Антигены и антитела.ppt
- Количество слайдов: 70



АНТИГЕНЫ

Понятие об антигенах Антиген – это полимер органической природы, генетически чужеродный для макроорганизма, при попадании в последний вызывающий иммунные реакции, направленные на его устранение. n Происхождение: из любого чужого организма или клетки; из собственного организма (эпигенетическая или генетическая мутация) клеток; получены искусственно. В любом случае молекулу АГ будет отличать генетическая чужеродность по отношению к макроорганизму, в который она попала.

Строение антигенов Функциональная часть – «антигенная детерминанта» , или «эпитоп» , 5 -6 аминокислотных или ЛПС остатков. Различают линейные, или секвенциальные антигенные детерминанты (например, первичная аминокислотная последовательность пептидной цепи) и поверхностные, или конфармационные (возникшие в результате вторичной, третичной или четвертичной конформации). Кроме того, существуют концевые (расположенные на концевых участках молекулы АГ) и центральные эпитопы. Определяют также «глубинные» , или «скрытые» , антигенные детерминанты, которые проявляются при разрушении биополимера. n

Строение антигенов n n n Денатурация приводит к полной или частичной потере антигенных детерминант или появлению новых, при этом теряется специфичность антигена. Поскольку молекулы большинства АГ имеют довольно большие размеры, в их структуре определяется множество антигенных детерминант, которые распознаются разными по специфичности АТ и клонами лимфоцитов. Валентность молекулы антигена: 1 эпитоп на 5000 дальтон относительной молекулярной массы молекулы антигена.
.")
Свойства антигенов: чужеродность; n антигенность; n специфичность; n иммуногенность; n макромолекулярность; n коллоидность (растворимость). n

Чужеродность n n n Чем дальше в филогенетическом развитии организмы отстоят друг от друга, тем большей иммуногенностью обладают их АГ по отношению друг к другу. Используют биологи и палеонтологи (при изучении филогенеза, уточнении классификации и т. д. ), судебно-медицинские эксперты и криминалисты (установление кровного родства, принадлежности улик, фальсификации пищевых продуктов и т. д. ). Перекрестно реагирующие антигены. Сходство детерминант стрептококка, сарколеммы миокарда и базальной мембраны почек, Treponema pallidum и липидной вытяжки из миокарда крупного рогатого скота, возбудителя чумы и эритроцитов человека 0 (I) группы крови. Явление, когда один микроб маскируется антигенами другого микроба или макроорганизма для защиты от факторов иммунитета, называется антигенной мимикрией.

Свойства антигенов n n Антигенность – потенциальная способность молекулы АГ к специфическому взаимодействию с факторами иммунной системы (антитела, клон лимфоцитов). Взаимодействие происходит не со всей молекулой одновременно, а только с антигенной детерминантой (эпитопом). Поэтому антигенность вещества зависит от наличия и числа антигенных детерминант в структуре его молекулы. Специфичность – способность антигена избирательно реагировать со строго определенными АТ или клонами лимфоцитов. Это свойство также зависит от наличия в структуре молекулы вещества тех или иных антигенных детерминант.

Иммуногенность – свойство АГ вызывать в макроорганизме иммунный ответ. Степень иммуногенности зависит от 3 групп факторов: свойства самого АГ; динамику поступления АГ в организм и его выведения; состояния макроорганизма. n

Свойства самого АГ n n n Чужеродность. Природа: белки → ЛПС, гликопротеины, липопротеины → чистые полисахариды, нуклеиновые кислоты и липиды. Химический состав: остатки ароматических аминокислот (тирозина, триптофана) и левовращающих изомеров высокоиммуногенны; полипептиды, построенные из Dаминокислот, практически лишены иммуногенности; сополимеры нескольких аминокислот более иммуногенны, чем одной. Молекулярная масса: минимальная масса 10 000 дальтон. На нуклеиновые кислоты описанные закономерности практически не распространяются. Структура: большей иммуногенностью обладают агрегаты молекул и карпускулярные АГ – целые клетки (эритроциты, бактерии и другие); глобулярные белки более иммуногенны, чем фибриллярные. Растворимость АГ: высокомолекулярные белки кератин и меланин не могут быть получены в виде коллоидного раствора и не являются АГ.

Динамика поступления и выведения АГ n n n Способ введения АГ: вакцину против полиомиелита вводят перорально, против сибирской язвы – накожно, БЦЖ – внутрикожно, АКДС – подкожно, против столбняка – внутримышечно. Количество поступающего АГ: чем его больше, тем более выражен иммунный ответ. Передозировка АГ вызывает обратную реакцию – иммунологическую толерантность. Чувствительность к катаболическому разрушению ферментами макроорганизма: белки, построенные из Lаминокислот, легко поддаются ферментативной деградации и высокоиммуногенны, полипептиды из правовращающих изомеров, напротив, медленно расщепляется ферментами макроорганизма и низкоиммуногенны, а при введении высоких доз приводят к развитию иммунологической толерантности.

Состояние макроорганизма n n Результат иммунизации связан с генотипом особи. Существуют чувствительные и нечувствительные роды и виды животных, которые используют в лабораторной работе. Даже внутри вида можно выделить группы близкородственных особей (инбредные линии животных), которые по-разному будут отвечать на вводимый АГ. Известны значительные индивидуальные различия между людьми в чувствительности к вакцинам: выделяют иммунологически реактивных и иммунологически инертных индивидуумов. Функциональное состояние макроорганизма – психоэмоциональный и гормональный фон, интенсивность обменных процессов и др.
 – способны неспецифически усиливать иммуногенность")
Управление иммуногенностью АГ Адьюванты (от лат. adjuvare – помогать) – способны неспецифически усиливать иммуногенность АГ. n Функции адьювантов: депо АГ; стимуляция фагоцитоза; митогенное действие на лимфоциты; продукция цитокинов. Адьюванты используют при вакцинации и в научноисследовательской работе. В качестве адьювантов используют гидроксид или фосфат алюминия, масляные эмульсии. Роль адьюванта может выполнить ЛПС грамотрицательных бактерий, служащий неспецифическим стимулятором В-лимфоцитов.

Иммуногенность и толерогенность АГ Иммунный ответ Пролиферация АРК Толерантность Гибель
 и эндогенные (возникшие внутри")
Классификация антигенов n n По происхождению: экзогенные (возникшие вне организма) и эндогенные (возникшие внутри организма). По природе: биополимеры белковой (протеины) и небелковой природы (полисахариды, липиды, ЛПС, нуклеиновые кислоты и др. ). По структуре: глобулярные (молекула имеет шаровидную форму) и фибриллярные (молекула имеет форму нити). По необходимости участия Т-лимфоцитов в индукции иммунного ответа: Т-зависимые и Т-независимые. Тнезависимые АГ имеют относительно простое строение – монотонно повторяющиеся последовательности с многочисленными однотипными эпитопами

Классификация антигенов По иммуногенности: полноценные и неполноценные. n Полноценные АГ обладают выраженной иммуногенностью и антигенностью, они как правило, имеют молекулярную массу более 10000, большой размер молекулы (частицы) в виде глобулы и хорошо взаимодействуют с факторами иммунитета. n Неполноценные АГ (гаптены) не облодают иммуногенностью, но обладают антигенностью. Чаще всего гаптенами являются низкомолекулярные соединения (молекулярная масса меньше 10000). Для придания гаптену свойств полноценного АГ необходимо соединить его с достаточно большой белковой молекулой ( «шлеппер» – тягач).

Классификация антигенов n n n По степени чужеродности: ксено-, алло и изоантигены. Ксеногенные АГ (или гетерологичные) – общие для организмов, относящихся к разным родам и видам. Эти АГ получили названия «антигены Форсмана» . Аллогенные АГ (или групповые) – антигены, общие для генетически не родственных организмов, но относящихся к одному виду: АГ группы крови (система АВ 0 и др. ), раковоэмбриональные АГ (α-фетопротеин, трансферрин) и др. Аллогенные ткани при трансплантации иммунологически несовместимы – они отторгаются макроорганизмом. Микробы на основании групповых АГ могут быть подразделены на серогруппы. Изогенные АГ (или индивидуальные) – антигены, общие только для генетически идентичных микроорганизмов, например для однояйцевых близнецов. Изотрансплантанты не отторгаются при пересадке. Примеры: АГ гистосовместимости людей, типовые АГ бактерий.

Классификация антигенов n Аутоантигены – АГ собственного организма. В нормальных условиях они не вызывают реакций иммунной системы в следствии либо иммунологической толерантности (невосприимчивости), либо недоступности для контакта с факторами иммунитета – это так называемые забарьерные антигены. При срыве толерантности или нарушении целостности биологического барьера (травма) компоненты иммунной системы начинают специфически реагировать на аутоантигены выработкой специфических факторов иммунитета (аутоантител, клон аутореактивных лимфоцитов).

Антигены организма человека n n Аллоантигенные АГ: Антигены системы АВ 0 синтезируются предшественниками эритроцитов и многими другими клетками организма. Это высокогликозилированные пептиды: 85% приходится на углеводную часть и 15% – на полипептидную. Иммуногенность определяется углеводной частью. АГ системы АВ 0 наследуются независимо аллельно, что определяет наличие в популяции 4 групп крови: 0 (I), A (II), В (III) и AB (IV). Переливание пациенту несовместимой по группе крови, как правило, приводит к развитию острого состояния – гемолитического шока. Антигены система резус (Rh) синтезируются предшественниками эритроцитов и обнаруживаются главным образом на эритроцитах, т. к. они не растворимы в биологических жидкостях. По химической структуре – термолабильный липопротеин. В зависимости от наличия или отсутствия резус-АГ в популяции людей различают две группы – резус-положительных и резус-отрицательных индивидуумов. При беременности резус-отрицательной матери резус-положительным плодом может развиться резус-конфликт, который проявляется выработкой антирезусных антител и невынашиванием беременности или желтухой новорожденного (внутрисосудистый иммунный лизис).

Антигены организма человека n Генетические локусы, ответственные за отторжение чужеродных тканей, образуют в хромосоме область, названную главным комплексом гистосовместимости (МНС, от англ. major histocompatibility complex). Продукты генов МНС у человека относятся к системе лейкоцитарных антигенов HLA (от англ. Human Leukocyte Antigen – АГ лейкоцитов человека). HLA – гликопротеины, которые прочно связаны с клеточной мембраной.

HLA I класса n HLA I класса находятся на мембранах всех клеток организма за исключением эритроцитов. HLA I класса обусловливают биологическую индивидуальность ( «биологический паспорт» ) и являются маркерами «своего» для иммунокомпетентных клеток (Т-хелперы, Ткиллеры). Клетки, отличающиеся по HLA I класса, уничтожаются как чужеродные. Распознавание антигена Т-лимфоцитом

HLA II класса n n HLA II класса обнаруживаются на клеточной мембране антигенпредставляющих клеток. HLA II класса участвуют в представлении чужеродного АГ иммунокомпетентным клеткам для их специфического распознавания.

Опухолевые антигены n n Злокачественное перерождение нормальной клетки сформированного макроорганизма сопровождается началом биосинтеза особых белков, которые встречаются лишь в эмбриональном периоде развития. Такие белки получили название опухолевых, или раково-эмбриональных АГ. К опухолевым АГ относятся α-фетопротеин, трансферрин и др. Лабораторное определение этих АГ имеет большое клиническое значение для ранней диагностики некоторых новообразований, в частности первичного рака печени.
. Около")
CD-антигены n n n Антигены кластерной дифференцировки (Cell Differentiation Antigens или Claster Definition). Около 200 вариантов. Гликопротеины, относятся к суперсемейству иммуноглобулинов. Маркеры иммунокомпетентных клеток: CD 3 – Т-лимфоциты; CD 4 – Т-хелперы; CD 8 – Тцитотоксические лимфоциты; CD 11 a – монои гранулоциты; CD 11 b – естественные киллеры; CD 19 -22 – В-лимфоциты и др.

Антигены бактерий n n У различных микроорганизмов различают группоспецифические АГ – встречаются у разных видов одного и того же рода или семейства, видоспецифические – у различных представителей одного вида и типоспецифические АГ – у разных вариантов в пределах одного и того же вида. По наличию типовых АГ виды микроорганизмов подразделяются на серологические варианты, или серовары. В структуре бактериальной клетки различают жгутиковые, соматические, капсульные и некоторые другие АГ.

Антигены бактерий n n Жгутиковые, или H-АГ представляют собой сократительный белок флагеллин. При нагревании H-АГ денатурируют и теряют свою специфичность. Фенол не действует на эти АГ. Соматический, или О-АГ связан с клеточной стенкой бактерий (ЛПС), термостабилен, но подвержен действию альдегидов (например, формалина) и спиртов.
, располагаются на поверхности клеточной стенки, встречаются у бактерий, образующих")
n n n Капсульные (К-АГ), располагаются на поверхности клеточной стенки, встречаются у бактерий, образующих капсулу. Чаще состоят из кислых полисахаридов (уроновые кислоты), но у бациллы сибирской язвы – из полипептидных цепей. По чувствительности к нагреванию различают три типа К-АГ: A, B и L. Вариантом К-АГ является Vi-АГ. Антигенными свойствами обладают бактериальные белковые токсины, ферменты и некоторые белки, которые секретируются бактериями (туберкулин). АГ с сильно выраженной иммуногенностью, которые могут полностью обеспечить иммунитет макроорганизма ко всему инфекционному агенту называются протективными. Антигены бактерий
, капсидные (или оболочечные) и суперкапсидные")
Антигены вирусов n n Вирусные АГ: ядерные (или коровые), капсидные (или оболочечные) и суперкапсидные (гемагглютинин, нейраминидаза и др. ). Вирусспецифические АГ; АГ – компоненты клетки хозяина (углеводы, липиды). Антигенная специфичность простых вирусов связана с рибо- и дезоксирибонуклеопротеидами; сложных с нуклеокапсидом и суперкапсидом. АГ многих вирусов отличаются высокой степенью изменчивости.

Процессы, происходящие с АГ в макроорганизме n n n Происхождение АГ: эндогенное, экзогенное. Пути проникновения АГ в макроорганизм: через дефекты кожных покровов и слизистых оболочек (как результат ранений, микротравм, укусов насекомых, расчесов и др. ); путем всасывания в ЖКТ (эндоцитоз эпителиальными клетками); межклеточно (при незавершенном фагоцитозе, облигатном или факультативном внутриклеточном паразитировании микроб может разноситься по всему организму); чресклеточно (так распространяются облигатные внутриклеточные паразиты, например вирусы). Проникнув в организм, АГ разносится лимфой по разным органам и тканям. Чаще всего он накапливается в лимфоидной ткани печени, селезенки, легких и других органов, где и вызывает иммунологические реакции.
 антигена")
Ответная реакция на АГ n n n n Пути инактивации и удаления (элиминации) антигена из макроорганизма: фильтрация и концентрирование АГ в лимфоидных образованиях; специфическое распознавания «свой-чужой» ; осуществление иммунного реагирования; выработка факторов регуляции и иммунитета (АТ, клоны лимфоцитов); связывание и блокирование биологически активных участков молекулы АГ; разрушение и отторжение АГ; полная утилизация, изоляция (инкапсуляция) или выведение из макроорганизма остатков АГ. В итоге достигается полное или частичное восстановление гомеостаза, формирование иммунологической памяти, ареактивности или гиперергии.

ИММУНОГЛОБУЛИНЫ: КЛАССИФИКАЦИЯ И ФУНКЦИИ. ПОЛНЫЕ И НЕПОЛНЫЕ АНТИТЕЛА.
 – это иммуноглобулины, вырабатываемые в ответ на введение АГ и способные специфически")
Антитела (АТ) – это иммуноглобулины, вырабатываемые в ответ на введение АГ и способные специфически связываться с АГ и участвовать во многих иммунологических реакциях. Поэтому они получили название иммуноглобулинов, и их обозначают символом Ig.

Антитела синтезируются В-лимфоцитами. Для этого необходим контакт с АГ и вызванное им созревание В-клеток в антителобразующие клетки (АОК). К АОК относятся плазматические клетки, секретирующие значительное количество АТ.
, рецепторных")
Иммуноглобулины существуют в форме: n n n циркулирующих АТ (сывороточные и секреторные ), рецепторных молекул на иммунокомпетентных клетках миеломных белков (белки Бенс-Джонса)

Каждый иммуноглобулин выполняет функции: 1. 2. Одна область его молекулы предназначена для связывания с АГ; Другая область осуществляет так называемые эффекторные функции: связывание иммуноглобулина с тканями организма, различными клетками иммунной системы, определенными фагоцитарными клетками и первым компонентом комплемента (С 1 q) при активации этой системы по классическому пути.

Антитела используют: n для диагностики, n профилактики и лечения соматических и инфекционных болезней. n На основе специфических иммуноглобулинов созданы соответствующие иммунобиологические препараты (лечебные и диагностические сыворотки и др. ).

Молекулярное строение АТ. Иммуноглобулины являются гликопротеинами. Их молекула состоит из полипептидной цепи, стабилизированной сахаридными остатками. Иммуноглобулины различаются по структуре, антигенному составу, а также по выполняемым ими функциям. По этим свойствам они подразделяются на 5 классов: Ig. G, Ig. M, Ig. А, Ig. E, Ig. D.

Антитела выполняют функции: n n n n Специфически связываясь с антигенными детерминантами, осуществляют маркирование АГ, инактивируют биологически активные молекулы (например, токсинов), участвуют в опсонизации АГ, антителоопосредованном лизисе клеток, иммунном фагоцитозе, в развитии реакции гиперчувствительности немедленного типа. антигенспецифического рецептора на поверхности Влимфоцитов.

Молекула иммуноглобулина состоит:

n Легкие и тяжелые полипептидные цепи молекулы иммуноглобулинов имеют определенные варианты структуры, или типы. Они определяются первичной аминокислотной последовательностью цепей и степенью их гликозилирования. Легкие цепи бывают двух типов – κ и λ. Известно 5 типов тяжелых цепей: α, γ, μ, ε и δ, – которые имеют также и внутреннее подразделение.
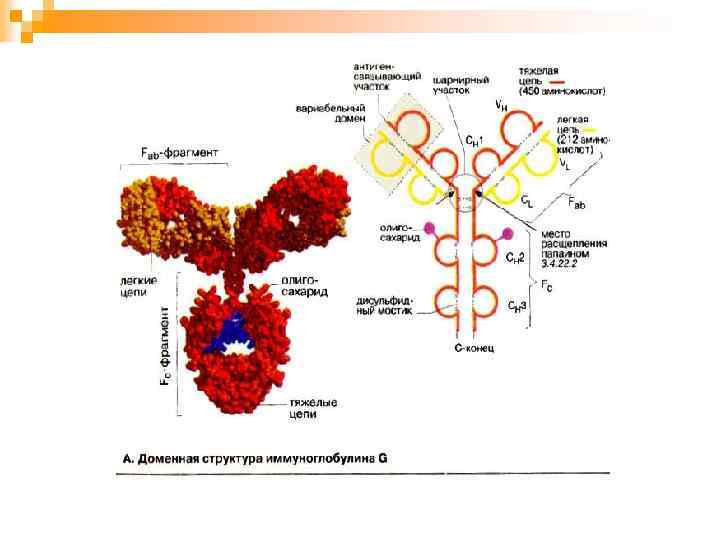

В структуре молекул иммуноглобулинов есть дополнительные полипептидные цепи: n n n J-пептид (от англ. join – соединяю). Он объединяет отдельные мономеры в единое макромолекулярное образование (полимерные молекулы Ig. M, Ig. A ); S-пептидом (от англ. secret – секрет) - секреторный компонент. Он предохраняет АТ в секрете слизистых оболочек от ферментативного расщепления; М-пептид (от англ. membrane – мембрана). Благодаря своим гидрофобным свойствам он прочно удерживается в липидном бислое цитоплазматической мембраны и, как якорь, фиксирует рецепторный иммуноглобулин на мембране иммунокомпетентной клетки (В-лимфоцитов и плазматических клеток).

n J- и M-пептиды присоединяются к молекуле иммуноглобулина в процессе биосинтеза этого белка. S-пептид синтезируется эпителиальными клетками и является их рецептором для Ig. A. Присоединение S-пептида к молекуле Ig. A происходит при его прохождении через эпителиальную клетку.

Антигенность АТ. n В молекуле иммуноглобулина различают 4 типа антигенных детерминант: n видовые, n изотипические, n Аллотипические n идиотипические

n n Видовые детерминанты характерны для иммуноглобулинов всех особей данного вида (например, кролика, собаки, человека). Они определяются строением легкой и тяжелой цепей. По этим детерминантам можно идентифицировать видовую принадлежность АТ. Изотипические детерминанты являются групповыми. Они локализуются в тяжелой цепи и служат для дифференцировки семейства иммуноглобулинов на 5 изотипов (классов) и множество подклассов.

n n Аллотипические детерминанты являются индивидуальными, т. е. присущими конкретному организму, они располагаются в легкой и тяжелой полипептидных цепях. На основании строения аллотипических детерминант можно различать особи внутри одного вида. Идиотипические детерминанты отражают особенности строения антигенсвязывающего центра молекулы иммуноглобулина. Они образованы V-доменами легкой и тяжелой цепи молекулы иммуноглобулина.

Механизм взаимодействия АТ с АГ. n В процессе взаимодействия с АГ участвует не вся молекула иммуноглобулина, а лишь ее ограниченный участок – антигенсвязывающий центр, или паратоп, который локализован в Fabфрагменте. АТ взаимодействует не со всей молекулой АГ сразу, а лишь с ее антигенной детерминантой.

n Связь АГ с АТ осуществляется за счет слабых нековалентных взаимодействий (ван-дер-ваальсовы силы, водородные связи, электростатические взаимодействия и гидрофобные взаимодействия) в пределах антигенсвязывающего центра.

Особенности АТ: n n Аффинность – сила специфического взаимодействия АТ с АГ (или энергия их связи), это прочность связи одного антигенсвязывающего центра с индивидуальным эпитопом АГ. Эта характеристика зависит от степени стерического (или пространственного) соответствия (комплементарности) структуры антигенсвязывающего центра и антигенной детерминанты. Чем выше их комплементарность, т. е. чем больше они подходят другу, тем больше образуется межмолекулярных связей и тем выше будет устойчивость образовавшегося ИК. Авидность - прочность связывания антитела и антигена (суммарная сила). Эта характеристика определяется аффинностью и числом антигенсвязывающих центров. При равной аффинности наибольшей авидностью обладают антитела класса М, так как они имеют 10 антигенсвязывающих центров.

Особенности антигена также влияют на эффективность его взаимодействия с антителом. Большое значение имеют стерическая (пространственная) доступность антигенной детерминанты для антигенсвязывающего центра молекулы иммуноглобулина и число эпитопов в составе молекулы антигена. Эффективность взаимодействия антитела с антигеном зависит от условий, в которых происходит реакция, прежде всего от р. Н среды, осмотической плотности, солевого состава и температуры среды.

Реакциям АГ-АТ свойственна высокая специфичность. Например, противокоревые АТ связываются с вирусами кори и создают иммунитет к этому заболеванию, но не способны связываться с вирусами других видов. n

Защитные функции антител АТ помогают предотвратить инфицирование и/или элиминировать возбудитель, активируя при этом несколько механизмов: n опсонизацию (иммунный фагоцитоз), n антитоксический эффект, n активацию комплемента n нейтрализацию.

Эфекторные функции антител Связывание с АГ; n активации системы комплемента; n избирательное взаимодействие с различными типами клеток при участии специальных рецепторов клеточной поверхности (фагоцитоз, зависимая от АТ n клеточная цитотоксичность, высвобождение медиаторов и презентация АГ ).

Структурно-функциональные особенности иммуноглобулинов различных классов. В зависимости от строения тяжелой цепи (т. е. наличия изотипических, или групповых, антигенных детерминант) различают классы, или изотипы иммуноглобулинов. Молекулы, содержащие тяжелую цепь α-типа, относят к изотипу А (сокращенно Ig. А), Ig. D обладает δ-цепью, Ig. Е — ε-цепью, Ig. G— γ-цепью и Ig. М — μ-цепыо. Соответственно особенностям строения подтипов тяжелых цепей различают и подклассы иммуноглобулинов. В структуре молекул иммуноглобулинов разных классов прослеживается общая закономерность: все они построены из одних и тех же элементов.

Иммуноглобулин класса G. Класс G составляет основную массу иммуноглобулинов сыворотки крови. На его долю приходится 70 – 80 % всех сывороточных иммуноглобулинов, при этом 50 % содержится в тканевой жидкости. Среднее содержание Ig. G в сыворотке крови здорового взрослого человека составляет 12 г/л. Период полураспада Ig. G равен 21 дню.

Ig. G – мономер, имеет два антигенсвязывающих центра (он может связать две молекулы антигена подряд, следовательно, его валентность равна двум), молекулярную массу – около 150 к. Да. Различают подтипы G 1, G 2, G 3 и G 4. Ig. G синтезируется зрелыми B-лимфоцитами (Вγ) и плазматическими клетками. Он хорошо определяется в сыворотке крови на пике первичного и при вторичном иммунном ответе. Ig. G составляют большинство АТ вторичного иммунного ответа, а также основную часть антитоксинов. Обладает высокой аффинностью, связывает комплемент. Ig. G 4, подобно Ig. Е, обладает сродством, к тучным клеткам и базофилам и участвует в развитии аллергической реакции I типа. Ig. G легко проходит через плацентарный барьер и обеспечивает гуморальный иммунитет новорожденного в первые 3 – 4 мес после рождения.

Обнаружение высоких титров Ig. G к АГ конкретного возбудителя указывает на то, что организм находится на стадии реконвалесценции или инфекционное заболевание перенесено недавно.

Иммуноглобулин класса М. Ig. М – пентамер, который имеет 10 антигенсвязывающих центров, т. е. его валентность равна 10. Его молекулярная масса около 950 к. Да. Различают подтипы М 1 и М 2. Период полураспада Ig. М составляет 5 дней. На долю Ig. М приходится около 5 – 10 % всех сывороточных иммуноглобулинов. Среднее содержание Ig. М в сыворотке крови здорового взрослого человека около 1 г/л. Иммуноглобулин, синтезируется предшественниками и зрелыми Bлимфоцитами (Вμ). Он образуется в начале первичного иммунного ответа, также первым начинает синтезироваться в организме новорожденного. АТ этого класса доминируют в качестве «ранних» АТ, чаще всего при иммунном ответе на сложные по антигенному составу патогенные микроорганизмы.

n n n Ig. М обладает высокой авидностью, связывает комплемент, участвует в формировании сывороточного и секреторного гуморального иммунитета. Он может выделиться в секрет слизистых оболочек путем диффузии. Ig. М не проходит через плаценту. Обнаружение специфических антител изотипа М в сыворотке крови новорожденного указывает на бывшую внутриутробную инфекцию или дефект плаценты. Наличие Ig. M к АГ конкретного возбудителя указывает на наличие острого инфекционного процесса.

Иммуноглобулин класса А Ig. А существует в сывороточной и секреторной формах. Около 60 % всех Ig. А содержится в секретах слизистых оболочек.

Сывороточный Ig. А: n n на долю Ig. А приходится около 10 – 15 % всех сывороточных иммуноглобулинов. В сыворотке крови здорового взрослого человека содержится около 2, 5 г/л Ig. А. Период полураспада Ig. А составляет 6 дней. Ig. А – мономер, имеет два антигенсвязывающих центра (т. е. 2 -валентный), его молекулярная масса около 360 к. Да. Различают подтипы А 1 и А 2. Ig. А синтезируется зрелыми B-лимфоцитами (Вα) и плазматическими клетками. Ig. А хорошо определяется в сыворотке крови на пике первичного и при вторичном иммунном ответе. Ig. А обладает высокой аффинностью, не связывает комплемент, не проходит через плацентарный барьер.

Секреторный Ig. А n n Секреторный Ig. А существует в форме ди- или тримера (4 - или 6 -валентный) и содержит J- и S-пептиды. Молекула секреторного Ig. А формируется при прохождении через эпителиальную клетку, где Ig. А присоединяется к секреторному компоненту. Молекулярная масса Ig. А 385 к. Да и более. Секреторный Ig. А – основной фактор местного иммунитета слизистых оболочек ЖКТ, мочеполовой системы и респираторного тракта. Этот иммуноглобулин препятствует адгезии микробов на эпителиальных клетках и генерализации инфекции в пределах слизистых оболочек. Секреторный Ig. А активирует комплемент и стимулирует фагоцитарную реакцию в слизистых оболочках.

Иммуноглобулин класса Е n n Ig. Е называют также реагином. Содержание его в сыворотке крови крайне невысоко — примерно 0, 00025 г/л. Обнаружить Ig. Е можно специальными высокочувствительными методами. Его молекулярная масса около 200 к. Да. Ig. Е — мономер. На его долю приходится около 0, 002 % всех циркулирующих иммуноглобулинов. Ig. Е синтезируется зрелыми B-лимфоцитами (Вε) и плазматическими клетками преимущественно в лимфоидной ткани бронхолегочного дерева и ЖКТ. Ig. Е не связывает комплемент, не проходит через плацентарный барьер. Он обладает выраженной тропностью к тучным клеткам и базофилам, участвует в развитии аллергической реакции I типа.

n n n Ig. E выявляется на мембране базофилов и тучных клеток у любого человека. Антигенсвязывающие Fabфрагменты молекулы Ig. E специфически взаимодействуют с АГ, попавшим в организм. Сформированный иммунный комплекс взаимодействует с рецепторами Fc-фрагментов Ig. E, встроенных в клеточную мембрану базофилов или тучных клеток. Это взаимодействие является сигналом для экзоцитоза гистамина, других биологически активных веществ и развертывания острой аллергической реакции. Кроме того, Ig. E сенсибилизированы клетки слизистых оболочек, в частности носовой полости, бронхов и конъюнктивы. Ig. E имеют существенное значение в антигельминтозном иммунитете

Иммуноглобулин класса D n n n Ig. D практически полностью содержится в сыворотке крови в концентрации около 0, 03 г/л (около 0, 2 % общего количества циркулирующих иммуноглобулинов). Ig. D имеет молекулярную массу около 180 к. Да, мономер. Ig. D не связывает комплемент, не проходит через плацентарный барьер, является рецептором предшественников B-лимфоцитов. Биологическая роль данного класса иммуноглобулинов до конца не известна. Предположительно он участвует в антиген-зависимой дифференцировке лимфоцитов.

Рецепторные или мембранные иммуноглобулины локализуются на цитоплазматической мембране Bлимфоцитов. Они выполняют функции антигенспецифических рецепторов. клетки. Нормальные антитела всегда определяются в сыворотке крови человека. Это антитела, направленные против эритроцитарных антигенов групп крови (система АВО), а также против бактерий кишечной группы, кокков и некоторых вирусов. Они постоянно образуются в организме без явной антигенной стимуляции. С одной стороны, они отражают готовность макроорганизма к иммунному реагированию, а с другой — могут свидетельствовать об отдаленном контакте с антигеном

Динамика антителопродукции. На появление во внутренней среде макроорганизма антигена иммунная система реагирует усилением биосинтеза специфических антител, что достигается путем размножения клонов антигенспецифичных клеток-антителопродуцентов. При этом антиген выступает в роли селектирующего фактора, и преимущества получают клоны с наивысшей к нему специфичностью, т. е. с наибольшей аффинностью рецепторных молекул иммуноглобулинов. Параллельно с размножением идет процесс дифференцировки B-лимфоцитов.
. Выделяют следующие фазы: латентную")
Антителопродукция в ответ на антигенный стимул имеет характерную динамику. ). Выделяют следующие фазы: латентную (индуктивную)(а), логарифмическую(б), стационарную(в) и снижения(г).

n n n В латентную фазу антителопродукция практически не изменяется и остается на базальном уровне. В этот период происходят переработка и представление антигена иммунокомпетентным клеткам, а также запуск пролиферации антигенспецифичных клонов клеток-антителопродуцентов. Ввиду того что клетки делятся дихотомически (т. е. надвое), их численность возрастает в геометрической прогрессии. Поэтому после первых циклов деления прирост числа клеток в общей массе невелик, и титры специфических антител практически не изменяются. Параллельно происходит созревание пре-Bлимфоцитов в зрелые формы, включаются процессы дифференцировки антителопродуцентов в плазматические клетки и переключение синтезируемых изотипов иммуноглобулинов. Во время логарифмической фазы наблюдается интенсивный прирост числа антигенспецифичных B-лимфоцитов, что находит отражение в существенном нарастании титров специфических антител. В стационарной фазе количество специфических антител и синтезирующих их клеток достигает максимума и стабилизируется. Освобождение макроорганизма от антигена устраняет антигенный стимул, поэтому вслед за стационарной фазой начинается фаза снижения. В этот период наблюдается постепенное уменьшение численности клонов специфических антителопродуцентов и титров соответствующих антител.
")
Динамика антителообразования имеет характерную временную зависимость. (Зависит от первичности или вторичности контакта с антигеном) При первичном контакте с антигеном развивается первичный иммунный ответ. Для него характерна длительная латентная (3 – 5 сут) и логарифмическая (7 – 15 сут) фазы. Первые диагностически значимые титры специфических антител регистрируются на 10 – 14 -е сутки от момента иммунизации. Стационарная фаза продолжается 15 – 30 сут, а фаза снижения – 1 – 6 мес. ' В течение первичного иммунного ответа созревают и размножаются клоны антигенспецифичных B-лимфоцитов. Их дифференцировка приводит к образованию плазматических клеток. Происходит также переключение биосинтеза иммуноглобулинов с изотипа М на А или G.

Повторный контакт иммунной системы с тем же антигеном ведет к формированию вторичного иммунного ответа. В отличие от первичного для вторичного ответа характерна: n Укороченная латентная фаза – от нескольких часов до 1 – 2 сут. n Логарифмическая фаза отличается более интенсивной динамикой прироста и более высокими титрами специфических антител. n Стационарной фазе и фазе снижения свойственна затяжная динамика (несколько месяцев или даже лет). При вторичном иммунном ответе в организме сразу же в подавляющем большинстве случаев синтезируется Ig. G. Характерная динамика антителопродукции обусловлена подготовленностью иммунной системы к повторной встрече с антигеном за счет формирования иммунологической памяти. В результате этого клоны антигенспецифичных B-лимфоцитов, оставшиеся после первичного иммунного реагирования, быстро размножаются и интенсивно включаются в процесс антителогенеза.
Антигены и антитела.ppt