АНТИТЕЛА 2.pptx
- Количество слайдов: 39



АНТИТЕЛА

Антителами называются сывороточные белки, образующиеся в ответ на действие антигена • Они относятся к сывороточным глобулинам, поэтому называются иммуноглобулинами (Ig). • Через них реализуется гуморальный тип иммунного ответа. : Антитела обладают 2 свойствами специфичностьюспособностью вступать во взаимодействие с АГ, аналогичным тому, который вызвал их образование; Гетерогенностью по физико-химическому строению, специфичности, генетической детерминированности образования (по происхождению).
 это особый класс гликопротеинов, присутствующих на поверхности Вклеток в виде мембраносвязанных рецепторов")
Антитела (иммуноглобулины) это особый класс гликопротеинов, присутствующих на поверхности Вклеток в виде мембраносвязанных рецепторов и в сыворотке крови и тканевой жидкости в виде растворимых молекул. • Они являются важнейшим фактором специфического гуморального иммунитета. • Антитела используются иммунной системой для идентификации и нейтрализации чужеродных объектов Антитела выполняют функции: 1. антиген-связывающую – распознавание и/или блокирование АГ посредством активных центров (участвуют Fab фрагменты). 2. эффекторную – осуществление эффекта элиминации (реализуется Fc фрагментом). Вызывают тот или иной иммунный ответ, например, запускают классическую схему активации комплемента). Fc фрагменты Ig обеспечивают взаимодействие комплексов АГ —АТ с клетками, обеспечивающими деструкцию и удаление АГ из организма. Для этого существуют Fc рецепторы для Ig различных классов на поверхности макрофагов, лимфоцитов, нейтрофилов, эозино и базофилов, тучных, и К клеток. Взаимодействуя с Fc областью Ig, рецепторы стимулируют фагоцитоз, противоопухолевую активность, дегрануляцию тучных клеток. •

• Антитела синтезируются ПК, которыми становятся В-л в ответ на присутствие АГ • Для каждого АГ формируются соответствующие ему специализировавшиеся ПК, вырабатывающие специфичные для этого антигена антитела. • Антитела распознают АГ, связываясь с определённым эпитопом — характерным фрагментом поверхности или линейной аминокислотной цепи антигена. • Существенным и уникальным свойством АТ, отличающим их от других иммуноглобулинподобных структур, является способность взаимодействия с цельными (нативными или необработанными) АГ. Следовательно, не требуется времени на предварительную обработку и эффекторная функция осуществляется немедленно! Антитела — единственный фактор безотлагательной защиты организма, например, от сильных ЯДОВ.

Молекулы Ig одной и той же специфичности по антигену присутствуют в организме в трех формах: § растворимой (кровь и другие биологические жидкости); § на мембране В лимфоцита в составе рецептора для антигена — ВСR; § в связи с клетками, но не в трансмембранном варианте, а связанными за Fc фрагмент Fc pe цептором клетки.

Все иммуноглобулины являются иммунными, т. е. образуются в результате иммунизации, контакта с АГ Тем не менее по происхождению они делятся: • на нормальные (анамнестические) АТ, которые обнаруживаются в любом организме как результат бытовой иммунизации; • инфекционные АТ, которые накапливаются в организме в период инфекционной болезни; • постинфекционные АТ, которые обнаруживаются в организме после перенесенного инфекционного заболевания; • поствакцинальные АТ, которые возникают после искусственной иммунизации.
 – суммарная сила")
• Сила связи AT с АГ обозначают термином авидность (жадность) – суммарная сила взаимодействия цельной молекулы АТ со всеми антигенными эпитопами. Чем выше авидность, тем прочнее комплекс АТ АГ. • Аффинность — мера прочности этой связи – сила связывания одного эпитопа с одним активным центром Ig. Зависит от точности совпадения пространственной конфигурации активных центров молекул AT с АГ детерминантами. Чем выше аффинитет, тем специфичнее связь AT с АГ.

Строение иммуноглобулинов • • • АТ являются относительно крупными гликопротеинами, имеющими сложное строение. Состоят из 2 идентичных тяжелых цепей (H цепи, в свою очередь состоящие из VH, CH 1, шарнира, CH 2 и CH 3 доменов) и 2 идентичных лёгких цепей (L цепей, состоящих из VL и CL доменов). К тяжелым цепям ковалентно присоединены олигосахариды. При помощи протеазы папаина АТ можно расщепить на 2 Fab антиген связывающих фрагмента и один Fc (фрагмент, способный к кристаллизации). В зависимости от класса и исполняемых функций АТ могут существовать как в мономерной форме (Ig. G, Ig. D, Ig. E, сывороточный Ig. A) так и в олигомерной форме (димер секреторный Ig. A, пентамер — Ig. M). Всего различают пять типов тяжелых цепей (α , γ , δ , ε и μ цепи) и два типа легких цепей (κ цепь и λ цепь).

• Активный центр антител антигенсвязывающий участок Fab фрагмента Ig, образованный гипервариабельными участками Н и L цепей, связывает эпитопы АГ. • В активном центре имеются специфичные комплементарные участки к определенным антигенным эпитопам • Fc фрагмент может связывать комплемент, взаимодействует с мембранами клеток и участвует в переносе Ig. G через плаценту. • Домены антител компактные структуры, скрепленные дисульфидной связью. • Так, в Ig. G различают: V домены легких (VL) и тяжелых (VH) цепей АТ, расположенные в N концевои части Fab фрагмента; • С домены константных участков легких цепей (СL) ; • С домены константных участков тяжелых цепей (СH 1, СH 2, СH 3). Комплементсвязывающий участок находится в СH 2 домене.

• Антигенные свойства антител • Изотип антител (класс, подкласс иммуноглобулинов Ig. M, Ig. Gl, Ig. G 2, Ig. G 3, Ig. G 4, Ig. A 1, Ig. A 2, Ig. D, Ig. E) определяется С доменам тяжелых цепей; • Идиотип антител определяется антигенсвязывающими центрами Fab фрагментов антител, т. е. антигенными свойствами вариабельных участков (V областей). Идиотип состоит из набора идиотопов антигенных детерминант V области антитела.
,")
ИЗОТИП, АЛЛОТИП, ИДИОТИП Антитела — это особые растворимые белки с определенной биохимической структурой (иммуноглобулины), которые присутствуют в сыворотке крови и других биологических жидкостях и которые организм вырабатывает для связывания АГ. Международная аббревиатура иммуноглобулинов — Ig. Заглавная буква один из пяти существующих классов иммуноглобулинов — М, A, G, E, D. Последующая арабская цифра обозначает субкласс. Субклассы есть лишь у иммуноглобулинов класса G —G 1, G 2, G 3, G 4 и Ig А — Al, A 2. Изотипы В зависимости от строения константных областей тяжелых цепей иммуноглобулины разделяют на основные группы, называемые классами Классы и подклассы, вместе взятые, называют изотипами. Таким образом, классов иммуноглобулинов 5, а изотипов — 9. Они различаются не только по аминокислотным последовательностям, но и по антигенной специфичности Аллотипы Этот тип разнообразия АТ обусловлен существованием аллельных форм (кодируемых аллельными генами одного локуса); Аллотипические детерминанты служат генетическими маркерами. Подобно тому как Эр у генетически различных индивидуумов могут различаться по системам АГ групп крови А, В, О, тяжелые цепи иммуноглобулинов различаются по экспрессии аллотипических групп Gm-маркеры на Ig. G. Аллотипические различия в конкретном локусе Gm обычно затрагивают одну или две аминокислоты полипептидной цепи. Возьмем, к примеру, локус Glm(a) на Ig. Gl. У особи с этим аллотипом каждая из молекул Ig. Gl будет содержать такую последовательность: Asp. Glu. Leu. Thr. Lys. У другой особи с иным аллотипом (a negative) имеют место две аминокислотные за мены: lu. Glu. Met. Thr. Lys. На сегодняшний G день на тяжелых у цепях обнаружено 25 маркеров Gm и три маркера на константных областях к цепей (Кт маркеры).

Идиотипы Мы убедились в том, что можно получить АТ, узнающие изотипические и аллотипические варианты; кроме того, можно получить антисыворотку, специфичную к конкретному АТ и способную отличить одно Мк. АТ от другого независимо от их изотипов и аллотипов. Такая антисыворотка определяет индивидуальные особенности, характерные для каждого АТ, в совокупности названные идиотипом. Неудивительно, что идиотипические детерминанты локализованы в вариабельной области антитела и ассоциированы с гипервариабельными участками. Антиидиотипические Ig, реагирующие только с одним определенным антителом, узнают частные (private) идиотипы, что подтверждает наличие у каждого АТ уникальной структуры. Часто молекулы АТ с похожими аминокислотными последовательностями вдобавок обладают одним и тем же идиотипом , и тогда мы говорим об общих (public) или перекрестно реагирующих идиотипах. Антиидиотипическая сыворотка служит удобным реагентом для обнаружения одной и той же вариабельной области на разных тяжелых цепях и в разных клетках, для идентификации специфических иммунных комплексов в сыворотке больных.

Разнообразие иммуноглобулинов v /v c /c H L H Изотип L Изотип идиотип Аллотип Гипервариабельный (антигенсвязывающий центр) Характер изменений Распределение Все варианты обнаружены в сыворотке нормальной особи Вариант Локализация Примеры Классы Подклассы Типы CL Ig. M, Ig. E lg. A 1, lg. A 2 к, X Подгруппы CL XOz+, XOz~ Подгруппы Изотипические v. H/v. L VHIII Аллотипические Альтернативные формы: находятся под генетическим контролем, поэтому обнаруживаются не у всех особей Аллотипы В основном c. H/c. L; иногда v. H/v. L Идиотипические Индивидуальная специфичность для каждой молекулы иммуноглобулина Идиотипы Вариабельные области Группы Gm(y человека) Вероятно, примером могут служить одна или более гипервариабельных областей, формирующих антигенсвязывающий центр
 иммуноглобулинов Ig. G, Ig. M,")
Классификация по тяжелым цепям • Различают пять классов (изотипов) иммуноглобулинов Ig. G, Ig. M, Ig. A, Ig. D, Ig. E, различающихся: • величиной • зарядом • последовательностью аминокислот • содержанием углеводов • Класс Ig. G классифицируют на четыре подкласса (Ig. G 1, Ig. G 2, Ig. G 3, Ig. G 4), • класс Ig. A — на два подкласса (Ig. A 1, Ig. A 2). Все классы и подклассы составляют девять изотипов, которые присутствуют в норме у всех индивидов. Каждый изотип определяется последовательностью аминокислот константной области тяжелой цепи.

Схема строения иммуноглобулинов 5 разных классов

• • Ig. M состоит из 5 субъединиц, соединенных дисльфидными связями дополнительной J- цепью, имеет 10 АГ-детерминант. Ig. M присутствует в цитоплазме и на поверхности В-л на ранних стадиях их созревания, и это первый класс АТ, продуцируемый активированными В-л во время первичного иммунного ответа. в основном, присутствует в крови, в отличие от Ig. G, распределённого между кровью и тканями равновесно. Ig. M - действенный активатор классического пути комплемента, эффективный опсонин и агглютинин. К этому классу относятся специальные АТ : холодовые агглютинины, гетерофильные АТ и изогемагглютинины. Иммунный ответ на Т-независимые АГ , по преимуществу, проявляется в синтезе Ig. M. мигрирует при электрофорезе, в основном, в β-глобулиновой и даже, частично, a 2 глобулиновой фракциях. Иммуноглобулин М Концентрация Ig. M в крови- 0, 5 - 2, 0 г/л, период полураспада- около недели. Ig. M способны агглютинировать бактерии, нейтрализовать вирусы, активировать комплемент, активизировать фагоцитоз, связывать эндотоксины грам(-) бактерий. Ig. M обладают большей чем Ig. G авидностью (10 активных центров), аффинность (сродство к антигену) меньше, чем у Ig. G.

Ig М • Ig. M наиболее ранний класс антител, образующихся при первичном попадании антигена в организм. Наличие Ig. M антител к соответствующему возбудителю свидетельствует о свежем инфицировании (текущем инфекционном процессе). Антитела к антигенам грамотрицательных бактерий, жгутиковым антигенам преимущественно Ig. M антитела. Ig. M основной класс иммуноглобулинов, синтезируемых у новорожденных и младенцев. Ig. M у новорожденных это показатель внутриутробного заражения (краснуха, ЦМВ, токсоплазмоз и другие внутриутробные инфекции), поскольку материнские Ig. M через плаценту не проходят.

• Составляет 75% от общего количества антител • Наиболее широко распространенный мономерный класс иммуноглобулинов сыворотки, электрофоретически мигрирующий в её γ-фракции. • важный противовирусный и противобактериальный фактор • потенциальный опсонин • главный участник вторичного иммунного ответа • единственный класс антител- нейтрализатор токсинов. • Это уникальный Ig, проходящий через плаценту и способный обеспечивать пассивный иммунитет новорожденным от 3 до 6 месяцев. • активирует комплемент по классическому пути. Иммуноглобулин G (Ig. G)

Ig. G • Концентрация в крови- от 8 до 17 г/л, период полураспада- около 3 - 4 недель. Это основной класс иммуноглобулинов, защищающих организм от бактерий, токсинов и вирусов. В наибольшем количестве Ig. G- антитела вырабатываются на стадии выздоровления после инфекционного заболевания (поздние или 7 S антитела), при вторичном иммунном ответе. Ig. G 1 и Ig. G 4 специфически (через Fab- фрагменты) связывают возбудителей (опсонизация), благодаря Fc- фрагментам Ig. G взаимодействуют с Fc- рецепторам фагоцитов, способствуя фагоцитозу и лизису микроорганизмов. Ig. G способны нейтрализовать бактериальные экзотоксины, связывать комплемент. Только Ig. G способны транспортироваться через плаценту от матери к плоду (проходить через плацентарный барьер) и обеспечивать защиту материнскими антителами плода и новорожденного. В отличие от Ig. M- антител, Ig. G- антитела относятся к категории поздних- появляются позже и более длительно выявляются в крови.

Подклассы Ig. G • В зависимости от типа тяжёлой цепи, иммуноглобулины класса G подразделяются на 4 подкласса: Ig. G 1–Ig. G 4 • Ig. G 1 (70%) > Ig. G 2 (20%) > Ig. G 3 (6%) і Ig. G 4 (4%) • Структура шарнирного участка заметно отличается у представителей различных подклассов иммуноглобулина G (наиболее длинный – у Ig. G 3, самый короткий – у Ig. G 2. • В целом, гибкость и подвижность частей молекулы относительно друга у подклассов иммуноглобулина G снижается в ряду: Ig. G 3 > Ig. G 1 > Ig. G 4 > Ig. G 2

Функции подклассов Ig. G • наиболее активен подкласс Ig. G 3, шарнирная область которого обеспечивает молекуле антител максимальное число степеней свободы • По способности взаимодействовать с С 1 q на первом месте стоит подкласс Ig. G 3 • У человека все подклассы Ig. G способны активно проникать через плаценту в организм плода, создавая пассивный иммунитет • Ig. G 2 человека играет ключевую роль в инактивации инкапсулированных бактерий

Динамика возрастных изменений концентрации Ig. G и его подклассов
 • • Ig. G 1 - 4,")
Концентрация подклассов Ig. G у взрослых (г/л) • • Ig. G 1 - 4, 9– 11, 4; Ig. G 2 - 1, 5– 6, 4; Ig. G 3 - 0, 2– 1, 1; Ig. G 4 - 0, 08– 1, 4

Ig. A мономер • • • Иммуноглобулин А димер Ig. A у человека в сыворотке присутствует на 80% как мономер, в слизистых - димер. Ig. A - главный секреторный продукт системы лимфоидной ткани верхних и нижних дыхательных путей, урогенитального и пищеварительного трактов, фактор иммуной защиты кожных секретов, молозива и молока. В основном Ig. A, особенно в пищеварительном тракте, имеет специальную конфигурацию, которая препятствует протеолитическому перевариванию. Такой секреторный Ig. A содержит 2 Ig. A молекулы, соединенные в комплекс специальной полипептидной цепью, и именно этот участок и называется секреторным. Ig. A особенно эффективны в обеспечении местной антимикробной защиты в различных участках слизистых. Существует 2 подкласса Ig. A: Ig. A 1 и Ig. A 2. При электрофорезе они мигрируют не только в γ- но и в β-глобулиновой фракции.

• Ig. A не проходит через плацентарный барьер, уровень его у новорожденных около 1% от концентрации у взрослых (а к году этот показатель составляет всего 20 % от уровня взрослых). Поэтому новорожденным в первые дни жизни секреторные Ig. A поступают с молозивом матери, защищая их дыхательный пути и желудочно кишечный тракт до тех пор, пока не сформируются механизмы его синтеза в собственном организме. • Возраст 3 мес. многими авторами определяется как критический период; этот период особенно важен для диагностики врожденной или транзиторной недостаточности местного иммунитета. Уровня, характерного для взрослого человека, достигает примерно к 5 летнему возрасту
 и секреторные Ig. A (Ig.")
• • • Выделяют сывороточные Ig. A (мономер) и секреторные Ig. A (Ig. As). Сывороточные Ig. A составляют 1, 4 4, 2 г/л. Секреторные Ig. As находятся в слюне, пищеварительных соках, секрете слизистой носа, в молозиве. Они являются первой линией защиты слизистых, обеспечивая их местный иммунитет. Ig. As состоят из Ig мономера, J цепи и гликопротеина (секреторного компонента). Выделяют два изотипа Ig. A 1 преобладает в сыворотке, субкласс Ig. A 2 — в экстраваскулярных секретах. Секреторный компонент вырабатывается эпителиальными клетками слизистых оболочек и присоединяется к молекуле Ig. A в момент прохождения последней через эпителиальные клетки. Механизм секреции Ig. A на поверхности слизистой оболочки. Клетки слизистой синтезируют иммуноглобулиновый рецептор, который встраивается в базальную мембрану. Димер Ig. A связывается с этим рецептором и транспортируется к апикальной поверхности клетки с помощью эндоцитарной вакуоли. При расщеплении рецептора высвобождается Ig. A, который все еще связан с частью рецептора, получившей название «секреторный компонент» (secretory piece). Секреторный компонент повышает устойчивость молекул Ig. As к действию протеолитических ферментов. Основная роль Ig. A- обеспечение местного иммунитета слизистых. Они препятствуют прикреплению бактерий к слизистым, обеспечивают транспорт полимерных иммунных комплексов с Ig. A, нейтрализуют энтеротоксин, активируют фагоцитоз и систему комплемента.

Ig. E • Представляет мономер, в сыворотке крови находится в низких концентрациях. Прежде всего, Ig E — это поверхностный клеточный рецептор. В сыворотке он пребывает транзиторно при передвижении от плазматических к тучным клеткам и базофилам. Основная роль - своими Fc- фрагментами прикрепляется к тучным клеткам (мастоцитам) и базофилам и опосредует реакции гиперчувствительности немедленного типа. К Ig. E относятся “антитела аллергии”- реагины. Уровень Ig. E повышается при аллергических состояниях, гельминтозах. Антигенсвязывающие Fab- фрагменты молекулы Ig. E специфически взаимодействует с антигеном (аллергеном), сформировавшийся иммунный комплекс взаимодействует с рецепторами Fc- фрагментов Ig. E, встроенных в клеточную мембрану базофила или тучной клетки. Это является сигналом для выделения гистамина, других биологически активных веществ и развертывания острой аллергической реакции.

Ig. D • Мономеры Ig. D обнаруживают на поверхности развивающихся В- лимфоцитов, в сыворотке находятся в крайне низких концентрациях (составляет менее 1% всех Ig сыворотки). Их биологическая роль точно не установлена. Полагают, что Ig. D участвуют в дифференциацировке В-клеток (мембранный Ig D вместе с Ig M являются критерием зрелости В лимфоцитов), способствуют развитию антиидиотипического ответа (участвуют в аутоиммунных процессах), развитии лимфопролиферативной патологии.

Роль антител в формировании иммунитета. Антитела имеют важное значение в формировании приобретенного постинфекционного и поствакцинального иммунитета. 1. Связываясь с токсинами, антитела нейтрализуют их, обеспечивая антитоксический иммунитет. 2. Блокируя рецепторы вирусов, антитела препятствуют адсорбции вирусов на клетках, участвуют в противовирусном иммунитете. 3. Комплекс антиген- антитело запускает классический путь активации комплемента с его эффекторными функциями (лизис бактерий, опсонизация, воспаление, стимуляция макрофагов). 4. Антитела принимают участие в опсонизации бактерий, способствуя более эффективному фагоцитозу. 5. Антитела способствуют выведению из организма (с мочой, желчью) растворимых антигенов в виде циркулирующих иммунных комплексов. Ig. G принадлежит наибольшая роль в антитоксическом иммунитете, Ig. M- в антимикробном иммунитете (фагоцитоз корпускулярных антигенов), особенно в отношении грамотрицательных бактерий, Ig. A- в противовирусном иммунитете (нейтрализация вирусов), Ig. As- в местном иммунитете слизистых оболочек, Ig. E- в реакциях гиперчувствительности немедленного типа.

Циркулирующие иммуноглобулины могут обеспечивать следующие формы антибактериальной защиты: • нейтрализацию бактериальных токсинов, например, столбнячного. • инактивацию факторов вирулентности и распространения бактерий, например, гиалуронидазы • блокаду бактериальных рецепторов ростовых факторов, например железосвязывающих белков. • опсонизирующий эффект для фагоцитоза бактерий • литический эффект через комплемент • блокаду подвижности бактериальных органоидов движения • предотвращение прикрепления бактерий к клеткам организма путём занятия соответствующих адгезивных молекул

МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА • настоящее время методами биотехнологии и/или генной инженерии получают иммуноглобулины, продуцируемые одним клоном клеток. • Они называются моноклональными антителами. Их продуценты — клетки-гибридомы, являющиеся потомками, полученными при скрещивании Влимфоцита (ПК) с опухолевой клеткой. • От плазматической клетки-гибридома наследуется способность к синтезу АТ, а от опухолевой клетки — способность длительно культивироваться вне организма.

Схема получения моноклональных антител

Смесь клеток помещают в селективную среду, которая позволяет расти только гибридным клеткам. Погибают не слившиеся миеломные клетки и Влимфоциты. Гибридные клетки пролиферируют, образуя клон гибридом. Гибридомы, проверяются на продукцию нужных антител.

«Мышиные фабрики» по производству моноклональных антител • Для получения больших объемов жидкости, содержащей моноклональные антитела, гибридомы могут быть введены внутрибрюшинно мышам реципиентам. • В результате в брюшной полости вырастает солидная опухоль и накапливается асцитическая жидкость, являющаяся источником моноклональных антител.

Применение МКАТ • • • Они являются наиболее четким инструментом поиска самых различных молекулярных структур клеток и тканей. С их помощью выявлена и продолжает изучаться молекулярная характеристика по CD антигенам клеток иммунной системы. Уточняются процессы дифференцировки иммунологически значимых клеток, их относительного участия в различных функциональных проявлениях, выясняется связь клеток иммунитета с клетками иных систем организма. В медицинской практике наибольшее применение нашли в онкологии. С помощью меченых АТ определяются метастазы различных форм опухолевого поражения для выработки наиболее адекватной терапии. Во всем мире ведутся обширные исследования в области использования МКАТ в качестве вектора, доставляющие фармакологические, токсические препараты в пораженные раковые и другие патологически измененные клетки. У нас в стране наиболее активно в этом направлении работает С. Г. Егорова (МГУ). Вклад Г. Келлера и Ц. Милыптейна в разработку технологии получения МАТ оценен присуждением им в 1984 г. Нобелевской премии по медицине.

Области применения моноклональных антител • идентификация субпопуляций лимфоцитов человека • выделение клеток • установление функций молекул клеточной поверхности • определение группы крови • диагностика опухолей и локализация опухолей • иммунорадиометрический анализ • анализ сложных смесей антигенов • анализ эмбрионального развития • анализ иммунного ответа • искусственные ферменты

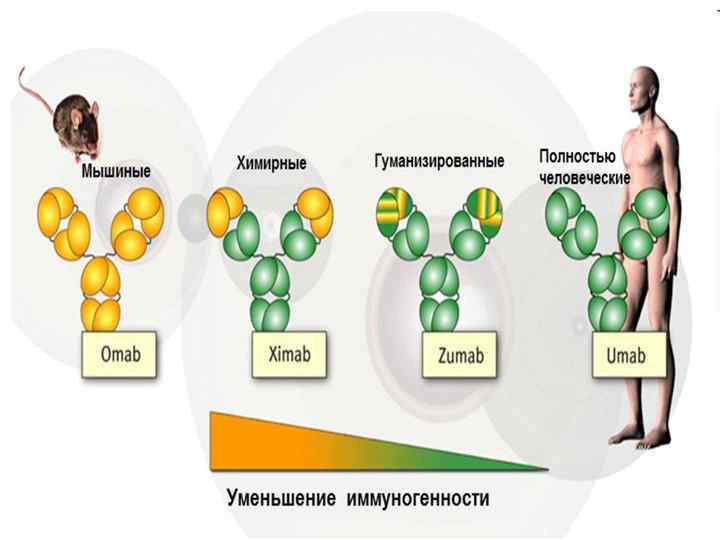
Терапевтические моноклональные антитела • • Ученые сумели принудить клетки грызунов к выработке антител с человеческими Fc-фрагментами (химерные АТ) Константная часть мышиных антител замещена соответствующей константной областью иммуноглобулина человека и в своей структуре имеют более 65% человеческого иммуноглобулина. Гуманизированные моноклональные антитела – до 95% состоят из человеческого иммуноглобулина. Трансгенные технологии (фаговый дисплей) были использованы для создания полностью человеческих моноклональных антител Моноклональные антитела, в отличие от традиционных препаратов, высокоспецифичны к определенным мишеням - антигенам. Поэтому, использование моноклональных антител в качестве терапевтических агентов стало для медицины стратегическим этапом в смене концепции лечения – от неспецифической к специфической (таргетной) терапии.

Химерные антитела получили окончание «-ximab» Гуманизированные антитела имеют окончание «-zumab» Препараты на основе мон-АТ - оканчиваются на «–mab» (от monoclonal antibody). Полностью человеческие антитела имеют окончание – «-umab» .
АНТИТЕЛА 2.pptx