2 r 0 р(h/2) – расширение поры h р(r 0) – сжатие поры

Энергия образования поры

В ЖК фазе БМ

Образование поры в ЭП– электрический пробой

В ЖК фазе БМ при электрическом пробое

Энергия поры при различных значениях мембранного потенциала
 при наложении разности потенциалов (2)")
Энергия дефекта в БМ в отсутствие ЭП (1) при наложении разности потенциалов (2)
 и гель - состоянии (Б) мембранных липидов")
Энергия поры в жидкокристаллическом состоянии БМ (А) и гель - состоянии (Б) мембранных липидов

Размеры липидных пор в модельных и клеточных мембранах Радиус поры r, нм Объект Соотношение радиусов пор Стрессовое состояние 3, 0 -4, 0 Эритроцит rжк с >rэп>=r Электрический пробой 2, 0 То же rжк с >r Осмотический гемолиз 1, 2 L-клетки rжк с >rэп>=r Электрический пробой 0, 2 -2, 0 Липосомы rжк с >r Осмотический лизис 0, 6 -0, 8 То же rжк с >rгель>=r Фазовый переход 0, 5 -2, 0 То же rжк с >rгель>=r Фазовый переход 1, 2 -1, 8 БЛМ rжк с >rгель>r>rэп гель Фазовый переход

Зависимость изменения энергии поры от радиуса

ТРАНСПОРТ ВЕЩЕСТВ ЧЕРЕЗ БИОЛОГИЧЕСКИЕ МЕМБРАНЫ Виды транспорта. Пассивный транспорт

• • • Живые системы - открытые системы на всех уровнях организации Необходимое условие существования клетки – транспорт веществ через БМ, который обеспечивает: -метаболизм клетки -биоэнергетические процессы создания потенциалов и генерации нервного импульса Нарушение транспортной функции БМ – развитие патологии

Потенциалы в клетке ХИМИЧЕСКИЙ ЭЛЕКТРОХИМИЧЕСКИЙ

Химический потенциал

Электрохимический потенциал

Виды транспорта через БМ Пассивный транспорт перенос вещества без затраты энергии Активный транспорт перенос вещества с затратами энергии Пассивный транспорт - это транспорт самопроизвольный, без затраты энергии, "под гору". Активный - требует затраты энергии.

Пассивный транспорт через БМ осмос фильтрация Простая диффузия Облегченная диффузия С подвижным переносчиком через липидный слой через липидные поры через Белковые поры С фиксированным переносчиком

Простая диффузия: а – через липидный слой б – через липидные поры в – через белковые поры

Пассивный транспорт Перенос k-ого вещества по градиенту ЭХП, то есть из мест с большим значением ЭХП к местам с меньшим значением ЭХП С 1 м мембрана > l С 2 м С 2 jm
 – величина, равная количеству вещества, перенесенного за единицу времени через единицу")
Плотность потока (J) – величина, равная количеству вещества, перенесенного за единицу времени через единицу площади поверхности, перпендикулярной направлению переноса [J]=1 моль/с*м 2 Уравнение Теорелла – плотность потока к-го сорта вещества при пассивном переносе

Причины пассивного транспорта Градиент концентрации Градиент ЭП

1 закон Фика С 1 м мембрана > l С 2 м С 2 jm


Нестационарная диффузия Jk S x x+dx x

Количество вещества, накапливающегося в объеме

Коэффициент диффузии Dk Зависит от размера и формы молекул Для малых молекул Для сферических молекул
 Органические кислоты")
Проницаемость мембран - Хорошая для: Неполярных веществ (хорошо растворимые в липидной фазе) Органические кислоты эфиры - Плохая для: Полярных (водорастворимых) веществ Соли Основания Спирты Сахара Аминокислоты

Образование кинков а – транс-конфигурация б - гош-транс-гош конфигурация

Пассивный транспорт через липидные поры Липидные поры –гидрофильные поры в липидном бислое Белковые поры

ЛИПИДНЫЕ ПОРЫ Размеры канала изменяются в зависимости от внешних условий и имеют динамических характер Размеры варьируются в широких пределах, поры могут «затекать» Нет выраженной избирательности каналов - универсальны БЕЛКОВЫЕ ПОРЫ Размер сохраняется на протяжении всей жизни поры Фиксированный набор радиусов Избирательность переноса

Пассивный транспорт: облегченная диффузия С ПОДВИЖНЫМ ПЕРЕНОСЧИКОМ С ФИКСИРОВАННЫМ ПЕРЕНОСЧИКОМ Отличия от простой диффузии: 1. Значительно быстрее 2. Имеет свойство насыщения (все молекулы переносчики - заняты) 3. Высокая специфичность 4. Вещества – блокаторы облегченной диффузии (ингибиторы)

Схема переноса валиномицином ионов калия через мембрану

Зависимость плотности потока веществ через БМ в клетку в зависимости от соотношения концентраций 1 – простая диффузия 2 – облегченная диффузия

Пассивный транспорт - осмос • Диффузия воды из мест с ее большей концентрацией в места с меньшей концентрацией Осмотическое давление Вода р1 р2

Пассивный транспорт фильтрация Движение раствора через поры под действием градиента давления
, облегченная диффузия через канал в мембране (в середине) электрофорез ионов -")
Простая диффузия (вверху), облегченная диффузия через канал в мембране (в середине) электрофорез ионов - внизу.

Насыщаемый и ненасыщаемый транспорт ионов. При обычной диффузии потоки невелики, но прямо пропорциональны концентрации иона в среде, из которой происходит перенос (нижняя прямая). При переносе через канал или с помощью подвижного переносчика потоки гораздо больше, но при увеличении концентрации ионов наступает насыщение.

Активный транспорт веществ через БМ

Активный транспорт • • • Перенос k-ого вещества против градиента ЭХП, то есть из мест с меньшим значением ЭХП к местам с большим значением ЭХП Сопровождается увеличением энергии Гиббса Не может идти самопроизвольно, а только в сопряжении с процессом гидролиза АТФ, те за счет энергии, запасенной в макроэргических связях

Схема активного транспорта k-ого вещества против градиента ЭХП, то есть из мест с меньшим значением ЭХП к местам с большим значением ЭХП Перенос k(х1) мембрана < l k (х2) jm

Функции АТ • Создание градиента концентрации вещества • Создание градиента электрического потенциала • Создание градиента давления АТ УДЕРЖИВАЕТ ОРГАНИЗМ В НЕРАВНОВЕСНОМ СОСТОЯНИИ

Опыты Уссинга: 1949 г. АТ показан на примере переноса ионов натрия через кожу лягушки

• • АТ ионов Na+ : в коже лягушки близок к переносу ионов через эпителий в кишечнике и почечных канальцах В нервных клетках локализован в плазматических мембранах АТ ионов Сa++ : В мембранах саркоплазматического ретикулума АТ ионов Н+ : В энергосберегающих мембранах митохондрий и хролопластов

Механизмы АТ Ионные - - - насосы Na+ насосы: локализованы в плазматических мембранах нервных клеток Сa++ насосы: локализованы в мембранах саркоплазматического ретикулума Н+ насосы (помпа): локализованы в энергосберегающих мембранах
")
Na+ насос (Na-К-АТФ-фаза)
")
Сa++ насос (Са-АТФ-фаза)
")
Н-насос (протонная помпа)

Схема механизма Na-К-АТФ-фазы

Схема механизма Na-К-АТФ-фазы • • 1. образование комплекса фермента с АТФ на внутренней поверхности мембраны 2. связывание комплексом 3 -х ионов натрия 3. фосфорилирование фермента с образованием АДФ 4. переворот (флип-флоп) фермента внутри мембраны 5. реакция ионного обмена натрия на калий на внешней поверхности мембраны 6. обратный переворот ферментного комплекса с переносом ионов калия внутрь клетки 7. возвращение фермента в исходное состояние с освобождением ионов калия и неорганического фосфата

Активный транспорт Электронейтральный Электрогенный Функционирование транспортной системы сопровождается обменом внутриклеточных ионов на внеклеточные в отношении «заряд на заряд» количество зарядов, переносимых за единицу времени в одном направлении НЕ компенсируется суммарным зарядом, переносимым в противоположном направлении Создаются дополнительные разности потенциалов

Индуцированный ионный транспорт Процессы переноса ионов через БМ, обусловленные наличием механизмов, облегчающих обмен ионами между внеклеточной средой и внутриклеточной средой
:")
Затраты энергии, необходимые для проникновения ионов из воды в БЛМ (формула Борна):

W: Барьер для прохождения ионов щелочных металлов через гидрофобную часть липидного бислоя Крупные органические ионы проникаю легче (r W ) воды БЛМ W Перенос ионов ИОНОФОРАМИ

Ионофор, образующий комплекс • • Могут образовывать с ионом комплекс большого радиуса – переносчик «малая карусель» и «большая карусель» Формируют пору в БМ, заполненную водой – ионные каналы b k b r

Каналообразующий ионофор Внешняя часть молекулы – гидрофобна, внутренняя – гидрофильна На одном из концах молекулы – «якорь» заряженные и сильно полярные группы, которые удерживают молекулу на одной стороне мембраны и позволяет ей пронизывать гидрофобную часть БМ

Виды каналообразующих ионофоров Грамицидин А-цепь из 15 гидрофобных аминокислот спираль полый цилиндр пора низкая селективность из-за высокой эластичности спиральной структуры • Аламецитин-пептидный антибиотик – 20 аминокислот в линейной цепи водная пора переменного диаметра • • Полиеновые антибиотики

Вторичный АТ: унипорт

Вторичный АТ: антипорт

Вторичный АТ: симпорт

БИОЛОГИЧЕСКИЕ ПОТЕНЦИАЛЫ Потенциал покоя

Биологические потенциалы • Основная функция БМ-генерация и передача биопотенциалов • Основа возбудимости клеток Регуляция внутриклеточных процессов Регуляция работы нервной системы Регуляция мышечного сокращения Регуляция рецепции • •
 Электроэнцефалография (активность головного мозга) Электромиография (мышечная активность)")
ЭП позволяют Электрокардиография (активность сердца) Электроэнцефалография (активность головного мозга) Электромиография (мышечная активность)

Электрические потенциалы • Окислительно-восстановительные – вследствие переноса электронов от одних молекул к другим Мембранные – возникают вследствие градиента концентрации ионов по разные стороны мембраны Мембранные потенциалы – биопотенциалы, которые в основном регистрируются в организме •

Экспериментальное исследование БП Разработка микроэлектродного метода внутриклеточного измерения БП Создание специальных усилителей БП Выбор объекта исследования – крупные клетки – АКСОН КАЛЬМАРА ( 0, 5 мм > позвоночных)

Экспериментальное исследование БП Стеклянная пипетка стеклянный электрод

Схема регистрации мембранного потенциала

Виды потенциалов Потенциал покоя Потенциал действия - разная концентрация ионов - диффузия ионов через БМ

Потенциал покоя Стационарная разность потенциалов в клетке, регистрируемая между наружной и внутренней поверхностью мембраны, находящейся в невозбужденном состоянии Обусловлен различными концентрациями ионов во внутриклеточном и наружном растворах

- разная концентрация ионов - диффузия ионов через БМ -Свн Снар -Мембрана проницаема Поток заряженных частиц через БМ Нарушение электронейтральности системы Потенциал препятствует дальнейшему перемещению ионов через БМ Равновесие ЭХП

Равновесие ЭХП формула Нернста-Планка для равновесного мембранного потенциала

Значения потенциала покоя

Формула Нернста-Планка для равновесного мембранного потенциала

Электродиффузионный транспорт через БМ Элетродиффузия – пассивный транспорт ионов через БМ 1. Отсутствуют суммарные ионные токи: Потенциал Нернста определяется концентрацией ионов в растворе 2. Ионные токи не равны 0: Пассивное движение ионов по градиенту ЭХП возникает диффузионная разность потенциалов, которая зависит от: Ионных концентраций Подвижности ионов

Способы описания процесса перехода ионов через БМ • Дискретный • Непрерывный Ионы преодолевают БМ с помощью нескольких дискретных перескоков через активационные барьеры Ионы движутся через БМ независимо и не взаимодействуя друг с другом. БМ является гомогенной фазой Описание транспорта ионов через селективные каналы клеточной мембраны Описание транспорта ионов через искусственные БЛМ

Непрерывное описание диффузии • Уравнение Нернста-Планка: 1. ЭХ равновесие 2. Отсутствие ЭХ равновесия 3. Приближение постоянного поля

ЭХ равновесие Потенциал Нернста – определение равновесной разности потенциалов на БМ – мембранный потенциал

Потенциал Нернста для одновалентных ионов 1902 г Берштейн – причина возникновения мембранного потенциала – диффузия ионов К+ из клетки во внеклеточную среду

Отсутствие ЭХ равновесия Подход Гендерсона-Планка: - Концентрация катионов и анионов в любой плоскости, перпендикулярной направлению переноса – одинаковая: С+=С-=С - В стационарном состоянии электрический ток через БМ отсутствует - Для бинарного электролита: Z+=-Z-=1

Отсутствие ЭХ равновесия

Уравнение Гендерсона U+ U—- подвижность катиона и аниона в БМ расчет диффузионного потенциала, возникающего между двумя электролитами Уравнение Гендерсона применимо: -Для мембран макроскопической толщины 1 мкм -Не применимо для липидных и клеточных мембран, где не выполняется условие локальной электронейтральности по всей толщине мембраны

Приближение постоянного поля Предположение о линейности изменения потенциала ЭП по всей толщине БМ Для тонких БМ, в которых концентрация носителей мала, а толщина двойного слоя – велика

Приближение постоянного поля Снаружи 2 1 Внутри м h x J 0 Co=KC 1 м C 2 м Ci=KC 2 м

Профиль поля - линеен
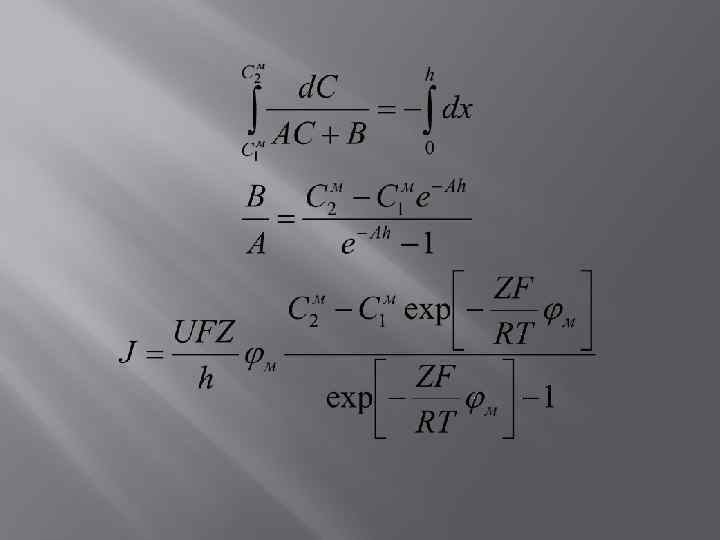


Р – коэффициент проницаемости БМ К – коэффициент растворения, который зависит от липофильности иона 1943 Гольдман + 1949 Ходжкин и Катц

Разность потенциалов, создаваемая в БМ в результате совместной диффузии ионов В равновесии

- - Потенциал на БМ определяется: Различием в стационарных концентрациях ионов по обе стороны БМ Разными коэффициентами проницаемости БМ для различных ионов

Соотношение Уссинга-Теорелла

Критерии пассивного транспорта - - - Поток ионов через БМ обусловлен: Только градиентом концентрации ионнов к-го типа Действием ЭП Нарушение соотношения Уссинга-Теорелла – существование АКТИВНОГО ТРАНСПОРТА

ПОТЕНЦИАЛ ДЕЙСТВИЯ

Потенциал действия Открыт в 18 в. Луиджи Гальвани: 1. мышечные сокращения препарированной лягушки могут вызваться электрическим импульсом 2. сама живая система является источником электрического импульса
")
Потенциал действия 19 в. Гельмгольц: показано, что скорость распространения нервного импульса (1 -100 м/с) значительно меньше скорости распространения электрического импульса по проводам (3*108 м/с)

Потенциал действия 20 в. А. Ходжкин: нервный импульс-импульс электрического тока 1963 г. Ходжкин, Хаксли и Иклс – Нобелевская премия по медицине «за оперирование нервных клеток»

Потенциал действия Электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения Методы: 1. Микроэлектродов с использованием высокоомных измерителей напряжения 2. Меченных атомов •

Исследование потенциала действия: схема опыта Г – генератор импульсов Р – регистратор напряжения

Регистрация потенциала действия

Потенциал действия – ПД – не зависит от амплитуды деполяризующего потенциала

Свойства ПД 1. 2. 3. 4. Наличие порогового значения деполяризующего потенциала Закон «все или ничего» Период рефрактерности Резкое снижение сопротивления БМ в момент возбуждения (покой: 0, 1 Ом*м 2, возбуждение: 0, 0025 Ом*м 2)

Положительный потенциал реверсии имеет Na природу

Проницаемость БМ В состоянии покоя: РК : РNa : РCl = 1 : 0, 04 : 0, 45 В состоянии возбуждения: РК : РNa : РCl = 1 : 20 : 0, 45

Уравнение Ходжкина-Хаксли

Эквивалентная электрическая схема элемента возбудимой мембраны

Т. Х-Х: Возбуждение элемента мембраны связано с изменением проводимости мембраны для ионов натрия и калия

Опыты с фиксацией напряжения 1. избавиться от емкостных токов 2. исключить изменение ионных проводимостей натрия и калия при изменении мембранного потенциала и изучить их изменение в различные фазы развития возбуждения

Схема исследования токов через мембрану с фиксацией мембранного потенциала 1. 2. 3. 4. 5. 6. Микроэлектрод Электрод сравнения Серебряный проводник Генератор постоянного напряжения Амперметр ОУ – операционный усилитель

Результаты исследования мембранного тока методом фиксации напряжения

Изменение проводимости БМ для ионов во время развития ПД

Распространение нервного импульса вдоль возбудимого волокна Возбудимость – способность клеток к быстрому ответу на раздражение, который проявляется через совокупность физических, физикохимических процессов и функциональных изменений. Нервная, мышечная, железистая Признак возбуждения: изменение электрического состояния клеточной мембраны: возбужденный участок клетки – электроотрицателен по отношению к невозбужденному участку

Потенциал действия - - Общее изменение разности потенциалов между клеткой и средой, происходящее при пороговом и сверхпороговом возбуждении клеток Обеспечивает проведение возбуждения по нервным волокнам Индуцирует процессы мышечного сокращения Индуцирует секрецию железистых клеток

Распространение потенциала действия по аксону кальмара

Распространение нервного импульса вдоль нервного волокна

Деполяризующий потенциал

Сальтаторное распространение ПД по миелинизированному волокну
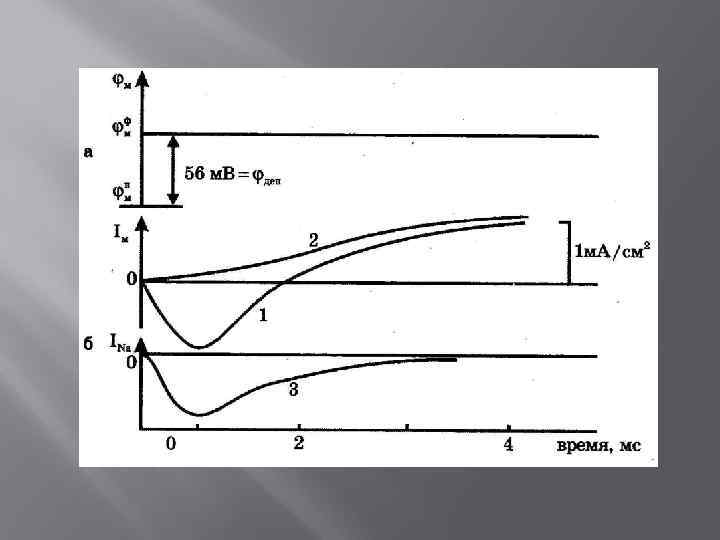

Натриевый и калиевый токи через мембрану миелинизированного аксона лягушки

Зависимость стационарных значений параметров Na-канала от мембранного потенциала

Дискретный характер проводимости ионных каналов
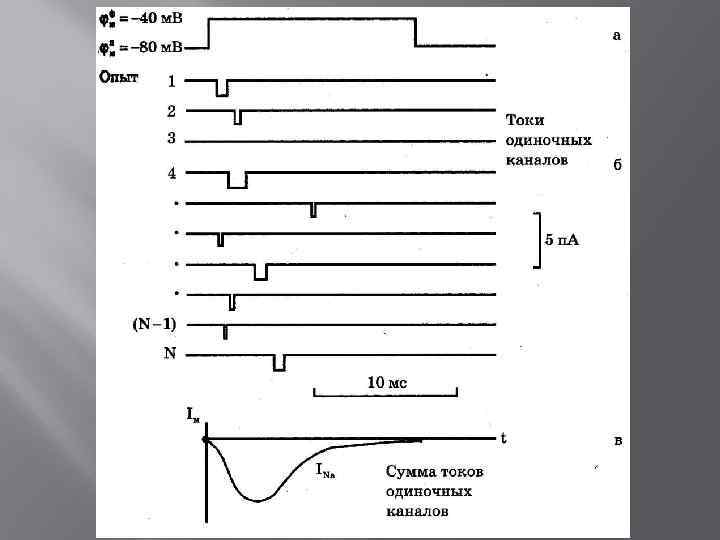

Схема строения Na-канала

Механизм генерации потенциала действия

Распределение концентрации ионов внутри и снаружи кардиомиоцита позвоночных

Процессы, происходящие при формировании ПД
Презентация по биофизике для ДИТИ МИФИ Ч3.ppt