Bioenergetika_I_S_Grig_1l_II.ppt
- Количество слайдов: 95



Биоэнергетика И. С. Григ.

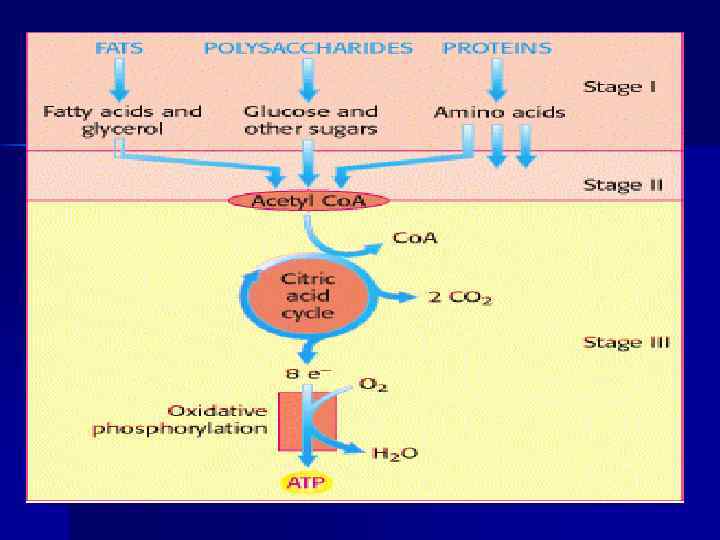
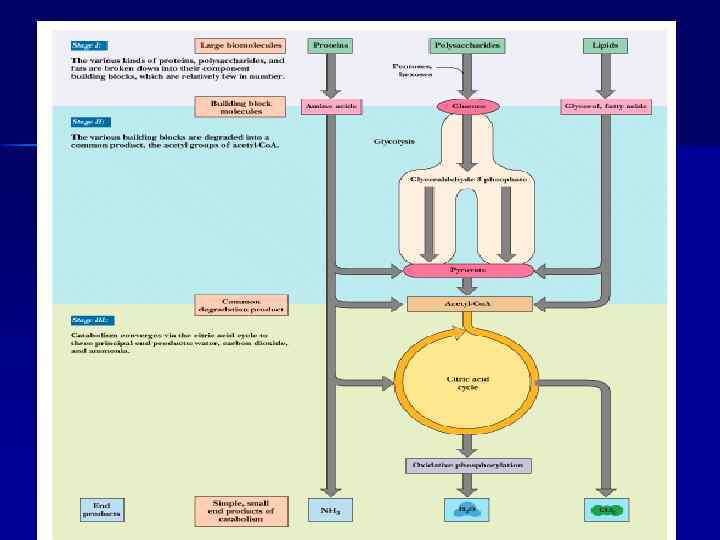
Обмен веществ включает 3 этапа: n поступление веществ в организм, n метаболизм, n выделение конечных продуктов из организма. Поступление веществ в организм происходит в результате дыхания (кислород) и питания. В ЖКТ продукты питания перевариваются (расщепляются до простых веществ). При переваривании происходит гидролиз полимеров (белков, полисахаридов и др. сложных органических веществ) до мономеров, всасывающихся в кровь и включающихся в промежуточный обмен.

Гетеротрофы. Аутотрофы • Живые организмы сточки зрения термодинамики — открытые системы. Между системой и окружающей средой - обмен энергии. n Живые существа по способу питания аутотрофы и гетеротрофы. Аутотрофы - используют СО 2 как источник углерода и трансформируют энергию n n солнечного света в энергию химических связей органических веществ; У гетеротрофов катаболизм орг. веществ приводит к освобождению энергии и запасанию её в виде АТФ. Основной путь получения энергии гетеротрофами - окисление органических веществ путем дегидрирования.

Гетеротрофы. Аутотрофы.

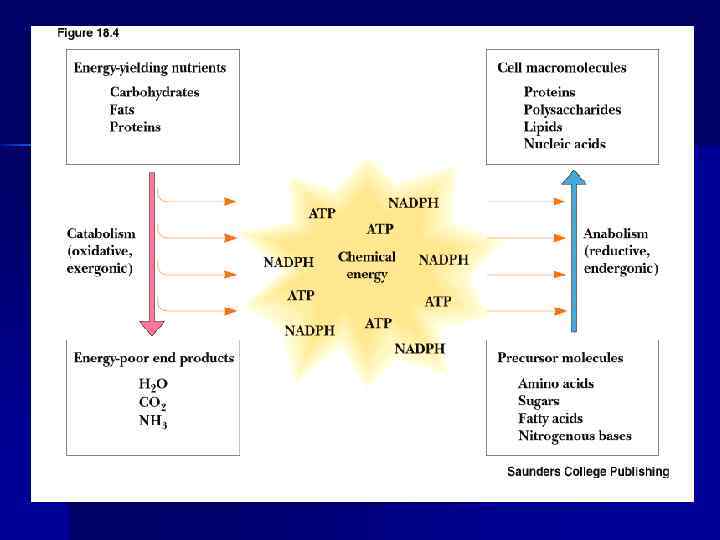
Метаболизм. Катаболизм. Анаболизм. n Метаболизм — совокупность химических реакций, происходящих в организме. n n n Метаболический путь — последовательность реакций, приводящих к образованию определенного продукта. Соединения, образующиеся в ходе превращений — метаболиты. В метаболизме можно выделить пути катаболизма, ведущие к расщеплению сложных молекул и пути анаболизма, предназначенные для биосинтезов.

Метаболические пути

Эндергонические и экзергонические реакции В результате катаболизма образуются конечные продукты обмена —СО 2 , Н 2 О и мочевина. Реакции катаболизма – - экзергонические реакции, сопровождаются выделением энергии. n n Анаболические реакции - эндергонические реакции, используют энергию, освобождающаяся при катаболизме. n Катаболические и анаболические пути во многом различаются, но они тесно связаны друг с другом. Катаболизм и анаболизм — это сопряженные взаимодополняющие процессы.

Эндергонические и экзергонические реакции n Каждое органическое вещество, входящее в состав живой материи, обладает определенным запасом внутренней энергии - Е. Часть этой внутренней энергии может быть использована для совершения полезной работы, это свободная энергия — G.

Эндергонические и экзергонические реакции n Направление химической реакции определяется значением G. Если G - , то реакция протекает самопроизвольно и сопровождается уменьшением G. n Реакции, сопровождающиеся уменьшением G — экзергонические. n Если G +, то реакция будет протекать только при поступлении свободной энергии извне, такие реакции называют эндергоническими.

Сопряжение эндергонических и экзергонических процессов n Термодинамически невыгодные эндергонические реакции протекают за счет энергии экзергонических реакций. n Процессы, протекающие с потреблением и выделением энергии, связаны между собой, происходят при участии АТФ. n АТФ — сопрягающий фактор, форма запасания энергии.

Роль АТФ в клеточной энергетике n Химическая энергия, освобождаемая в процессе катаболизма, запасается путем фосфорилирования АДФ с образованием АТФ; n Энергия АТФ затем используется ходе эндергонических реакций (процессы биосинтеза, активный транспорт, сокращение мышц и т. д. ). n Основные источники энергии: углеводы, жиры, белки.
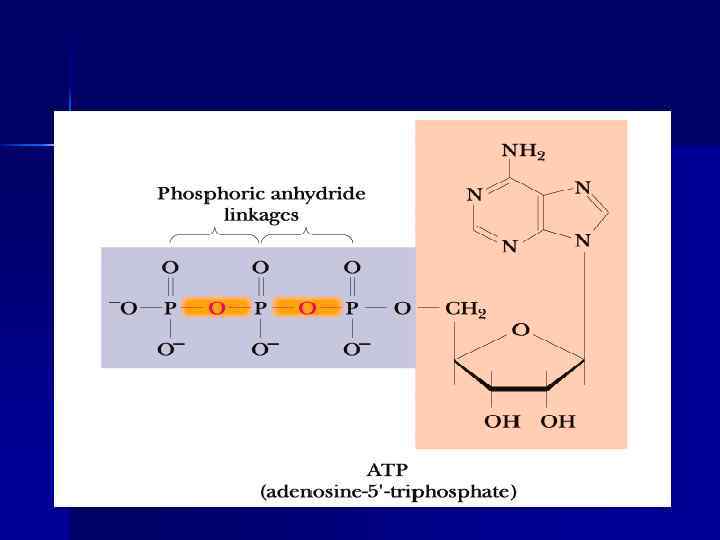

АТФ и НАДФ

АТФ, АДФ, АМФ

Роль АТФ

Роль окислительно-восстановительных процессов в метаболизме n Основной путь получения энергии гетеротрофами - окисление органических веществ путем дегидрирования. Его роль: n обеспечение организма энергией; n терморегуляция

Другие функции биоокисления в клетке Роль прямого присоединения к S кислорода: n образование биологически важных метаболитов. n обезвреживание токсических веществ.

Макроэргические соединения n Если энергия, освобождающаяся при гидролизе веществ, превышает 30 к. Дж/моль (7, 3 ккал/моль), то гидролизуемая связь – высокоэнергетическая, (макроэргическая, ~ ). n Макроэргические соединения — вещества, содержащие в своем составе высокоэнергетические (макроэргические ) связи, при гидролизе которых высвобождается более 30 к. Дж энергии на 1 моль связей.

Макроэргические соединения n n n n n К ним относят: нуклеозидтрифосфаты (АТФ, ГТФ, УТФ, ЦТФ), нуклеозиддифосфаты, фосфоэнолпируват, 1, 3 дифосфоглицерат, карбамоилфосфат, сукцинил-Ко. А, ацетил-Ко. А, креатинфосфат, пирофосфат

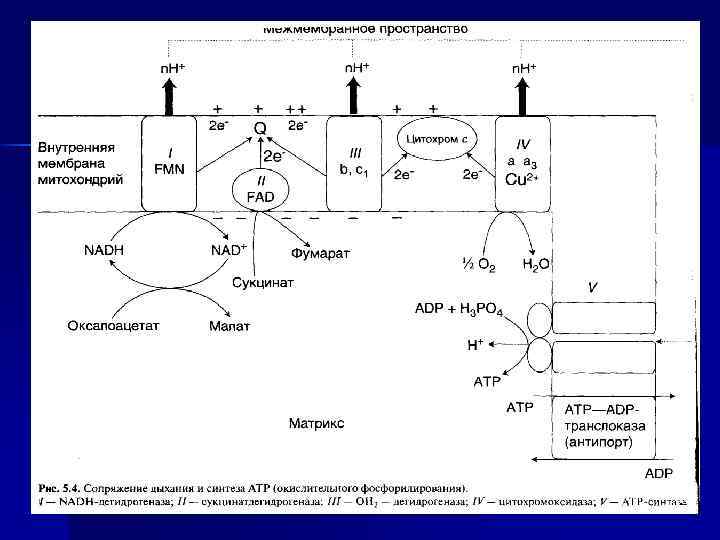
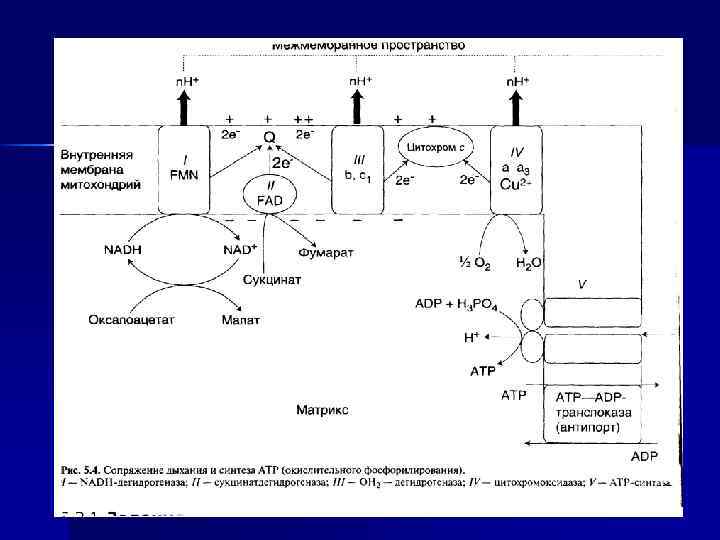
Виды фосфорилирования как реакции образования АТФ: окислительное, субстратное, фотофосфорилирование Окислительное фосфорилирование n Синтез АТФ из АДФ и Н 3 РО 4 за счет энергии переноса е- по ЦПЭ на О 2 - окислительное фосфорилирование n АДФ + Фн АТФ + Н 2 О n Энергетическое сопряжение реакций переноса Н (е- и Н+) и синтеза АТФ происходит при участии МТХ мембраны и Н+-АТФ-синтетазы.
 высвобождается")
Субстратное фосфорилирование, фотофосфорилирование n субстратное фосфорилирование — энергия, необходимая для образования АТФ (ГТФ) высвобождается при гидролизе связей других макроэргических соединений. Синтез АТФ происходит за счет энергии макроэргических связей S. Механизм сопряжения не требует участие мембраны.

Субстратное фосфорилирование, фотофосфорилирование Гликолиз: n ФЭП +АДФ АТФ + ПВК; n 1, 3 дифосфоглицерат + АДФ АТФ + 3 -фосфоглицерат; ЦТК: сукцинил-Ко. А сукцинат. Мышцы: креатинфосфат + АДФ креатин + АТФ n фотофосфорилирование — для синтеза АТФ используется энергия электромагнитного излучения (видимого света), которая запасается в виде электрохимического потенциала (у зеленых растений).

Субстратное фосфорилирование

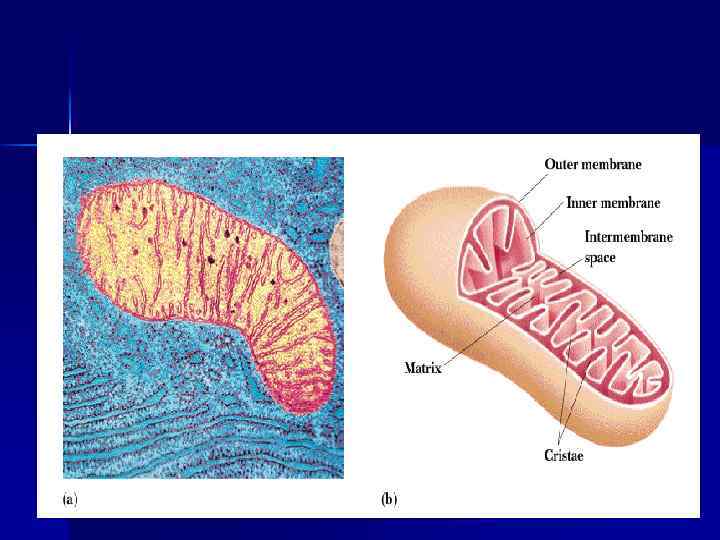
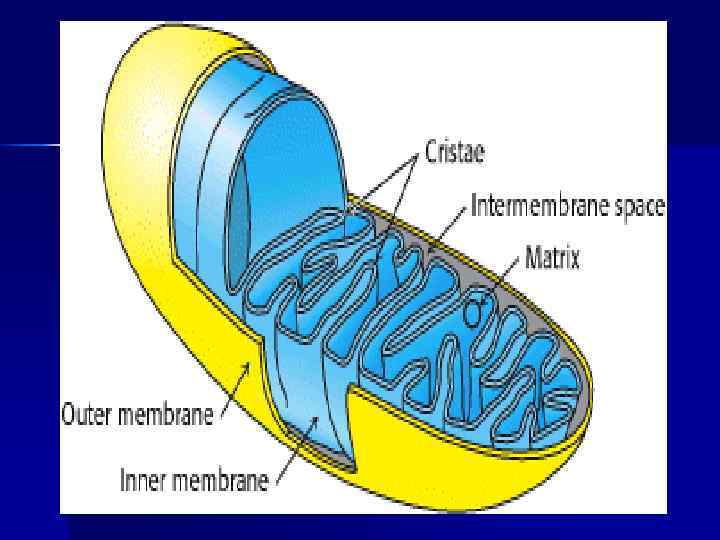
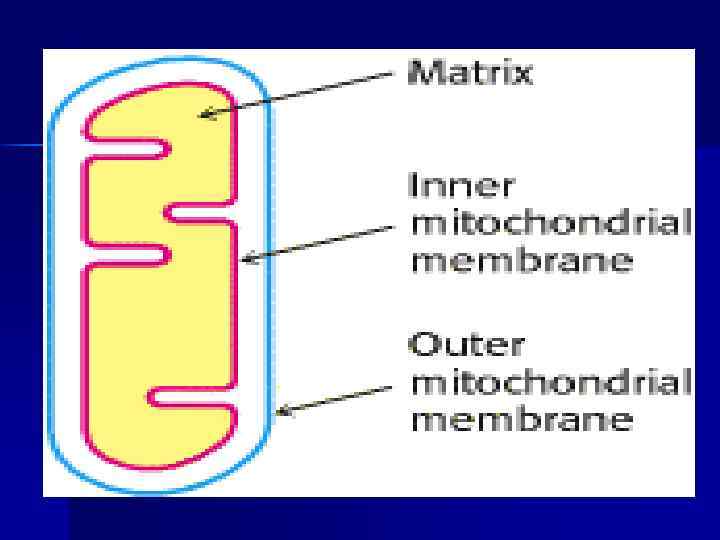
Тканевое дыхание. Дыхательная цепь n Окисление орг. веществ в клетках, сопровождающееся потреблением О 2, образованием Н 2 О и освобождением энергии — тканевое дыхание. n Тканевое дыхание включает: а)отнятие Н (е- и Н+) от S (дегидрирование); б) многоэтапный процесс переноса е- на О 2. n Тканевое дыхание и синтез АТФ энергетически сопряжены.

Тканевое дыхание. Дыхательная цепь n О 2 в процессе биоокисления используется как акцептор Н окисляемых (дегидрирумых) S, в результате чего синтезируется Н 2 О (400 мл в сутки) -2 Н+ SH 2 + ½ О 2 S + Н 2 О+220 к. Дж/моль. -2 е
 — путь переноса Н+ и е- от органических")
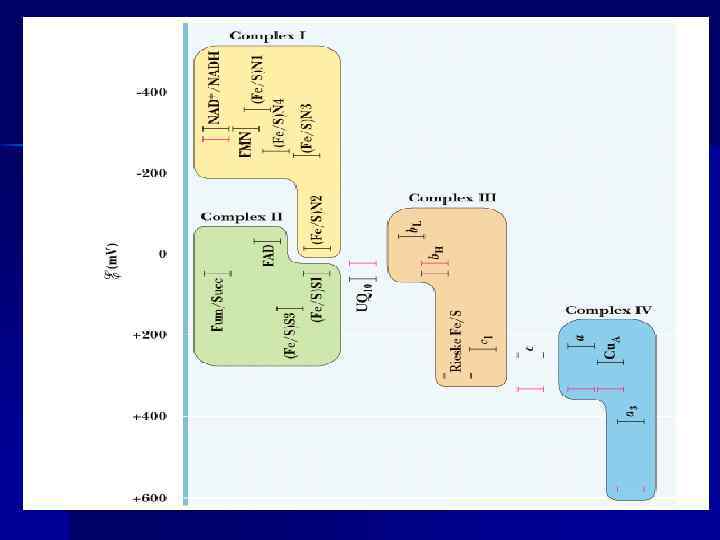
Дыхательная цепь n Дыхательная цепь (ЦПЭ) — путь переноса Н+ и е- от органических веществ на О 2 воздуха. Это система переносчиков, структурно закрепленная на мембране МТХ и расположенная в соответствии со своими редокс-потенциалами. n Последовательность расположения компонентов дыхательной цепи определяется окислит. -восстановит. потенциалами их окисленных и восстановленных форм.
. n")
В митохондриях процессы: n Перенос е- с окисляемых S на О 2 (дыхание). n Перенос Н+ из матрикса МТХ в межмембранное пространство и образование трансмембранного электрохимического потенциала. n Синтез АТФ за счет потока Н+ из мембранного пространства в матрикс.

Окислительно-восстановительные реакции. Окислительновосстановительный потенциал n Окисление – отщепление е-, присоединение е- —восстановление. n Окисленные и восстановленные формы одного и того же соединения сопряженные пары, или редокс пары. Мерой сродства редокс-пары к е - — окислительно-восстановительный потенциал или редокс-потенциал (Е 0 ). Выражается в вольтах, чем меньше (отрицательнее), тем меньше сродство к е-. Чем больше сродство, тем больше окислительно-восстановительный потенциал. n

Редокс-потенциал n Е 0 NAD+/NADH = - 0, 32 v, + + 2 е- /Н О = + Е 0 ½ О 2 + 2 Н 2 О, 82 v. n. Разные редокс- потенциалы — разное -. Те, у кого сродство к е сродство <, отдают е - тем, у кого оно >.

Уравнение Нернста Е 0 связано с G 0 уравнением: G 0 = - n F E 0 , где п — число перенесенных е-; F – постоянная Фарадея (23 061 ккал/моль вольт); E 0 — разность окислительно-восстановительных n потенциалов электрон-донорной и электрон-акцепторной систем = [0, 82 – (-0, 32) =1, 14]. G 0 = 220 к. Дж/моль (52, 6 ккал/моль) n Положительное значение E 0 указывает на экзергонический характер реакции, протекающей в стандартных условиях. E 0 - стандартная величина окислит-восстановит. потенциала (определяется в стандартных условиях). Концентрация веществ = 1 М, Р газов = 1 атм. , а р. Н = 7, 0.

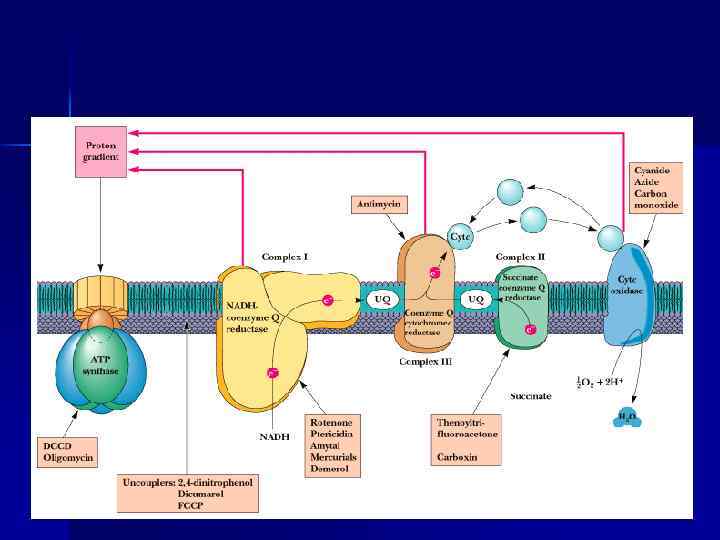
Строение дыхательной цепи n n n Во внутренней МТХ мембране расположены четыре белковых комплекса. Жирорастворимый кофермент коэнзим Q и водорастворимый цитохром c перемещаются между белковыми комплексами. Перемещение электронов в дыхательной цепи происходит по градиенту окислитвосстановит. потенциала.
")
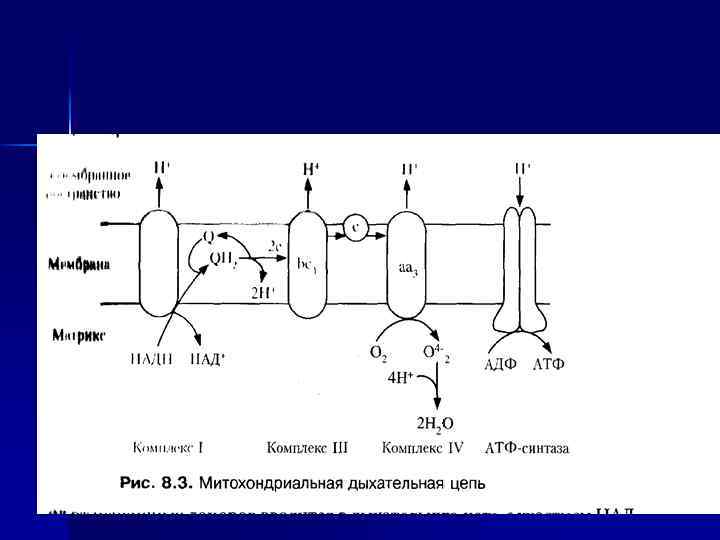
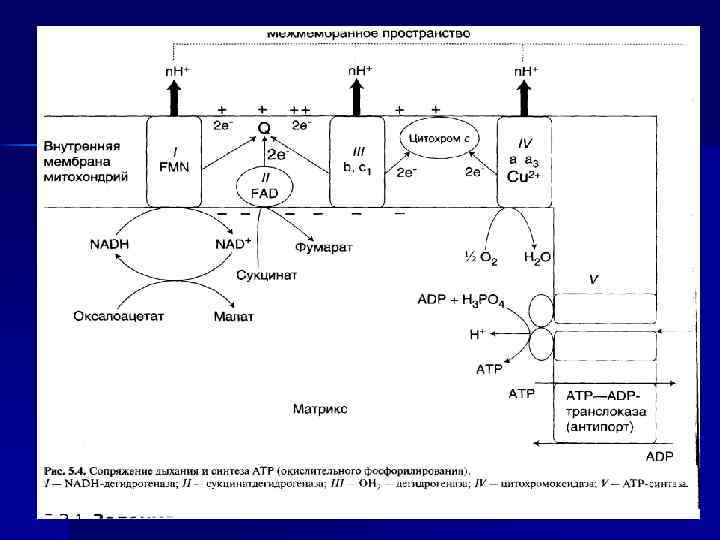
I, III, IV, V комплексы n НАДН-дегидрогеназа (НАДН – Ко. Q- редуктаза, комплекс I) - FMN содержащий Е. n Сукцинат-дегидрогеназа (СДГ)— комплекс II (кофермент ФАД). n Ко. Q 2 -дегидрогеназа (коэнзим. Q – цит. С-редуктаза) — комплекс III. n ЦТХ-оксидаза — комплекс IV. n Н+-АТФ-синтетаза — комплекс V.

Митохондриальная цепь переноса е

Митохондриальная цепь переноса е

Е и коферменты дыхательной цепи n За исключением Ко. Q — все компоненты — белки. Субстраты НАД и НАДФ-зависимых дегидрогеназ — в матриксе МТХ и в цитозоле. Первичные доноры Н для НАДзависимых дегидрогеназ: пируват, изоцитрат, α-кетоглутарат, малат, глутамат. n У аэробных организмов конечным акцептором е-, получаемых от молекул, является О 2.
 —")
Комплекс I n НАДН-дегидрогеназы ((НАДН – Ко. Q-редуктаза, комплекс I, FMN содержащий Е) — во внутренней мембране, окисляет НАДН, образующийся в МТХ матриксе. n S — НАДН (перенос 2 е- и Н+ на FMN. 2 й Н+ — из матрикса).

ФМН

Пиридиновые нуклеотиды: НАД+ и НАДФ+

Окисленная и восстановленные формы НАД

Окисленная и восстановленные формы НАД

Комплекс II. n n Fe S центры СДГ — комплекс II. ФАД-зависимые дегидрогеназы (СДГ, дегидрогеназа ЖК) переносят Н на убихинон (коэнзим Q, Ко. Q). Суммарная реакция: сукцинат + коэнзим Q фумарат + коэнзим QH 2 Первичные доноры Н для ФАД-зависимых дегидрогеназ - сукцинат, ацил-Ко. А, α-глицерофосфат. n Большинство ФАД-зависимых дегидрогеназ — локализовано в матриксе МТХ, за исключением СДГ (во внутренней мембране МТХ).

Комплекс II.

ФАД

ФМН. ФАД

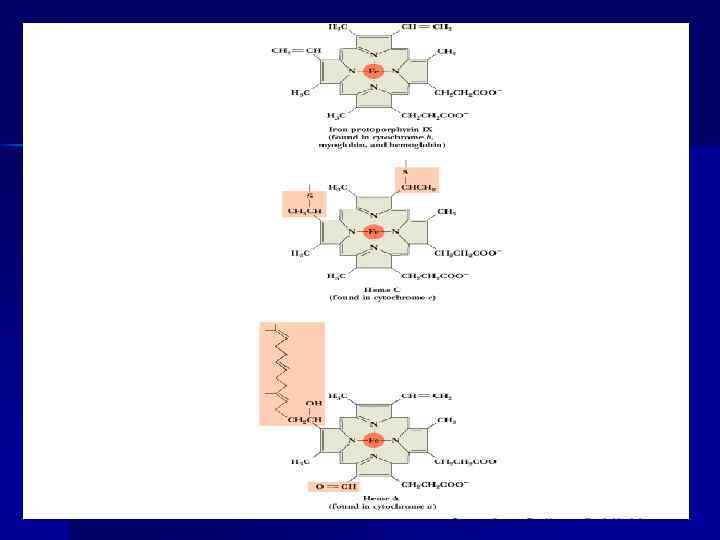
Fe S центры n Fe S центры входят в состав ферментных комплексов I, II и III (во флавопротеидах, ЦТХ). n содержат негемовое Fe. n Атомы Fе могут отдавать и принимать е- поочередно, переходя в ферро (Fe 2+) и ферри (Fe 3+) состояния.

Типы железо-серных центров

Комплекс III n n n Ко. Q — коллекторная функция, присоединяет Н от НАДНдегидрогеназы и др. флавинзависимых дегидрогеназ (СДГ). Ко. Q-дегидрогеназа (коэнзим Q – цит. С-редуктаза) — комплекс III. Ко Q передаёт электроны на цитохром с и выкачивает протоны в межмембранное пространство (Q цикл);

Убихинон

Убихинон

Ко. Q
 на")
Комплекс III n n Ко. Q-дегидрогеназа переносит е- от убихинола (Ко. QН 2) на цит. С. Внутри комплекса III епереносится от цит. В на Fe. Sцентры, на цит. С 1, а затем на цит С. Гем цитохромов, подобно Fe. S центрам переносит только по 1 е-. Цит. С - периферический водорастворимый мембранный белок, сод. 1 полипептидную цепь и молекулу гема.

Q-цикл n Ko. Q переносит е- от комплекса I к комплексу III и Н+ в межмембранное пространство, совершая своеобразные циклические превращения — Q-цикл. n Донором е- для комплекса III служит Ko. QН 2, а акцептором — цитохром С. n В мембране — стационарный общий фонд Q/QH 2, из которого каждая молекула QH 2 в одном цикле обеспечивает перенос Н+ из матрикса в межмембранное пространство и е-, которые в конечном итоге поступают на О 2.

Цитохромы n ЦТХ ЦПЭ обозначают буквами b, с1, с, а и а 3, расположены в ЦПЭ в порядке увеличения редокс-потенциала. ЦТХ - гемопротеины. Атом Fe в геме ЦТХ меняет валентность, присоединяя и отдавая е-. Fe 2+ ↔ Fe 3+ + е. Комплекс III, включающий ЦТХ b и с1, функционирует как Ko. QН 2 - дегидрогеназа, осуществляет перенос е- с QH 2 на цит. С. епоследовательно проходят через атомы Fe ЦТХ b и с1, а затем поступают на цит. С. С Ko. QН 2 передаются 2 е- , т. к. ЦТХ переносят за 1 цикл по 1 е- , то перед символом ЦТХ ставится коэффициент 2. n
, включает 2 ЦТХ (а-а 3), содержит ионы")
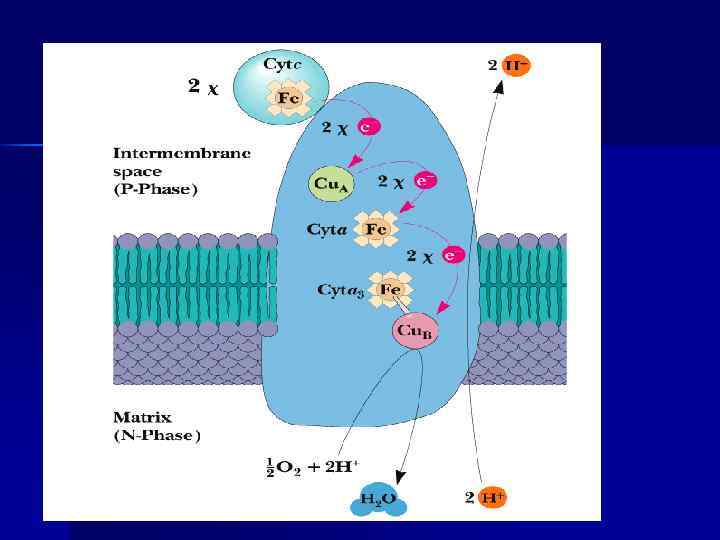
n IV-комплекс ЦТХ-оксидаза (цтх с-редуктаза -IV комплекс), включает 2 ЦТХ (а-а 3), содержит ионы меди. Перенос е- комплексом а-а 3 включает р-ции : Си+ ↔ Си 2+ + е. Fe 2+ ↔ Fe 3+ + еn Переносит е- с цит. С на О 2. Каждый из атомов О 2 последовательно присоединяет по 2 е- и 2 Н+, превращаясь в Н 2 О. n О 2 + 4 е- + 4 Н+ 2 Н 2 О

Комплекс IV Cytochrome Oxidase c

Комплексы I, III, IV
. Ингибитор QH")
Ингибиторы дыхательной цепи n n Ингибиторы NADH-дегидрогеназы: ротенон, амитал (из группы барбитуратов). Ингибитор QH 2 -дегидрогеназы – антимицин А (антибиотик). n Ингибиторы ЦТХ-оксидазы – цианид, СО, Н 2 S. Цианид присоединяется к Fе ЦТХ-оксидазы. n Переносчики цепи, стоящие в цепи непосредственно перед блокированным этапом, становятся более восстановленными, а стоящие после – более окисленными

Сопряженное окислительное фосфорилирование вдыхаемый О 2 - SH 2 ----2 Н+-- 2 е- + ½ О 2 S + Н 2 О (окисление, экзергонический процесс) S восст S окислен ↓ энергия---- тепловая энергия ↓(40%) АДФ + Фн АТФ + Н 2 О (фосфорилирование, эндергонический процесс)

Окислительное фосфорилирование n n n NADH+H++1/2 О 2 NAD++ Н 2 О+ 52, 6 ккал/моль (220 к. Дж/моль)— экзергонический процесс. АДФ + Н 3 РО 4 + 7, 3 ккал/моль (30, 5 к. Дж/моль) АТФ + Н 2 О — эндергонический процесс. Синтез АТФ из АДФ и Н 3 РО 4 за счет энергии переноса е- по ЦПЭ — окислительное фосфорилирование.

Векторное действие Е ЦПЭ n Е ЦПЭ фиксированы в МТХ мембране так, что их действие векторно, т. е. характеризуется не только величиной скорости реакции, но и пространственной направленностью. Основным проявлением векторности в ЦПЭ является перенос Н+ с внутренней стороны мембраны (со стороны матрикса) на наружную (в межмембранное пространство).

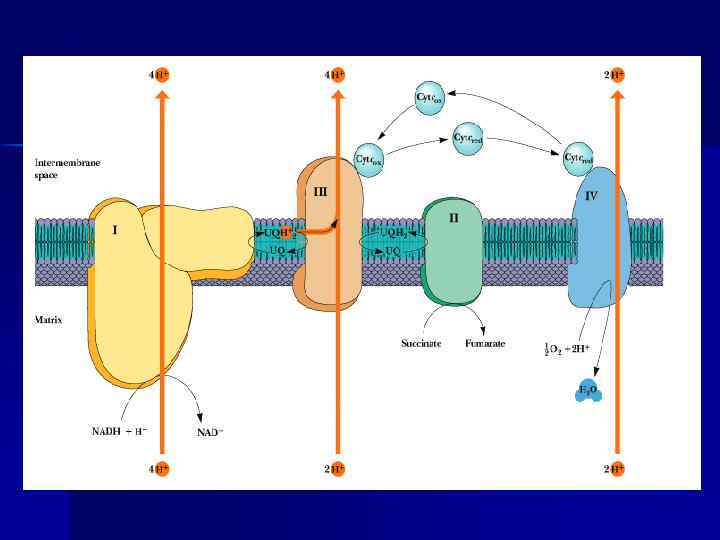
Пункты фосфорилирования n. В дыхательной цепи есть 3 пункта, связанные с перекачкой протонов: комплексы I, III, IV — пункты сопряжения или фосфорилирования.
 во внутренней")
АТФ-синтаза n n Переносчики е- организованы в 3 комплекса (I, III, IV) во внутренней мембране МТХ. Эти комплексы, используя энергию е-, обеспечивают перенос Н+ из матрикса в межмембранное пространство (векторное действие). В результате возникает протонный электрохимический потенциал (μН+). При достижении определенного значения ΔμН+ происходит активация АТФ-синтазы (комплекс V), в ней открывается канал, через который Н+ возвращаются в матрикс из межмембранного пространства, а энергия ΔμН+ используется для синтеза АТФ. Каждый из 3 комплексов ЦПЭ (I, III, IV) обеспечивает необходимый градиент Н+ для активации АТФ-синтазы и синтеза 1 молекулы АТФ. Образующаяся АТФ при участии АДФ/АТФтранслоказы из матрикса ЦП. Та же транслоказа переносит АДФ из ЦП в матрикс МТХ.

Механизм сопряжения. Протонный градиент и электрохимический потенциал. Наиболее активный транспорт H + — на участках, соответствующих I, III и IV комплексам — пункты сопряжения или фосфорилирования Электрохимический и концентрационный градиенты — электрохимический потенциал. n ΔμН+ — источник энергии для синтеза АТФ. n

Н+-АТФ-синтетаза n Электрохимический потенциал вынуждает Н+ двигаться в обратном направлении — с наружной поверхности внутрь. Однако мембрана непроницаема для них, за исключением участков, где располагается Е Н+-АТФсинтетаза, катализирующую р-цию: АДФ + Н 3 РО 4 АТФ + Н 2 О. n

ΔμН n На работу, совершаемую при выкачивании Н+, расходуется часть свободной энергии, которая освобождается при переносе е- по градиенту редокс-потенциала. n Энергия электрохимического потенциала (ΔμН+) используется для синтеза АТФ, если Н+ возвращается в матрикс через ионные каналы АТФ-синтетазы.
. n n n Перенос электронов по дыхательной цепи")
Хемиосмотическая теория П. Митчела (Нобелевская премия). n n n Перенос электронов по дыхательной цепи от НАД. Н+Н+ к кислороду сопровождается выкачиванием Н+ из матрикса митохондрий через внутреннюю мембрану в межмембранное пространство. Протоны, перенесенные из матрикса в межмембранное пространство, не могут вернуться обратно в матрикс, так как внутренняя мембрана непроницаема для протонов. Создается протонный градиент, при котором концентрация протонов в межмембранном пространстве больше, а р. Н меньше, чем в матриксе.

Хемиосмотическая теория П. Митчела Каждый протон несет положительный заряд, и вследствие этого появляется разность потенциалов по обе стороны мембраны: отрицательный заряд на внутренней стороне и положительный на внешней. n В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал Н+– источник энергии для синтеза АТФ n

Гипотеза Митчелла

Протонный градиент и АТФ-синтаза

Н+-АТФ-синтетаза n n n Н+-АТФ-синтетаза — интегральный белок внутренней мембраны МТХ, Олигомерный белок, состоит из 2 х комплексов — F 0 и F 1. Гидрофобный комплекс F 0 (трансмембранная часть) погружен в мембрану, состоит из нескольких субъединиц, образующих гидрофильный канал, по которому Н+ переносятся в матрикс. F 1 -субъединица (9 субъединиц) содержит активные центры Е Н+-АТФ-синтетазы, синтезирующей АТФ. ΔμН+ заставляет Н+ двигаться по каналу АТФсинтетазы в матрикс. Под действием ΔμН+происходят конформационные изменения в субъединицах и активация АТФсинтетазы, в результате чего синтезируется АТФ.

АТФ-синтаза

АТФ-синтаза

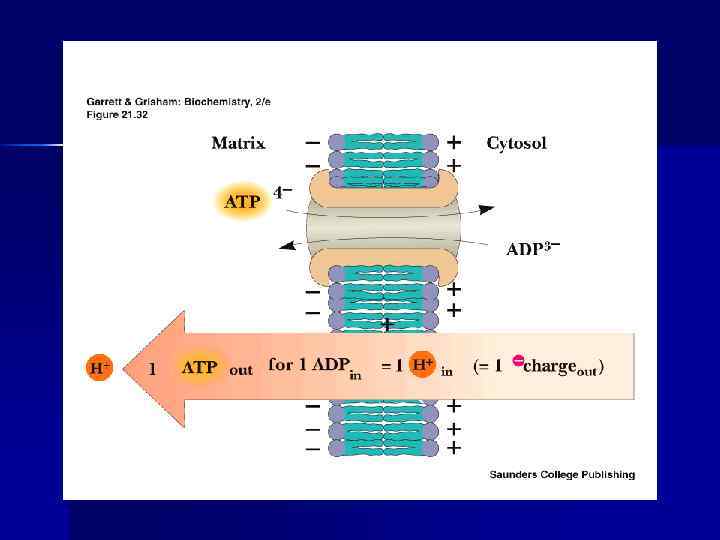
АТФ-АДФ-транслоказа n n n В большинстве клеток синтез АТФ происходит в митохондриях, а основные потребители АТФ расположены вне её; В мембране есть белок АТФ/АДФ-антипортер (транслоказа), осуществляющий перенос этих метаболитов через мембрану; Молекула АДФ поступает в митохондриальный матрикс только при условии выхода молекулы АДФ из матрикса; Движущая сила такого обмена - мембранный потенциал переноса электронов по ЦПЭ; На транспорт АТФ и АДФ расходуется около четверти свободной энергии протонного потенциала. Т. О. образующаяся АТФ при участии АДФ/АТФтранслоказы из матрикса ЦП. Та же транслоказа переносит АДФ из ЦП в матрикс МТХ.

Р/О n n В расчете на каждый атом поглощенного О 2 МТХ образуют 3 молекулы АТФ, т. е. 3 молекулы Н 3 РО 4 связываются с АДФ. Отношение количества связанной Н 3 РО 4 к количеству поглощенного О называют коэффициентом фосфорилирования, обозначают Р/О. ФАД-зависимые-дегидрогеназы не являются протонными насосами: в этом случае в ЦПЭ действуют только два пункта перекачки протонов — комплексы III и IV и коэффициент Р/О не может быть >2. Т. о. , Р/О = 3 или<3.

Дыхательный контроль n n n Результат окисления НАДН и фосфорилирования АДФ в ЦПЭ можно представить так: НАДН+Н++ ½ О 2 НАД + Н 2 О 3 АДФ + 3 Н 3 РО 4 3 АТФ + 3 Н 2 О n Зависимость дыхания МТХ от концентрации АДФ — дыхательный контроль. n Скорость синтеза АТФ определяется потребностью клетки в энергии. При увеличении расходования АТФ, увеличивается концентрация АДФ, а это ведет к ускорению дыхания и фосфорилирования.

Разобщение дыхания и фофорилирования n Разобщение потока электронов и окислительного фосфорилирования происходит при нарушении протонного градиента; Разобщители являются гидрофобными (липофильными) молекулами, способными диссоциировать и проходить через липофильный слой мембраны. n Они перемещаются через мембрану, перенося протоны из межмембранного пространства обратно в матрикс, нарушая протонный градиент. n
 диффундирует через")
Разобщение дыхания и фофорилирования n n n 2, 4 динитрофенол (липофильное вешество) диффундирует через МТХ мембрану как в ионизированной, так и в неионизированной форме и поэтому может переносить Н+ через мембрану в сторону их меньшей концентрации в обход протонных каналов. 2, 4 динитрофенол уничтожает ΔμН+ МТХ мембраны и энергия рассеивается в виде тепла (разобщители обладают пирогенным действием. ). Потребление О 2 и окисление S при этом происходят, но синтез АТФ не возможен. Разобщители: дикумарол, билирубин, тироксин в высоких концентрациях, ВЖК.

Терморегуляторная функция – Разобщающий белок – термогенин способен переносить анионы ЖК, служащие разобщителями.
Bioenergetika_I_S_Grig_1l_II.ppt