
Антитіла ( antibody = AB ). Будова

. Будова і властивості. Гени імуноглобулінів. Біосинтез антитіл.")










")




 і константні ( С- constant )")










 може набувати")













")





 продемонстровано активний центр у")
 універсальність: антитіла можуть бути виробленими проти")

 ге нів імуноглобулінів: Н, κ")
 Локус генів імуноглобулінів")

 генів імуноглобулінів 1) сайт-специфічна рекомбінація V J ( для генів")




 множинність гаметних")





 генів V- області імуноглобулінів")












")

 = ( diversification ) імуноглобулінів Комбінаторна диверсифікація (див. табл. кластерів")
 , урацил-ДНК-глікозилаза ( UNG))")




 мають колір,")
")





")


lekcii_4_5._immunoglobul__2015.ppt
- Размер: 46.6 Мб
- Автор:
- Количество слайдов: 122
Описание презентации Антитіла ( antibody = AB ). Будова по слайдам
Антитіла ( antibody = AB ). Будова і властивості. Гени імуноглобулінів. Біосинтез антитіл.
Тема 4. Антитіла: будова і властивості
Основні функції антитіл
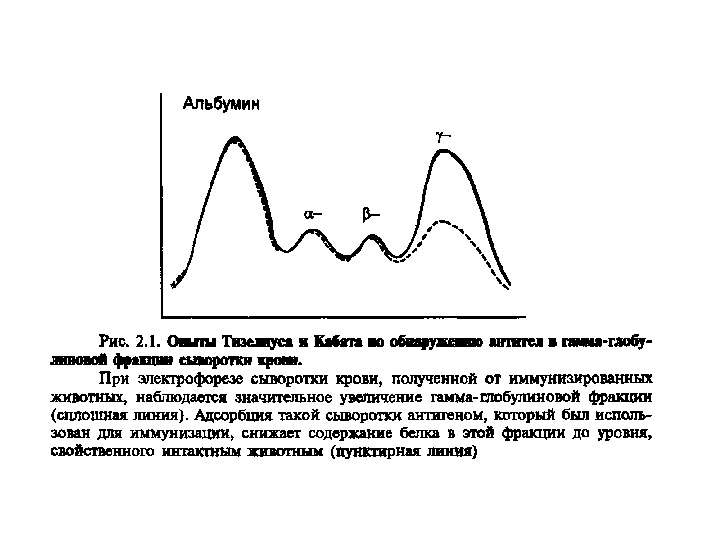
Антитіла – продуковані В- лімфоцитами глікопротеїни, відкриті як γ — глобулінова фракція сироватки крові, що здатні специфічно взаємодіяти з антигеном. В 1972 році Родні Портер та Джеральд Едельман отримали Нобелівську премію за вивчення структури імуноглобулінів. Згідно з їхніми дослідженнями було створено модель молекули антитіла
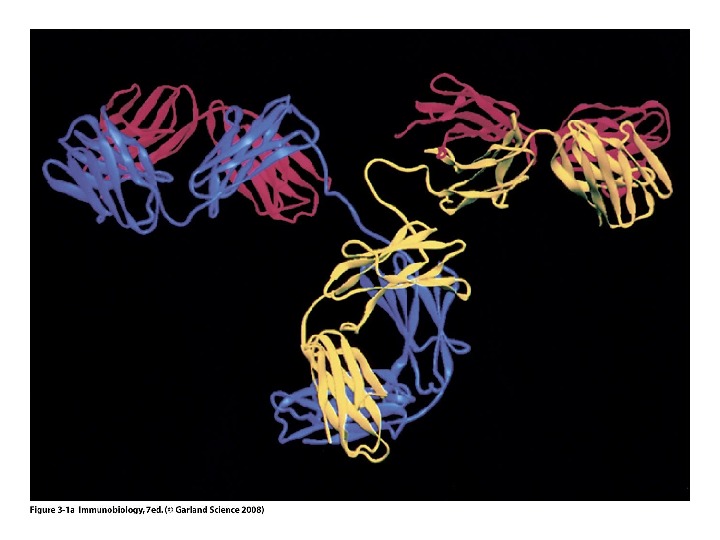
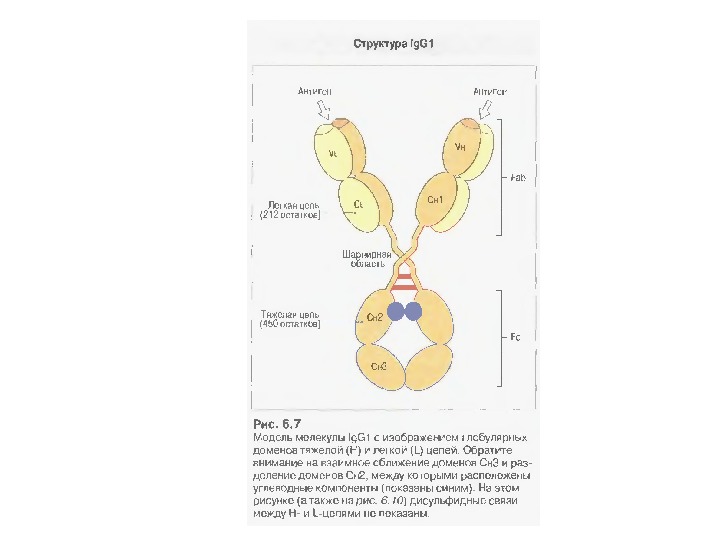
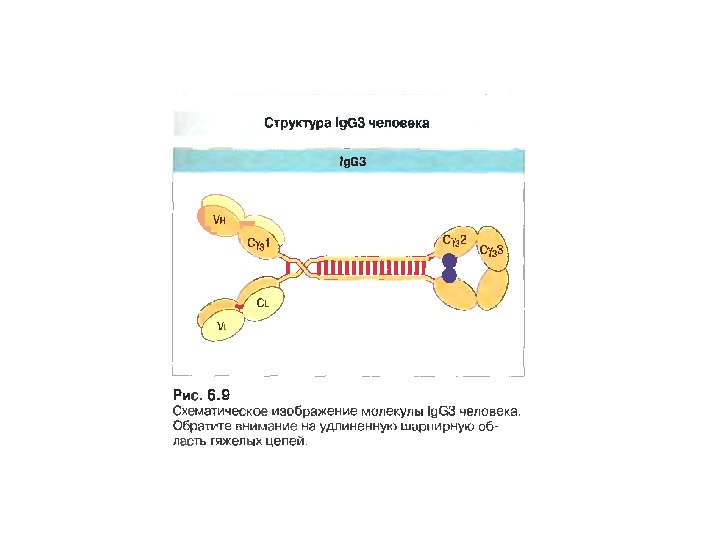
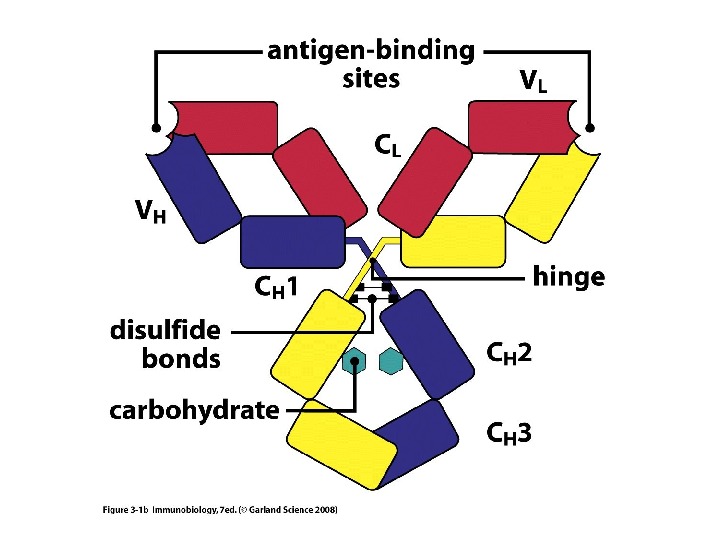
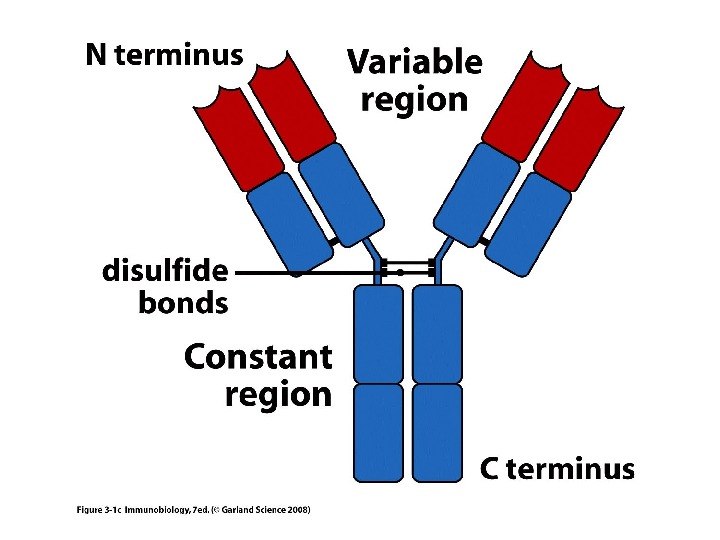
Будова антит і ла Структура Ig. G
Будова антит і л
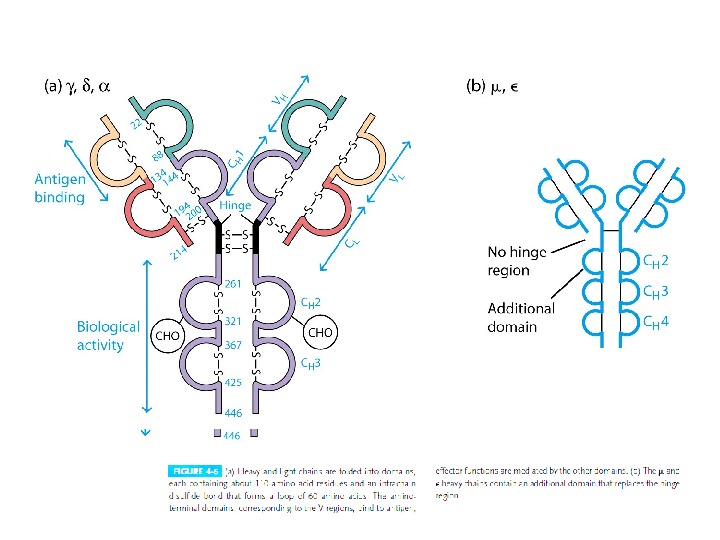
Структура імуноглобулінів Н-важкі поліпептидні ланцюги 5 класів ланцюгів — відповідні класи ( ізотипи) імуноглобулінів α -Ig A підкласи α 1, α 2 γ — Ig G підкласи γ 1, γ 2, γ 3, γ 4 δ — Ig D ε — Ig E μ — Ig M L- легк і поліпептидні ланцюги 2 типи — κ , λ Підтипи 1 κ і 4 λ
Доменна організація імуноглобулінів
Доменна організація імуноглобулінів
Доменна організація імуноглобулінів
Доменна організація імуноглобулінів
Доменна організація імуноглобулінів (всередині-цілісна молекула білка, зліва-константні домени, справа – варіабельні домени)
Типова молекула імуноглобуліну. Структурна організація легкого ланцюга імуноглобуліну
Типова молекула імуноглобуліну. Виділено структурну організацію легкого ланцюга імуноглобуліну
Надродина імуноглобуліноподібних молекул, яка нараховує у людини приблизно 765 членів
Імуноглобуліни — глікопротеїни, містять до 12% вуглеводневих залишків. Олігоцукор ( N-a цетилглюкозамін, манноза, галактоза), що зв ’ язаний N -глікозидним зв ’ язком із залишком аспарагіну у F с фрагменті Ig G
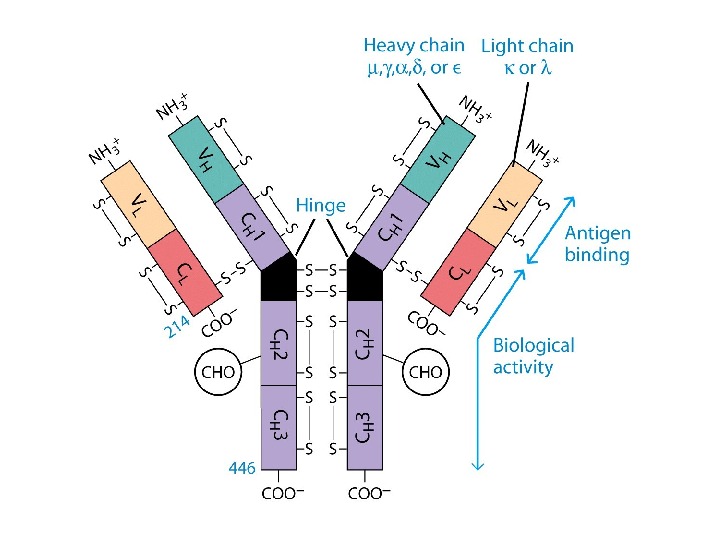
Будова імуноглобулінів: варіабільні ( V — variable ) і константні ( С- constant ) домени
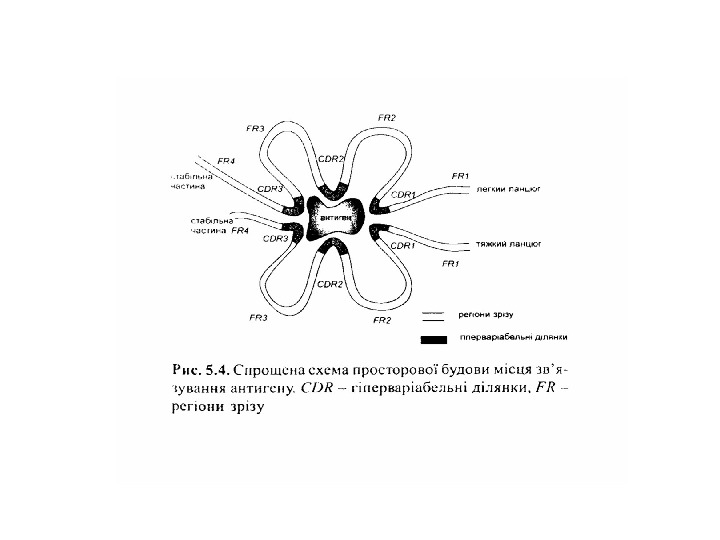
Будова імуноглобулінів HV — hypervariability regions — гіперваріабельні ділянки = CDR -complementarity — determining regions (більш загальний термін) ; FR — framework region — каркасні ділянки =регіони зрізу
Будова імуноглобулінів. Організація активного центру антитіл Організація V L – варіабельного домену легкого ланцюга, що структурує активний центр антитіла
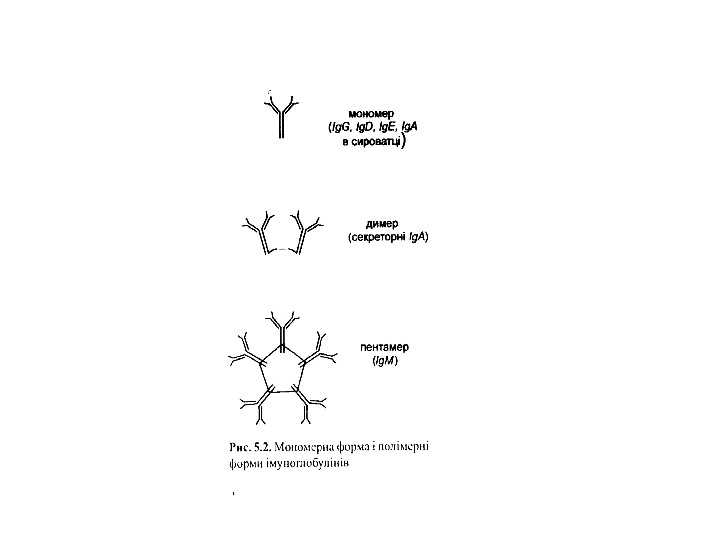
Четвертинний рівень організації імуноглобулінів
Четвертинний рівень організації імуноглобулінів
Протеолітична фрагментація молекули антитіла
Протеолітична фрагментація молекули антитіла
Імуноглобуліни можна розглядати як антигени. На них відрізняють чотири типи антигенних детермінант: • видові, • алотипічні , • ізотипічні , • ідіотипічні.
Функційний дуалізм описується гетерогенністю і варіабельністю. Гетерогенність визначає властивості І g , які зумовлені константною частиною молекули, що вирізрізняє роботу різних класів цих молекул. Варіабельність – це індивідуальна характеристика І g , що належать до одного класу або підкласу. Виявляється у специфічній антигензв ’ язуючій активності і характеризується варіабельними доменами молекул антитіл.
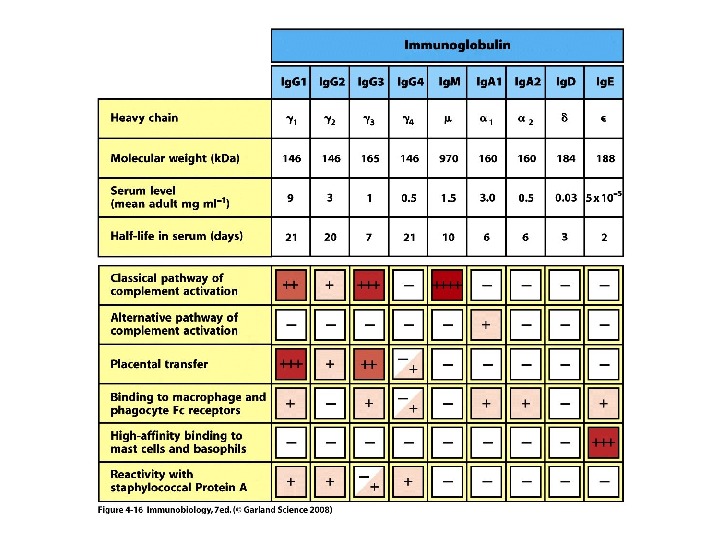
Класи імуноглобулінів
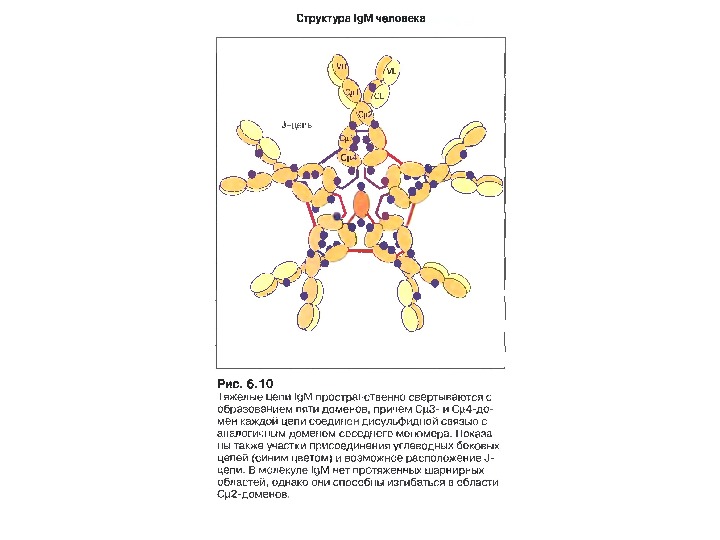
Ig. M 5 -10% сироваткових Ig , Мм мономерної форми- 190 к. Да, пентамерної форми — 970 к. Да
Форми Ig. M Вільний Ig. M ( не зв’язаний з антигеном) може набувати форми зірки, а Ig. M , зв’язаний з антигеном, що містить лінійно повторювані епітопи, набуває форми краба (на підставі мікрофотографій R. Feinstein і співавт. ).
Ig. M
Ig. G 70 -80% сироваткових І g, Мм в середньому 150 к. Да
Процес опсонізації
Процес опсонізації
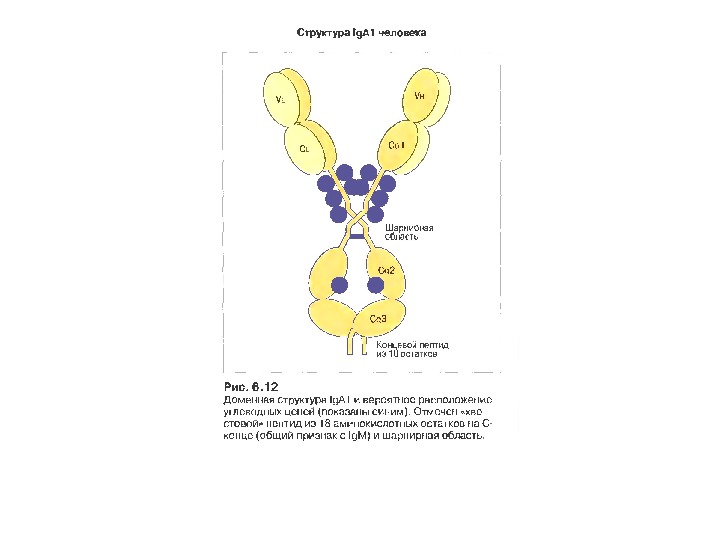
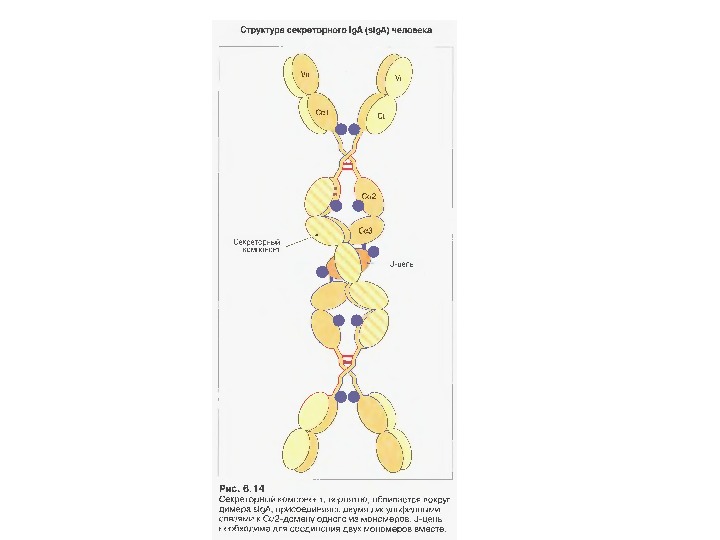
Ig А 13 -15 % сироваткових І g, Мм в середньому 150 к. Да
Механізм переносу s І g А через епітеліальну клітину
Механізм переносу s І g А через епітеліальну клітину
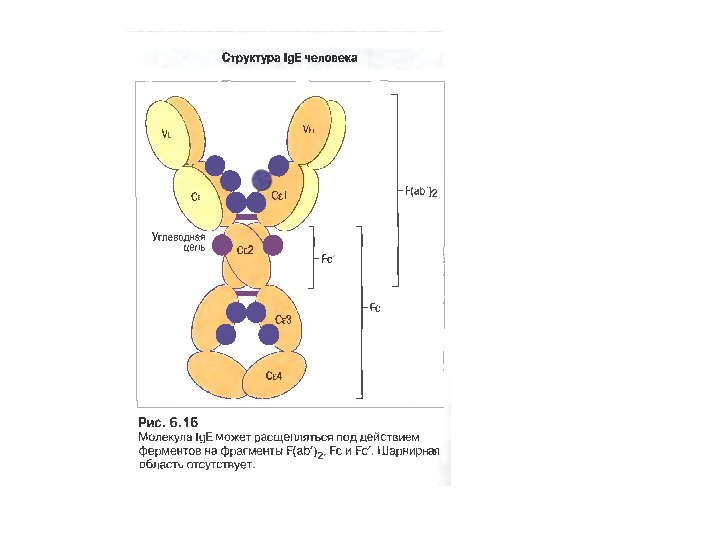
Ig Е 0, 002 % сироваткових І g, Мм в середньому 190 к. Да
Функціонування Ig.
Функціонування Ig.
І g. D 2 % сироваткових І g, Мм в середньому 185 к. Да
Ig
Середня концентрація у сироватці (мг/мл)
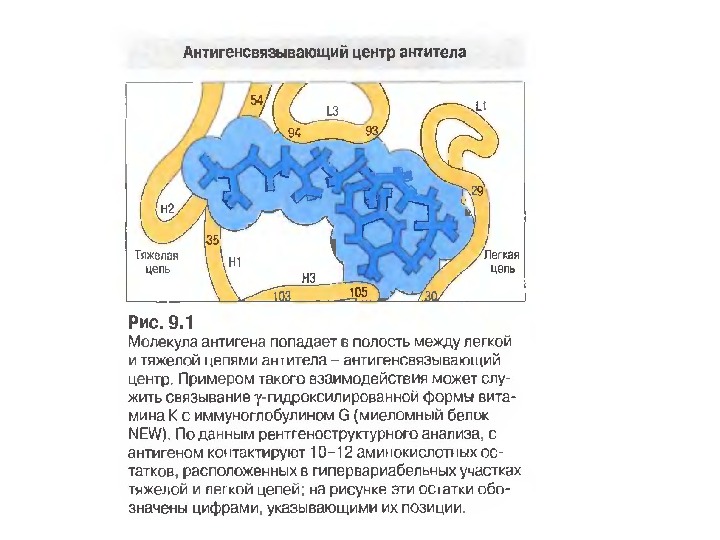
Будова активного центру антитіл 1. До складу активного центру антитіла входять гіперваріабільні амінокислотні залишки. 2. Зв’язок антигену з антитілом не ковалентний, а іонний, водневий, гідрофобний , Ван-дер-Ваальсівський. 3. Розміри активного центру, де може розміститися 5 -6 залишків амінокислот чи глюкози, становлять, відповідно, 1, 5 х0, 6 х0, 8 нм і можуть варіювати у різних антитіл. 4. Активний центр — це порожнина, заглиблена у Fv фрагмент приблизно на 1, 5 нм. 5. У 70 -ті роки 20 ст. було проведено рентгеноструктурний аналіз мієломних імуноглобулінів, який підтвердив дані, отримані раніше, і показав, що активний центр антитіла — це порожнина, в утворенні якої беруть участь гіперваріабільні ділянки F а b фрагменту, зближені у третинній структурі. За цими ж данними приблизно 10 -12 амінокислотних залишків гіперваріабільних ділянок важкого і легкого ланцюга беруть участь у контактуванні з антигеном.
Будова активного центру антитіл
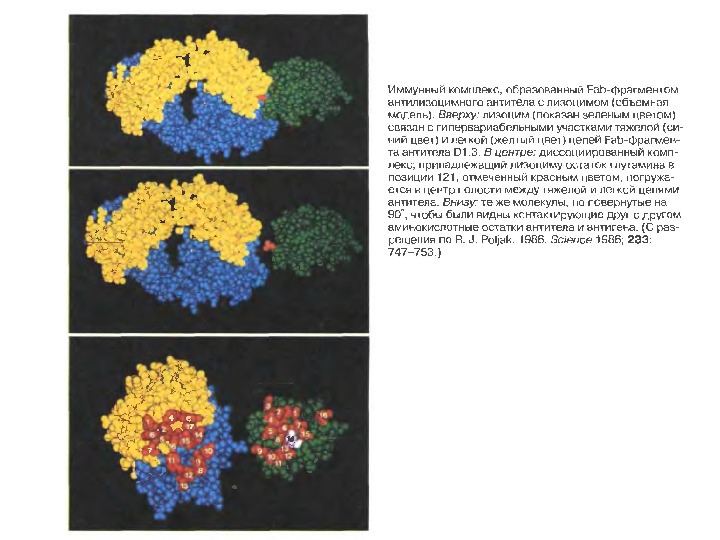
Будова активного центру антитіл Комплекс лізоциму курячого яйця з антитілом D 1. 3. Фіолетові- важкі ланцюги; легкі ланцюги- жовті ; лізоцим- блакитн ий. Червоним позначено залишок глутаміну 121, що належить лізоциму, який міститься між двома V — доменами і утворює водневі зв’язки, що стабілізують взаємодію АГ-АТ
Будова активного центру антитіл
Будова активного центру антитіл
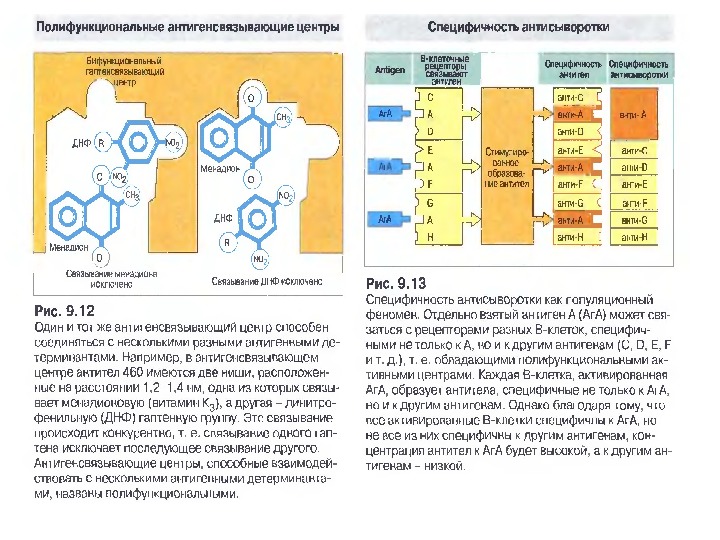
Будова активного центру антитіл • На рис. A ) продемонстровано активний центр у вигляді кишені ( pocket =жовтий), де зв’язується малий гаптен ферроцен (зелений). • B ) В антигензв’язуючому центрі знаходиться білковий антиген (вірусу HIV ), занурений в жолобок ( groove ) , утворений V -доменами важкого і легкого ланцюга. • С) активний центр у вигляді поверхні ( surface ) зв’язує білковий антиген –лізоцим курячого яйця з залученням всіх 6 CDR обох ланцюгів. • D ) Великий HIV антиген gp 120 утворює комплекс з антитілом у формі виступаючої поверхні ( protruding surface ) , а саме видовжена петля CDR 3 антитіла входить у заглибину в молекулі антигену.
Чотири риси гуморальної імунної відповіді 1) універсальність: антитіла можуть бути виробленими проти будь-якого хімічного угрупування. Важко уявити, скільки повинно бути варіантів антитіл, якщо вважати, що кожний антиген потребує окремого антитіла; 2) специфічність : антитіла розрізняють о- чи p-положення функціональних груп, стереоізомери, білки, що відрізняються на один амінокислотний залишок; 3) гетерогенність: на один антиген виробляється багато типів (популяцій) антитіл, інколи більше 100 на одну антигенну детермінанту; 4) поліфункціональність: одне антитіло здатне зв’язувати більш, ніж один антиген. Поліфункціональність антитіл сприяє універсальності імунної відповіді, а гетерогенність — її високій специфічності.
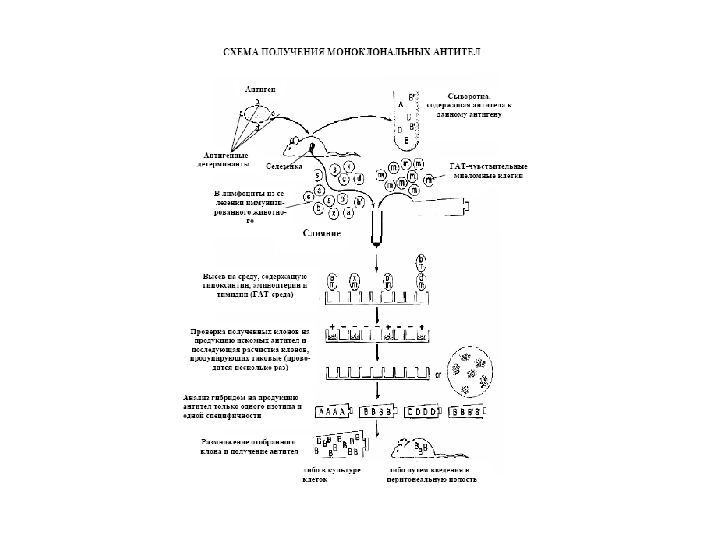
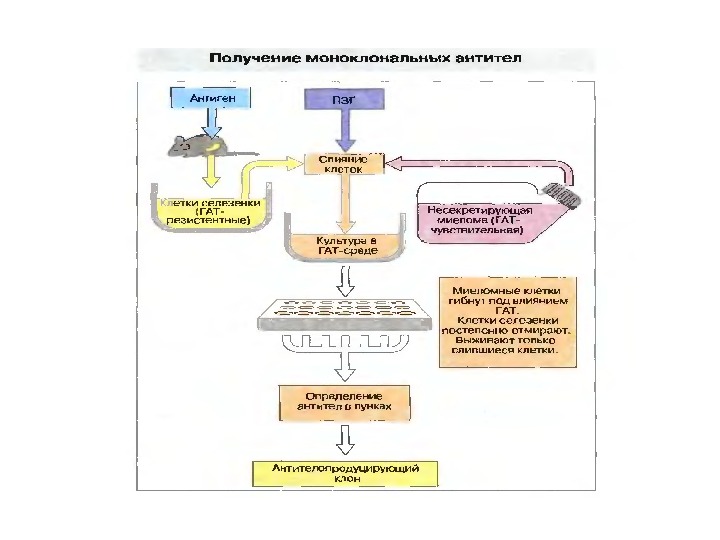
Тема 5. Ген и імуноглобулінів. Біосинтез антитіл 1976 р. С. Тонегава і співавт. довели можливість реаранжування генів 1987 — Нобелівська премія за вивчення генів імуноглобулінів
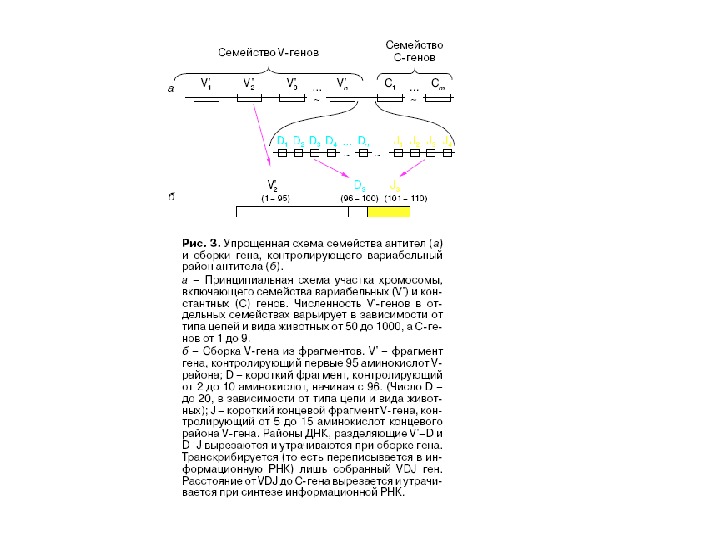
Ген и імуноглобулінів. Біосинтез антитіл Три кластери (локуси) ге нів імуноглобулінів: Н, κ , λ. У людини – Н- 14 хромосома, κ — 2 хромосома, λ — 22 хромосома. У миші — Н- 12 хромосома, κ — 6 хромосома, λ -16 хромосома. Входять V , C — гени важких і легких ланцюгів, J — гени ( joining ) , D -гени ( diversity ). 1. Кластер генів λ -ланцюга. 30 V генів, 4 (6) пар J λ -і С λ – генів, які є зчепленими між собою. 2. Кластер генів κ –ланцюга. 40 V генів, 5 J -генів, 1 С κ -ген. 3. Кластер генів Н-ланцюгів. Більше 1000 V — генів (функційні – 51), 27 D -генів, 6 J -генів та С-гени всіх класів імуноглобулінів. V — сегменту передує ділянка ДНК, що кодує лідерний пептид ( L).
Загальна характеристика хромосом людини ( http: //www. ensembl. org ) Локус генів імуноглобулінів людини: Н- 14 хромосома, κ — 2 хромосома, λ — 22 хромосома
Організація локусу генів зародкової лінії , що кодують імуноглобуліни людини
Реорганізація (реаранжування) генів імуноглобулінів 1) сайт-специфічна рекомбінація V J ( для генів легких ланцюгів)-одна рекомб і нац і йна подія; 2) сайт-специфічна рекомбінація DJ (1) і V + DJ (2)- для генів важкого ланцюга- дві рекомбінаційні події; 3) транскрипція цілого блоку V ( D ) J — intron — С; 4) сплайсинг РНК з утворенням функціонального транскрипту V ( D ) JC ; 5) трансляція; 6) посттрансляційне відщеплення лідерної послідовності та модифікації. (гени, що кодують варіабельні домени, утворюються в результаті сайт-специфічної рекомбінації — 1, 2)
Транскрипційний промотор активується тільки після проходження рекомбінації, тобто транскрипція неперебудованої ДНК є неможливою
Схема перебудови генів імуноглобулінів
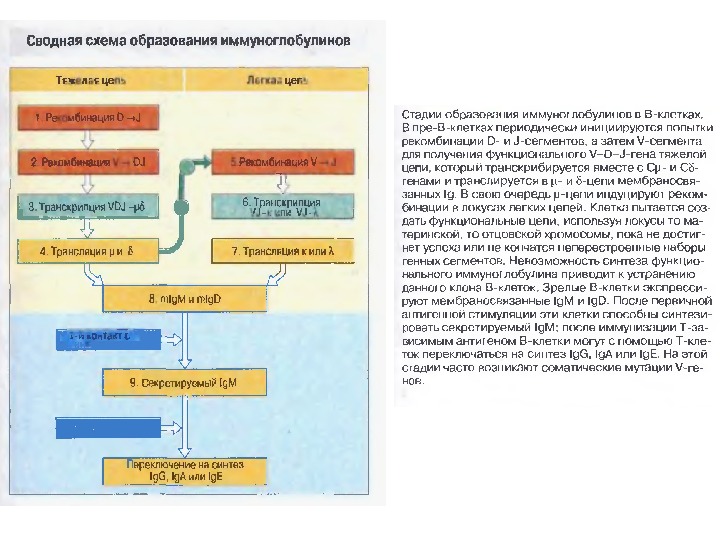
Стадії 0 A B C D E FСтадії дозрівання В-лімфоцитів (1)
0 -лімфоїдна стовбурова кровотворна клітина. А -ранні про-В-клітини — не мають рецепторів В клітин, започатковується реаранжування DJ — сегментів в локусі важкого (Н) ланцюга імуноглобулінів, може відбуватись в обох гомологічних хромосомах. В — пізні про-В-клітини — V — DJ реаранжування генів (підлягає алельному виключенню — тобто проходить лише в одній хромосомі). На цій стадії перебудовуються гени і “ експресується” важкий ланцюг в комплексі з сурогатним легким, що є сигналом для позитивної селекції цих клітин. С — великі пре-В-клітини — “експресують” пре-В-клітинний рецептор, мають форму бластів, утворюють клони : 6 поділів, не працюють рекомбінази. D — малі пре-В-клітини , перебудова генів легкого κ –ланцюга, який “експресується” на мембрані разом з перебудованим μ — важким ланцюгом. Непродуктивність одного гена -включається інший, якщо ні — перебудова генів в λ -локусі. E — незрілі В-клітини , мають сформовані Ig M -рецептори, але підлягають негативному відбору. На цій стадії В клітини виходять з кісткового мозку до селезінки і вступають на стадію дозрівання. F — зрілі наївні В-клітини , які рециркулюють (до зустрічі з антигеном). Таким чином, перебудова генів та синтез білкового продукту пов’язані з: а) перебудовою важкого ланцюга І g на стадії про-В-клітин; б) перебудовою легкого ланцюга І g на стадії пре-В-клітин. Стадії дозрівання В-лімфоцитів (2)
Джерела різноманіття антитіл Виділяють 6 можливих джерел різноманіття І g : 1) множинність гаметних генів (існування копій майже кожного генного сегмента); 2) соматичні рекомбінації (урізноманітнення за рахунок сайт- специфічної рекомбінації варіабельних ділянок молекули); 3) вбудовування (інсерції) та делеції додаткових нуклеотидів= junction diversity =урізноманітнення з’єднань; 4) механізм комбінаторної диверсифікації (утворення можливих варіантів імуноглобулінів); 5) соматичний мутагенез; 6) генна конверсія (активація роботи псевдо-генів)
Джерела різноманіття антитіл
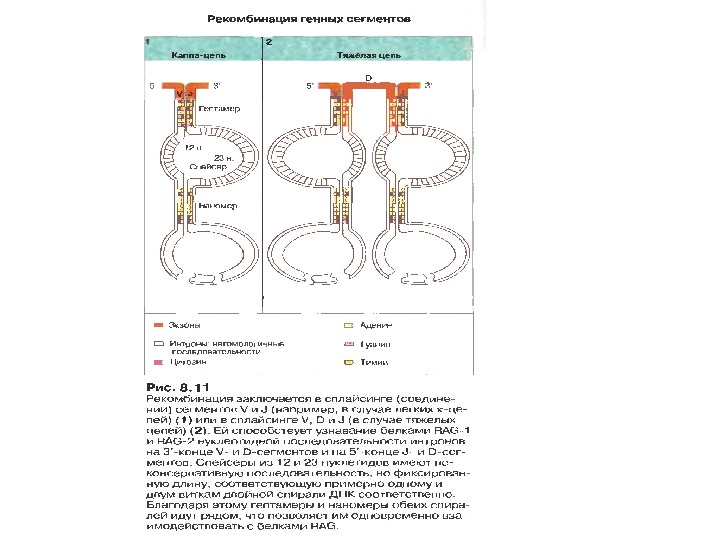
Механізми сайт- специфічної рекомбінації. Організація сигнальних послідовностей RSS s ( recombination signal sequences ) в процесі рекомбінації генів імуноглобулінів
Рекомбінація генних сегментів
Механізми сайт- специфічної рекомбінації На етапах рекомбінації залучені: • Продукти генів RAG 1 і RAG 2 — рекомбінази. Саме початок експресії цих генів є сигналом для початку перебудови генів імуноглобулінів. • У складному процесі рекомбінації на етапі з’єднання кінців ДНК беруть участь білки, які працюють в системі репарації дволанцюгових розривів ДНК [ консервативний процес NHEJ = nonhomologous DNA end — joining = негомологічне з’єднання кінців ДНК], модифікації кінців розривів і т. д. , а саме: ДНК-залежна протеїнкіназа ( DNA — PK ) , білок Ки (гетеродимер Ки 70: 80), який асоціює з DNA — PK , ендонуклеаза Artemis , процес лігування відбувається за участі ДНК-лігази IV та ДНК-репараційного білка XRCC 4 , ендо- та екзонуклеази, термінальні дезоксинуклеотидил-трансферази ( Td. T ), полімерази та інші.
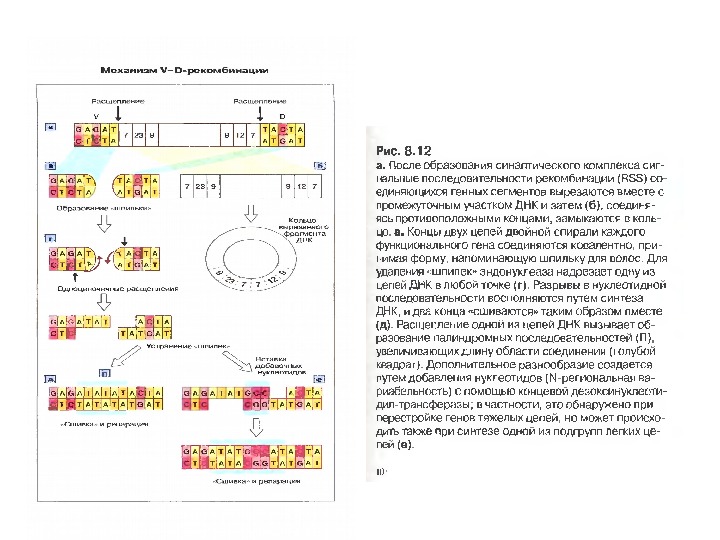
Механізм сайт- специфічної рекомбінації
Механізм сайт- специфічної рекомбінації (RAG- залежна рекомбінація ) генів V- області імуноглобулінів
Схема приєднання нуклеотидів Р до рекомбінуючих генів D і J М. Якобисяк ”Імунологія” : “Спочатку відбувається перерізання подвійної ДНК на межі сигнальних і кодуючих послідовностей з утворенням характерних «шпильок для волосся». Потім ці «шпильки» відкриваються в місцях, позначених ламаними стрілками. Виникають одноланцюгові відрізки ДНК , в яких два останні нуклеотиди комплементарні до нуклеотидів початкової послідовності (нуклеотиди Р-паліндромні). Тепер може відбутись і видалення, а також приєднання поодиноких нуклеотидів (не позначене на рисунку). Після приєднанн я комплементарних нуклеотидів відбувається відновлення кодуючої послідовності ДНК”
P — i N- нуклеотидні вставки при з ’ єднанні кодуючих сегментів генів імуноглобулінів
Будова імуноглобулінів Кодування CDR- complementarity determining regions Рекомбінація запускає транскрипцію перебудованого ланцюга, що припиняє рекомбінацію інших варіантів відповідного гена і, таким чином, визначає клональний характер лімфоцитів: кожна клітина продукує антитіла тільки однієї специфічності. Гіперваріабільні ділянки V — доменів ( CDRs ) кодуються: — на легкому ланцюгу: CDR 1, CDR 2 — на V — гені, CDR 3 — на з’єднанні V J ; — на важкому ланцюгу: CDR 1, CDR 2 — на V — гені, CDR 3 — на D -сегменті. У будові активного центру антитіл беруть участь продукти V-, D — і J — генів.
Синтез легких ланцюгів імуноглобулінів В процесі дозрівання в результаті рекомбінації ДНК відбувається зближення одного із V та J сегментів ( V + J ) – 1 рекомбінаційна подія. При цьому всі гени, що знаходяться між ними, видаляються із хромосомної ДНК (ДНК сплайсинг). Після цього з сайту ініціації транскрипції вказаного V –гену починається транскрипція з лідерної ділянки, яка закінчується на термінаторі транскрипції гена С. Утворена пре-м. РНК підлягає сплайсингу, при цьому видаляються інтрони та додаткові J -сегменти, проходить з′єднання екзонів варіабельної і константної області та лідерної послідовності, після кепування та поліаденілювання така м. РНК транслюється у легкий ланцюг каппа типу або за такими ж правилами – лямбда типу, не дивлячись на те, що в останньому J і С- сегменти є зчепленими. Для генів легких ланцюгів: V -сегмент кодує 1 -95 ам. зал. , J — сегмент -96 -108, далі — С-сегмент. (Всього-220 -230 а. з. ).
Синтез легких ланцюгів імуноглобулінів
Утворення легких κ — ланцюгів імуноглобулінів
Утворення легких κ — ланцюгів імуноглобулінів
Утворення легких λ — ланцюгів імуноглобулінів
Синтез важких ланцюгів імуноглобулінів При формуванні функційного гену важкого ланцюга відбувається ряд перебудов на рівні ДНК, а саме 2 рекомбінаційні події ( V + D J ). Спочатку відбувається об’єднання вибраних D J сегментів. При цьому всі сегменти ДНК, що є між ними, делетуються. На другому етапі — об’єднання перебудованих D J з V сегментами з наступним видаленням всіх проміжних сегментів ДНК. Таким чином шляхом рекомбінації V , D , J сегментів утворюється перебудований V ген (ДНК-сплайсинг). Нижче J генів розташовані всі гени ( відстань між V і С – 8, 5 тис. п. н. ) константних областей. Розпочинається транскрипція, яка закінчується на термінаторі транскрипції, що розташований після кластера генів Сμ і С δ. Перед кожним Сн екзоном, крім Сδ, містяться ділянки переключення — світч- сайти ( SWITCH SITE = S ), які побудовані з нуклеотидних послідовностей, що повторюються, і є в основному схожими на послідовності switch site перед екзоном Сμ. Саме switch site дозволяють певному Сн гену рекомбінувати з перебудованим геном варіабельних доменів ( VDJ ). Утворена пре-м. РНК підлягає сплайсингу, після кепування та поліаденілювання така м. РНК транслюється. Для важких ланцюгів- V –кодує 1 -99 амінок. зал, D -100 -107 амінок. зал, J — 108 -123 амінок. зал. Всього- від 440 -450 амінокислот (крім Ig. M і Ig Е).
Схема збірки активного гена важкого ланцюга імуноглобулінів
Синтез важких ланцюгів імуноглобулінів
Схема збірки активного гена важкого ланцюга імуноглобулінів
Важкі ланцюги імуноглобулінів Червоні малі квадратики — switch site ( sequence) μ δ γ 3 γ 1 γ 2 b γ 2 a ε α S — switch site ( sequence) мають колір, відповідний певним Сн — генам
Ізотипи імуноглобулінінів кодуються кластерами генів константних доменів важких ланцюгів
Механізми диверсифікації (різноманітності) = ( diversification ) імуноглобулінів Комбінаторна диверсифікація (див. табл. кластерів генів) 1) L -ланцюги : λ -ланцюг — VJ : 30 V х 4 J λ = 120; κ –ланцюг- VJ : 40 V х 5 J к =200; 120 + 200= 320 варіантів L -ланцюга 2) Н ланцюги: DJ : 27 D х 6 J = 162; V ( DJ ) : 51 х 162 = 8262; D ген має три вірогідні рамки зчитування: 8262 х 3 = 24786 варіантів Н-ланцюга. 3 ) Комбінація Н х L : 320 х 24786 = 7, 9 х 10 6 4. Мутації у сайтах рекомбінації , генна конверсія, випадковість сайтів розщеплення за участі ендонуклеази Artemis , додавання Р і N -нуклеотидів — додають різноманітності >
Соматичний гіпермутагенез ( активаційно-індукована дезаміназа (АІ D ) , урацил-ДНК-глікозилаза ( UNG))
Алельне виключення
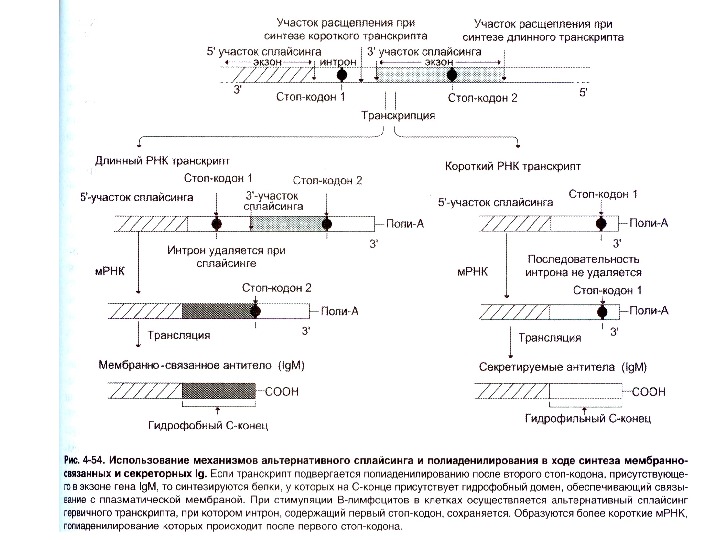
Переключення з мембранної на секретовану форму- на прикладі І g М
SC = S ecretion С oding; p. A s — polyadenylation site МС = М embrane С oding ; p. A m — polyadenylation site
Переключення з мембранної на секретовану форму- на прикладі І g М
Переключення класів імуноглобулінів S — switch site ( sequence) мають колір, відповідний певним Сн — генам
Важкі ланцюги імуноглобулінів Червоні малі квадратики — switch site ( sequence) μ δ γ 3 γ 1 γ 2 b γ 2 a ε α S — switch site ( sequence) мають колір, відповідний певним Сн — генам
Переключення класів імуноглобулінів
Переключення класів імуноглобулінів
Переключення класів імуноглобулінів за участі class — switch рекомбінації ( активаційно-індукована дезаміназа ( АІ D ) , урацил-ДНК-глікозилаза ( UNG ), апурин/апиримідин- ендонуклеаза ( АРЕ 1 )
Переключення з Ig. M на Ig. G
Переключення синтезу різних класів імуноглобулінів відбувається за участі class — switch рекомбінації ( один з типів негомологічної рекомбінації ДНК ), яка відрізняється від сайт — специфічної V ( D ) J — рекомбінації за такими ознаками: • Відбувається після антигенної стимуляції в основному в зародкових центрах та залежить від репертуару Т- хелперів. • Роль RSS тут виконують switch — site ( sequence ), що знаходяться в інтронах між Jн та С μ геном чи іншим Сн –геном, та є залученими у процес рекомбінації (об’єднання кодуючих послідовностей після делеції — рівень ДНК). • Певну роль відіграє активаційно-індукована дезаміназа (АІ D ) , а також урацил-ДНК-глікозилаза ( UNG ) та апурин/апиримідин- ендонуклеаза ( АРЕ 1 ), а не RAG білки.
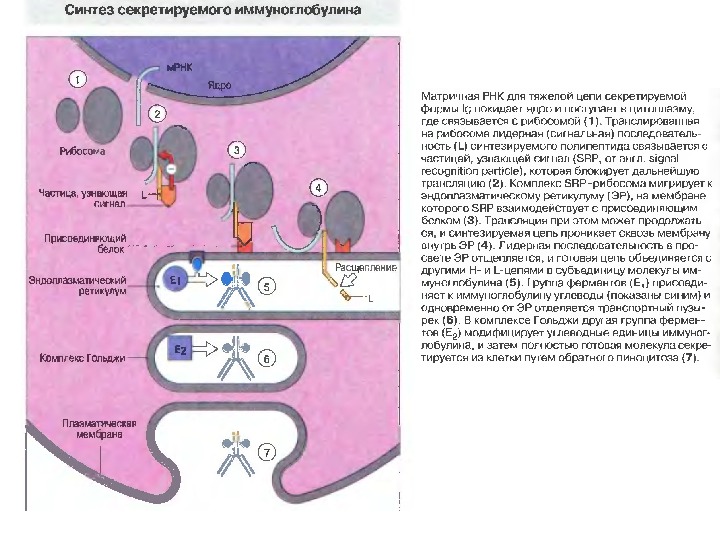
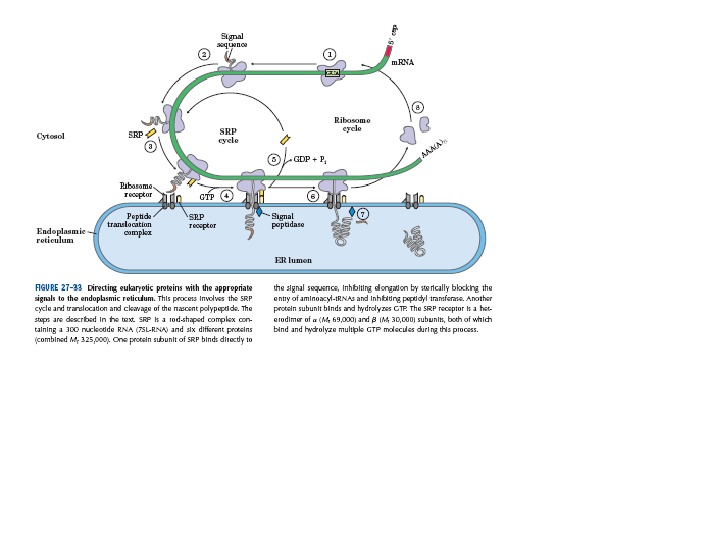
Основні етапи біосинтезу антитіл на рівні трансляції і спрямування білків ( Protein targeting )
Синтез і транспорт Ig М у мембрани й у позаклітинний простір: 1 — цис -; 2 – транс — сторони апарата Гольджі
Додатки