Методы определения мутагенной активности химКаз Бакалавриат.ppt
- Количество слайдов: 32



Бактериялардың ДНҚ репарациясының SOS-жүйесі Қоршаған орта факторларының мутагенділігін зерттеу әдістері
 ашқан Escherichia coli бактериясындағы SOS-жауап УФсәулелену, көміртегі тотығы,")
• Мирослав Радман (Radman, 1974) ашқан Escherichia coli бактериясындағы SOS-жауап УФсәулелену, көміртегі тотығы, митомицин, блеомицин және т. б агенттердің ДНҚ-ны зақымдау кезінде пайда болатын шамамен 40 геннің координирленген индукциясы болып табылады.
")
• SOS- жүйенің пайда болуы туралы гипотеза келесі мәліметтер негізінде қаланған: • 1) сәулеленген фагтар бұрынғы сәулеленген E. Coli клеткасымен культивирленген кезде, УФ сәулеленген λ фагтардың әсерім (реактивациясы) біршама артатынын бақылау; • 2) λ профагасының және лизис бактерияларының УФ сәулеленуі кезіндегі индукциясы. • 3) E. coli клеткасының УФ-сәулеленуге жауап беру филаментациясы. • Осы аталғандардың барлығы клеткалық бөлінудің кешігуі, λ профагы индукциясы механизмі және УФсәулеленуге ұшыраған индуцирленген мутациялар арасындағы байланыстың бар екендігі деген ой туғызды.

• SOS- жүйесінің жұмыс жасау кезінде клеткалардың репарациясына және ДНҚ репликациясына қабілеттігі артады, олардың зақымдануы мен мутагенездің интенсивтілігіне төзімділігі артады. • SOS-жауаптың индукциясы кезінде маңызды ролді lex. A және rec. A гендерінің өнімдері атқарады, сондай-ақ, ДНҚ-дағы біржіпшелі жыртылулардың (разрыв) болуы (ss. DNA) маңызды роль атқарады. • lex. A гені Lex. A белогын кодтайды. Ол белок SOS-регулон гендерінің репрессоры болып табылады. • Ол SOS-бокс деп аталатын ДНҚ-ның арнайы кезектілігімен тығыз байланысты. Ол бокс SOS-гендердің промоторлық аудандарында орналасады және ол қалыпты физиологиялық жағдайлар кезінде SOS- жүйесінің әлсіз базальды экспрессиясын анықтайды • ДНҚ-ның зақымдануына жауап ретінде rec. A генімен кодталатын Rec. A реттегіш ақуызы юелсенеді.

SOS- жауап индукциясы

• SOS мутагенездің болуы маңызды биологиялық мәнге ие. • Прокариоттардың тіршілік ету қағидасындағы маңызды факторлардың бірі болып жоғары өзгергіштік табылады. • Мүмкін, осыбактерияларда жаңа мутациялардың пайда болуындағы роль SOS-жүйеге тән.

• Қазіргі кезде мутагенділікті бағалау мен анықтаудың 100 -ден астам әдістері бар. • Индикатор ретінде микроорганизмдер, сүтқоректілер мен in vitro адамның клеткасы, өсімдіктер, жануарлар алынады.

• Клеткалы макромолекулалы мутагендердің молекулалық дозиметриясы үшін ( мутагендер кешендерін-ДНҚ-ны, мутагенбелок немесе алкилді қосылыстарды) алуантүрлі биохимиялық, иммунохимиялық әдәстер мен моноклоналды антидене қолданылады. • Мутацияның барлық түрлерін анықтауға арналған универсалды әдіс жоқ. Осыған байланысты мутагенділікті зерттеу үшін мутацияның әртүрлі санаттарының индукциясын тіркеуге мүмкіндік беретін бірнеше әдістер қолданылады.

• Сынақтың сатылы жүйесі барлық химиялық генетикалық қауіпті заттарды скринингтің қарапайым немесе жылдам әдістерінің (егу) көмегімен анықтауға болатындығына негізделеді. • Скринингті тест- жүйелер қатарына мутагенділік индикаторы ретінде микроорганизмдер пайдаланылатын әдістер жатады. • Скрининг барысында табылған мутагендер in vitro және in vivo сүтқоректілерінің клеткасында генетикалық бұзылулардың индукциясын ескеруге мүмкіндік беретін тест жүйелердегі жан жақты зерттеулерге әкеледі.

• Эймс тесті әртүрлі елдердегі скринингтік бағдарламалардағы басты әдістердің бірі болып табылады, және ол қосылыстардың көп мөлшерін салыстырмалы түрде тез болжауға, сонымен қатар олардың арасынан адам үшін мутаген болып табылатынын іріктеуге мүмкіндік береді.
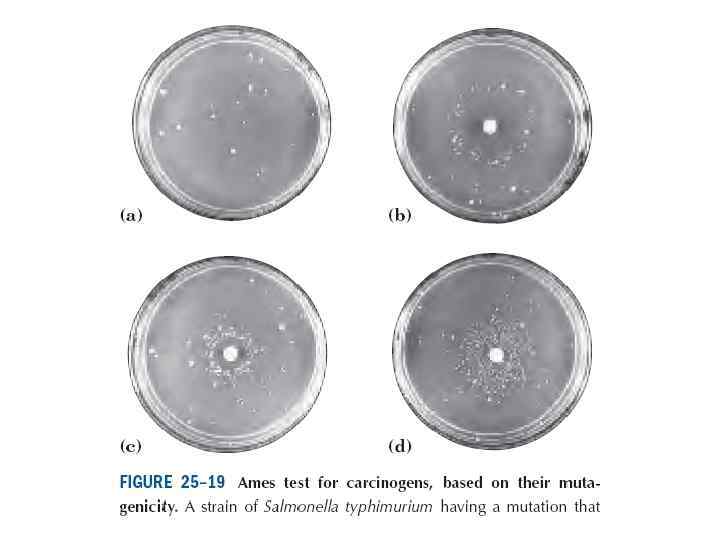

• Эймс тесті. Әдістің басты қағидасы мутантты бактерия жасушалары, ауксотрофтарды зерттелетін заттың қатысуымен өсіруге негізделген. • Ауксотрофты деп мутация нәтижесінде биосинтезі бұзылған метаболиттердің қосылуынсыз аз қоректік ортада өсуге қабілетсіз бактериалық немесе басқа жасушаларды атаймыз. • Эймстың классикалық тестінде көбінесе Salmonella typhimurium жасушасын қолданады, олар амин қышқылы бойынша ауксотрофты, олардың ауксотрофтылығы жалғыз нүктелік мутацияның нәтижесінде туындайды.

• Бактерия суспензиясын зерттелетін заттың қатысуымен инкубациялайды және қатты ортаға отырғызады, ол қоректік ортаның құрамында бактериялары ауксотрофты болатын заттардың аз мөлшері бар. • Бұл заттар көрінетін колония түзбей жасушалар бірнеше бөлінуді іске асыру үшін жеткілікті болуы керек. • Соған қарамастан түзілген мутация бактерия геномында фиксацияланады. Локуста реверсия туындаған кезде бұндай бактериялар қатты қоректік ортада көрінетін колониялар түзе отырып, жоғарыда атылған заттың қатысуынсыз өсуге қабілетті болады. • Осылай егер сыналатын зат мутагендік әсерге ие болса, ревертанттардың түзілу әсерлігі заттың қатысуымен жоғары болады, ол көптеген бактериялық колониясы түрінде көрінеді және тағамдық қоспаның қатысуынсыз қатты қоректік ортада өседі.

• Эймс тестінің модификациясының көптеген түрі бар. • Маңыздырағы болып промутагендердің белсенуі үшін кеміргіштердің бауырының микросомалық фракцияларын пайдалану табылады. • Бауырдың микросомалық фракцияларының құрамында ксенобиотиктердің метаболиттік белсенуінің барлық негізгі ферменттері бар, сол үшін сыналатын зат промутагендік мутацияға ие болса, ол in vitro микросомасының ферменттерімен белсенеді және бұндай нәтиже Эймс тестінде оңай анықталады.

• SOS-хромотесті • Бұл тест мутациялық процестің механизмі туралы заманауи көріністердің негізінде қаланған • E. coli жасушасында SOS-жауап өзіне ДНҚ-ның зақымдалуы немесе оның синтездерінің тоқталуына жауап ретінде индуцирленетін қызметтерді біріктіреді. • Сондай sfi. A генімен басқарылатын. SOS-қызметтердің бірі клеткалық бөліну мен жіпше тәрізді өсудің ингибирленуінде көрініс табады. • В клетках тестерного штамма E. coli p. Q 37 под контроль промотора гена sfi. A введен структурный ген – β-галактозидазы (Quillardet P. et. al. , 1982). • Осылай, β-галактозидазасының белсенділігі sfi. A генінің экспрессиясының дәрежесіне бірден бір тәуелді болады және SOS-қызметтің индукциясының көрсеткіші болып табылады.

• Бар мәліметтердің қорытындысы бойынша, SOSхромотест келесі аспектілер бойынша мутагенезге Эймс тестін толтыра алады: • - сұйық ортаның активациясы барысында мутагенділік көрсететін канцерогендерді табу; • -басткриялар үшін улы қосылыстарды орнату, яғни ісікке қарсы блеомицин препараты, тек қана Эймс тестінде S. typhimurium TA 102 штамындағы мутаген ретінде идентифицирленетін неокарциностатин және митомицин С препараттары. • -ДНҚ синтезінің антиметаболитті ингибиторлерін анықтау үшін.

• SOS-хромотесттің басты артықшылығы болып оның қарапайымдылығы табылады. • Тек ғана бір штамм қолданылады. • Бұл өте маңызды, себебі штаммдардың санының артуы жұмысты қиындатады және қымбаттатады. • Сандық колориметрлік жауап бірнеше сағат аралығнда алынуы мүмкін. • SOS-хромотестінің негізгі кемшілігі болып ДНҚ-дағы интеркалирлеуші агенттердің белсенділігін анықтамайтындығы табылады.

• Цитогенетикалық әдістер • Хромосомалық аберрацияны есептеу • Адам мен жануардың соматикалық жасушасында туындаған барлық хромосомалық аберрациялар басты екі топқа бөлінеді: хромосомалық және хроматидалық. • Екі аберрацияны хромосомалық немесе хроматидалыққа жатқызу хромосоманың қандай дәрежеде зақымданғандығына байланысты болады.
, ол")
• Хромосомалық типтегі аберрация предсинтетикалық кезеңдегі хромосоманың зақымдануын көрсетеді (G 1 -фаза), ол бір тізбекті боладығанда хроматиндік типтегі аберрация хромосоманың екі тізбекті кезеңде зақымданған кезінде туындайды, яғни S және G 2 фазасында. • Сирегірек хромосоманың қом хроматидасының зақымданғаны кезінде ұқсас локустарда изохроматидалық үзінділер туындайды, олар хроматидалық типтегі аберрациядан морфологиялық жағы бойынша ұқсас. • Аберрациялар қарапайым және ауыспалы болуы мүмкін. • Бос немесе онымен бірге фрагменттердің түзілуімен бір немесе бірнеше хромосоманың бұзылуын аберрацияның қарапайым типіне жатқызады, ал басқа немесе сол хромосоманың участоктарының қайта құрылуы немесе бірнеше хромосоманың арасында участоктармен қайта комбинациялануы ауыспалы типке жатады.

• Ауыспалы аберрация хромосома ішілік және хромосома аралық болуы мүмкін. • Локализацияғға байланысты хромосома ішілік аберрацияның өзі иық ішілік және иық аралық деп бөлінеді. • Хромосомалық алмасу кезінде егер ацентрлік фрагменттер центрлікпен байланысатын болса, онда алмасу симметриялық деп, ал центрлік фрагменттердің бірігуі кезінде асимметриялық деп жіктелінеді. • Соңғы жағдайда полицентрлік хромосома мен хроматида пайда болады. • Хромосома ішілік және хромосома аралық алмасулар толық және толық емес болуы мүмкін. Птолық алмасу кезінде зақымданған хромосоманың барлық қайта комбинацияланған участоктарының қайта қосылуы жүреді.

• Адамдағы химиялық және радиациялық мутагенезді зерттеу үшін кеңінен қолданылады. • Адам лейкоцитінің культурасының артықшылығына материалдың қолжетімділігі, жасушалық популяциның синхрондылығы, спонтпндық мутацияның төмен деңгейі, хромосома морфологиясының зерттелгендігі жатады.

• Перифериялық қанның лимфоциттерін культивирлеу арқылы цитогенетикалфы препараттарды дайындаудың әдістері.

• Химиялық қосылыстарды мутагендік белсенділікке тестілеу кезінде хромосомалық аберрацияны кариотиптеусіз есептейді. • Метафазалық пластинканы кариотиптеу арнайы зерттеулерде қолданылады, мысалы хромосома тобы бойынша зақымданудың таралуын зерттеу кезінде.

• Қалыпты жағдайда спонтандық деңгейден асып түсетін хромосомалық аберрацияның жиілігі бойынша мутагендік белсенділік туралы талқылайды. • Химиялық мутагенмен индуцирленген аберрация спонтанды аберрацияға аналогты.

• Химиялық қосылыстарды тестілеу кезінде тәжірибені екі кезеңнен өткізеді. • Бірінші кезеңде максималды концентрацияда заттың мүмкін болатын әсерін табуға негізделген. Культураны өңдеу 48 сағатқа созылады және жасушалар 72 сағатта фиксацияланады. • Әсердің пайда болуы кезінде доза әсердің тәуелділігін зерттеуге өтеді. • (2 -ші кезең).

• Қарапайым бояу кезіндегі пластинка

• Радиациялық индукцияланған хромосомалық аберрациясы бар метафазалық пластинка Химиялық индукцияланған аберрацияға ие метафазалық пластинка
. • in vivo химиялық заттардың генотоксиндік әсері геномның дестабилизациясы")
• Туыстас хроматидалық алмасулар(ТХА). • in vivo химиялық заттардың генотоксиндік әсері геномның дестабилизациясы және оның қайта құрылуымен жүзеге асады. • Егер мутагенмен байланыс ДНҚ үзілістерінің түзіліне әкеп соқса, олардың репарациясы кезінде интерфазалық ядрода туыс хроматидалардың гомологиялық участоктермен алмасуы байқалады.

• Өсімдік пен жануардың соматикалыжасушасынан ТХА-ны табуға арналған әдістер құрастырылды. Бұл жасушалар in vitro 5 бромдезоксиуридиннің қатысуымен инкубацияланады ол кезде тимидиннің орынына олардың ДНҚ-сына енеді. • Хромосомадағы хроматиданың бірі ДНҚ-ның қос тізбегіне нуклеозидтің аналогын қосады. • Флуоресцентті бояғыш әрқалай осындай гомологиялық хроматидалармен әсерлеседі, ол қарапайым микроскоптың көмегімен боялудың интенсивтілігі бойынша анықталады, – гомологиялық хроматидның бірі басқасына қарағанда ашық болып келеді. • ТХА-ды тапқан кезде жеке хромосоманың хроматидасында ашық жәнее қара участоктар кезектеседі және осындай участоктардың саны ТХА-ның жиілігінің өлшемі болып табылады. ТХА-ы жоғары болған сайын соматикалық жасушаға мутагендік әсер ету интенсивті бола түседі.

• Туыс хроматиданың алмасуы

• Эймс тестінен айырмашылығы ТХА-ды анықтау адам ағзасына және жануарға мутагендік әсерді ретроспективті бағалау үшін пайдаланылуы мүмкін, себебі ТХА болмағаны кезінде мутагеннің иницирлеуші әсерінен кейін жалғасады, мысалы иондаушы сәулеленудің көзін жойғаннан соң немесе ксенобиотиктің толық детоксикациясын жойған соң.

Назар аударғандарыңызға рахмет!
Методы определения мутагенной активности химКаз Бакалавриат.ppt