

Лекция 11-3-дв.ан.ppt
- Количество слайдов: 51
 аксон мотонейрона двигательная единица спинной мозг аксон мотонейрона двигательная единица Лекция 11. Кожный и двигательный анализатор (Часть 3 - Проприорецепция). Мышечное веретено. Рецептор растяжения рака как модель механорецептора. Структура соматосенсорного анализатора. Нейронная организация спинного мозга. Соматотопическая проекция. Колончатая организация соматосенсорной коры.
аксон мотонейрона двигательная единица спинной мозг аксон мотонейрона двигательная единица Лекция 11. Кожный и двигательный анализатор (Часть 3 - Проприорецепция). Мышечное веретено. Рецептор растяжения рака как модель механорецептора. Структура соматосенсорного анализатора. Нейронная организация спинного мозга. Соматотопическая проекция. Колончатая организация соматосенсорной коры.
 В связи с усложнением движения у позвоночных развивается сложная система проприоцепции – мышечного и суставного чувства. Это чувство складывается из информации, поступающей от кожных, мышечных, суставных и сухожильных рецепторов, а также от рецепторов углового и линейного ускорения. Основную роль при этом играет информация, приходящая от мышц и сухожилий. Проприорецепция дает нам представление о расположении частей тела в пространстве и относительно друга; эта информация необходима для управления движениями.
В связи с усложнением движения у позвоночных развивается сложная система проприоцепции – мышечного и суставного чувства. Это чувство складывается из информации, поступающей от кожных, мышечных, суставных и сухожильных рецепторов, а также от рецепторов углового и линейного ускорения. Основную роль при этом играет информация, приходящая от мышц и сухожилий. Проприорецепция дает нам представление о расположении частей тела в пространстве и относительно друга; эта информация необходима для управления движениями.
,") Периферический отдел проприоцептивного анализатора представлен проприорецепторами (от лат. proprius – собственный, receptor – принимающий), расположенными в мышцах, связках, сухожилиях, суставных сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца Пачини, свободные нервные окончания. Поступление информации от мышц обеспечивается мышечными веретенами, а от сухожилий - сухожильными органами Гольджи.
Периферический отдел проприоцептивного анализатора представлен проприорецепторами (от лат. proprius – собственный, receptor – принимающий), расположенными в мышцах, связках, сухожилиях, суставных сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца Пачини, свободные нервные окончания. Поступление информации от мышц обеспечивается мышечными веретенами, а от сухожилий - сухожильными органами Гольджи.
 Мышечное веретено представляет собой скопление тонких коротких поперечнополосатых мышечных волокон, которые окружены соединительнотканной капсулой. Эти волокна получили название интрафузальных, в отличие от обычных мышечных волокон, которые составляют основную массу мышц и называются экстрафузальными, или рабочими, волокнами. Мышечное веретено с интрафузальными волокнами расположено параллельно экстрафузальным, поэтому возбуждаются при расслаблении (удлинении) скелетной мышцы.
Мышечное веретено представляет собой скопление тонких коротких поперечнополосатых мышечных волокон, которые окружены соединительнотканной капсулой. Эти волокна получили название интрафузальных, в отличие от обычных мышечных волокон, которые составляют основную массу мышц и называются экстрафузальными, или рабочими, волокнами. Мышечное веретено с интрафузальными волокнами расположено параллельно экстрафузальным, поэтому возбуждаются при расслаблении (удлинении) скелетной мышцы.
 Нервно-мышечное веретено — сложный рецептор, который включает видоизмененные мышечные клетки, афферентные и эфферентные нервные отростки. Контролирует скорость и степень сокращения и растяжение скелетных мышц. Выделяют два главных типа интрафузальных мышечных волокон: сумчато-ядерные и цепочечно-ядерные.
Нервно-мышечное веретено — сложный рецептор, который включает видоизмененные мышечные клетки, афферентные и эфферентные нервные отростки. Контролирует скорость и степень сокращения и растяжение скелетных мышц. Выделяют два главных типа интрафузальных мышечных волокон: сумчато-ядерные и цепочечно-ядерные.
 В полость каждого мышечного веретена проникают нервные волокна и кровеносные сосуды. Внутри мышечного веретена одиночное крупное волокно ветвится и посылает терминали дендритов к каждому интрафузальному мышечному волокну. Конечные нервные ветви обвиваются вокруг средних частей интрафузальных волокон, образуя так называемое аннулоспиральное окончание (лат. : anulus — колечко, завиток; anulo- в форме кольца). Афферентные нервные волокна, образующие аннулоспиральные окончания, принадлежат к группе Iа. Их называют афферентными волокнами мышечных веретен.
В полость каждого мышечного веретена проникают нервные волокна и кровеносные сосуды. Внутри мышечного веретена одиночное крупное волокно ветвится и посылает терминали дендритов к каждому интрафузальному мышечному волокну. Конечные нервные ветви обвиваются вокруг средних частей интрафузальных волокон, образуя так называемое аннулоспиральное окончание (лат. : anulus — колечко, завиток; anulo- в форме кольца). Афферентные нервные волокна, образующие аннулоспиральные окончания, принадлежат к группе Iа. Их называют афферентными волокнами мышечных веретен.
 Мышечные веретена состоят из иннервированных мышечных волокон, окруженных капсулой В состав мышечного веретена входит в среднем 2 -4 миниатюрных волокна скелетной мышцы, заключенных в капсулу соединительной ткани. Капсула обеспечивает механическую защиту элементов веретена, расположенных в полости капсулы, регулирует химический состав жидкой среды этих элементов.
Мышечные веретена состоят из иннервированных мышечных волокон, окруженных капсулой В состав мышечного веретена входит в среднем 2 -4 миниатюрных волокна скелетной мышцы, заключенных в капсулу соединительной ткани. Капсула обеспечивает механическую защиту элементов веретена, расположенных в полости капсулы, регулирует химический состав жидкой среды этих элементов.
 Любые интрафузальные мышечные волокна могут сокращаться за счет сократительных элементов, расположенных по их полюсам, где имеются нити актина и миозина. Центральные зоны интрафузальных мышечных волокон сократительных элементов не имеют. Напряжение сократительных элементов интрафузальных мышечных волокон, как и напряжение экстрафузальных мышечных волокон, управляется сигналами, поступающими к ним по эфферентным нервным волокнам от специализированных мотонейронов спинного мозга.
Любые интрафузальные мышечные волокна могут сокращаться за счет сократительных элементов, расположенных по их полюсам, где имеются нити актина и миозина. Центральные зоны интрафузальных мышечных волокон сократительных элементов не имеют. Напряжение сократительных элементов интрафузальных мышечных волокон, как и напряжение экстрафузальных мышечных волокон, управляется сигналами, поступающими к ним по эфферентным нервным волокнам от специализированных мотонейронов спинного мозга.
 Для каждой разновидности интрафузальных мышечных волокон существуют собственные регуляторы-мотонейроны и общие регуляторы для нескольких их разновидностей. Любой из двух сократительных элементов на полюсах одного интрафузального мышечного волокна может иметь собственную эфферентную иннервацию. Это обеспечивает возможность их раздельного независимого друг от друга сокращения.
Для каждой разновидности интрафузальных мышечных волокон существуют собственные регуляторы-мотонейроны и общие регуляторы для нескольких их разновидностей. Любой из двух сократительных элементов на полюсах одного интрафузального мышечного волокна может иметь собственную эфферентную иннервацию. Это обеспечивает возможность их раздельного независимого друг от друга сокращения.
 В каждой поперечно-полосатой мышце - от 6 до 1300 мышечных веретен. Их длина несколько миллиметров, диаметр — несколько десятых долей миллиметра.
В каждой поперечно-полосатой мышце - от 6 до 1300 мышечных веретен. Их длина несколько миллиметров, диаметр — несколько десятых долей миллиметра.
 Они воспринимают информацию об уровне процесса изменения длины мышцы, то есть они определяют среднее значение процесса. Выделяют две разновидности сумчатоядерных мышечных волокон: рецепторы с большим быстродействием, они реагируют на быстрые растяжения при сокращениях мышц измерение скорости растяжения мышцы. Другая разновидность рецепторы с малым быстродействием.
Они воспринимают информацию об уровне процесса изменения длины мышцы, то есть они определяют среднее значение процесса. Выделяют две разновидности сумчатоядерных мышечных волокон: рецепторы с большим быстродействием, они реагируют на быстрые растяжения при сокращениях мышц измерение скорости растяжения мышцы. Другая разновидность рецепторы с малым быстродействием.
 Цепочечно-ядерные мышечные волокна - являются рецепторами с малым быстродействием. Они воспринимают информацию об уровне процесса изменения длины мышцы, о среднем значении изменения длины мышцы - измерение силы растяжения мышцы
Цепочечно-ядерные мышечные волокна - являются рецепторами с малым быстродействием. Они воспринимают информацию об уровне процесса изменения длины мышцы, о среднем значении изменения длины мышцы - измерение силы растяжения мышцы


 Сухожильные органы Гольджи Сухожильный орган Гольджи, или нервно-сухожильное веретено — рецепторный орган, располагающийся в местах соединения мышечных волокон с коллагеновыми пучками сухожилий. Впервые сухожильные органы Гольджи появились в эволюции у первых наземных позвоночных. Сухожильный орган Гольджи. 1 -нервное волокно. 2 -сухожильные пучки. 3 -мышечное волокно. 4 -ветвления нервных волокон
Сухожильные органы Гольджи Сухожильный орган Гольджи, или нервно-сухожильное веретено — рецепторный орган, располагающийся в местах соединения мышечных волокон с коллагеновыми пучками сухожилий. Впервые сухожильные органы Гольджи появились в эволюции у первых наземных позвоночных. Сухожильный орган Гольджи. 1 -нервное волокно. 2 -сухожильные пучки. 3 -мышечное волокно. 4 -ветвления нервных волокон
 Сухожильный орган Гольджи состоит из коллагеновых нитей, отходящих примерно от 10 экстрафузальных мышечных волокон и заключенных в соединительнотканную капсулу. К ней подходят один или два толстых миелинизированных афферентных аксона (отростки сенсорных нейронов СМ). Войдя в капсулу, афферентные волокна разделяются на более тонкие отростки, утрачивают миелиновую оболочку и образуют сильно разветвленные окончания среди сухожильных нитей. Их окончания спирально закручиваются вокруг коллагеновых нитей. Скорость передачи импульса волокон — 80 -120 м/сек.
Сухожильный орган Гольджи состоит из коллагеновых нитей, отходящих примерно от 10 экстрафузальных мышечных волокон и заключенных в соединительнотканную капсулу. К ней подходят один или два толстых миелинизированных афферентных аксона (отростки сенсорных нейронов СМ). Войдя в капсулу, афферентные волокна разделяются на более тонкие отростки, утрачивают миелиновую оболочку и образуют сильно разветвленные окончания среди сухожильных нитей. Их окончания спирально закручиваются вокруг коллагеновых нитей. Скорость передачи импульса волокон — 80 -120 м/сек.
 Рецепторы Гольджи достигают у человека 2 – 3 мм в длину и 1 – 1, 5 мм в ширину. Они включены относительно скелетной мышцы последовательно, поэтому возбуждаются при ее сокращении вследствие натяжения сухожилия мышцы. Рецепторы Гольджи контролируют силу мышечнюго сокращения, т. е. напряжения.
Рецепторы Гольджи достигают у человека 2 – 3 мм в длину и 1 – 1, 5 мм в ширину. Они включены относительно скелетной мышцы последовательно, поэтому возбуждаются при ее сокращении вследствие натяжения сухожилия мышцы. Рецепторы Гольджи контролируют силу мышечнюго сокращения, т. е. напряжения.

 Органы Гольджи предотвращают повреждения мышцы при чрезмерной нагрузке Когда мышца сокращается, она растягивает сухожилие. Это растяжение деформирует концевые разветвления аксона, подходящему к органу Гольджи, что способствует открытию на мембране чувствительного нейрона катионных каналов. В результате аксон деполяризуется, и в нем, а затем и в остальных нервных окончаниях сенсорного нейрона возникает нервный импульс. Потом чувствительные нейроны возбуждают тормозные интернейроны спинного мозга, которые затем тормозят соответствующие двигательные нейроны, что предотвращает перерастяжение мышцы.
Органы Гольджи предотвращают повреждения мышцы при чрезмерной нагрузке Когда мышца сокращается, она растягивает сухожилие. Это растяжение деформирует концевые разветвления аксона, подходящему к органу Гольджи, что способствует открытию на мембране чувствительного нейрона катионных каналов. В результате аксон деполяризуется, и в нем, а затем и в остальных нервных окончаниях сенсорного нейрона возникает нервный импульс. Потом чувствительные нейроны возбуждают тормозные интернейроны спинного мозга, которые затем тормозят соответствующие двигательные нейроны, что предотвращает перерастяжение мышцы.
 Порог возбуждения рецепторов сухожильного органа Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен; это означает, что для возбуждения сухожильного органа Гольджи требуется чрезвычайно сильное растяжение или сокращение по сравнению с возбуждением мышечных веретен. Это позволяет мышце сокращаться без всяких помех до определенного предела. Активация органов Гольджи запускает спинномозговой рефлекс, расслабляющий мышцу
Порог возбуждения рецепторов сухожильного органа Гольджи при механическом растяжении мышцы выше, чем аналогичный порог возбуждения мышечных веретен; это означает, что для возбуждения сухожильного органа Гольджи требуется чрезвычайно сильное растяжение или сокращение по сравнению с возбуждением мышечных веретен. Это позволяет мышце сокращаться без всяких помех до определенного предела. Активация органов Гольджи запускает спинномозговой рефлекс, расслабляющий мышцу
 Рецептор растяжения рака как модель механорецептора Проприоцептивную информацию о взаиморасположении частей тела и состоянии двигательной системы у низших беспозвоночных дают механорецепторы находящиеся в основном в самих органах движения и в местах тела, где они отходят от него. У членистоногих такого рода рецепторы формируются в области кутикулярных сочленений и растяжения мышц. У раков и омаров такими рецепторами растяжения снабжены разгибатели брюшка.
Рецептор растяжения рака как модель механорецептора Проприоцептивную информацию о взаиморасположении частей тела и состоянии двигательной системы у низших беспозвоночных дают механорецепторы находящиеся в основном в самих органах движения и в местах тела, где они отходят от него. У членистоногих такого рода рецепторы формируются в области кутикулярных сочленений и растяжения мышц. У раков и омаров такими рецепторами растяжения снабжены разгибатели брюшка.
. Этот препарат удобен") Рецепторы растяжения речного рака (кодирование стимула было проанализировано Эйзагиром и Куффлером). Этот препарат удобен тем, что клеточное тело рецептора растяжения находится не в ганглии, а расположено изолированно на периферии, где его можно видеть в живом препарате. Клетка достаточно велика для введения внутриклеточных микроэлектродов. Она проникает своими дендритами в ближайший тонкий мышечный тяж, а свой аксон посылает в сегментарный ганглий. Схематическое изображение рецептора растяжения ракообразного
Рецепторы растяжения речного рака (кодирование стимула было проанализировано Эйзагиром и Куффлером). Этот препарат удобен тем, что клеточное тело рецептора растяжения находится не в ганглии, а расположено изолированно на периферии, где его можно видеть в живом препарате. Клетка достаточно велика для введения внутриклеточных микроэлектродов. Она проникает своими дендритами в ближайший тонкий мышечный тяж, а свой аксон посылает в сегментарный ганглий. Схематическое изображение рецептора растяжения ракообразного
 Рецепторный потенциал и потенциалы действия в рецепторе растяжения А. Схематическое изображение рецептора растяжения ракообразного. Стимул деполяризует дендриты, вызывая рецепторный потенциал, который регистрируется в соме. Б. Ритмические потенциалы действия, которые возникают в основании аксона под влиянием рецепторного потенциала.
Рецепторный потенциал и потенциалы действия в рецепторе растяжения А. Схематическое изображение рецептора растяжения ракообразного. Стимул деполяризует дендриты, вызывая рецепторный потенциал, который регистрируется в соме. Б. Ритмические потенциалы действия, которые возникают в основании аксона под влиянием рецепторного потенциала.
 У ракообразных существует два типа рецепторов растяжения, различающихся своими структурными и физиологическими свойствами, а также тем, что они своими дендритами иннервируют различные типы мышц. Одни из них бурно активируются в начале растяжения, но затем их реакция быстро ослабевает. Такое снижение ответной реакции в процессе действия стимула постоянной интенсивности называется адаптацией. Второй тип называется медленно адаптирующимся: его реакция сохраняется неизменной во время продолжительного растяжения.
У ракообразных существует два типа рецепторов растяжения, различающихся своими структурными и физиологическими свойствами, а также тем, что они своими дендритами иннервируют различные типы мышц. Одни из них бурно активируются в начале растяжения, но затем их реакция быстро ослабевает. Такое снижение ответной реакции в процессе действия стимула постоянной интенсивности называется адаптацией. Второй тип называется медленно адаптирующимся: его реакция сохраняется неизменной во время продолжительного растяжения.
 Специализированные рецепторы возникли для точной координации движений и анализа положения тела. У некоторых рыб в плавниковых мышцах имеются медленно адаптирующиеся механорецепторы, которые сигнализируют об их тоническом напряжении. Располагаются они не в самих мышечных волокнах, а в окружающей их соединительной ткани. У амфибий впервые появляются мышечные веретена. Они включают видоизмененные мышечные волокна, на которых располагается плоское или спиральное нервное окончание. Последнее посылает в мозг быстрые сигналы при удлинении мышцы.
Специализированные рецепторы возникли для точной координации движений и анализа положения тела. У некоторых рыб в плавниковых мышцах имеются медленно адаптирующиеся механорецепторы, которые сигнализируют об их тоническом напряжении. Располагаются они не в самих мышечных волокнах, а в окружающей их соединительной ткани. У амфибий впервые появляются мышечные веретена. Они включают видоизмененные мышечные волокна, на которых располагается плоское или спиральное нервное окончание. Последнее посылает в мозг быстрые сигналы при удлинении мышцы.
 У птиц и млекопитающих веретена состоят из нескольких мышечных волокон с первичными быстро реагирующими и вторичными медленно реагирующими гроздевидными рецепторами, формирующими сигналы как при сжатии, так и при растяжении мышцы. Мышечный рецептор млекопитающих
У птиц и млекопитающих веретена состоят из нескольких мышечных волокон с первичными быстро реагирующими и вторичными медленно реагирующими гроздевидными рецепторами, формирующими сигналы как при сжатии, так и при растяжении мышцы. Мышечный рецептор млекопитающих
 Для точной координации движений и анализа положения тела возникли разнообразные суставные и сухожильные рецепторы. В соединительнотканной капсуле суставов имеется много специализированных рецепторов, напоминающих немного модифицированные кожные рецепторы типа телец Руффини, и Пачини. Кроме того, в суставных связках присутствуют немиелинизированные свободные нервные окончания. В сухожилиях имеются миелинизированные рецепторы Гольджи с медленной адаптацией к растяжению-сжатию. Обильная иннервация суставов характерна только для млекопитающих и птиц. У низших позвоночных как сами суставные сумки, так и связки практически не иннервированы, а суставные рецепторы расположены в окружающей соединительной ткани.
Для точной координации движений и анализа положения тела возникли разнообразные суставные и сухожильные рецепторы. В соединительнотканной капсуле суставов имеется много специализированных рецепторов, напоминающих немного модифицированные кожные рецепторы типа телец Руффини, и Пачини. Кроме того, в суставных связках присутствуют немиелинизированные свободные нервные окончания. В сухожилиях имеются миелинизированные рецепторы Гольджи с медленной адаптацией к растяжению-сжатию. Обильная иннервация суставов характерна только для млекопитающих и птиц. У низших позвоночных как сами суставные сумки, так и связки практически не иннервированы, а суставные рецепторы расположены в окружающей соединительной ткани.
 Переработку соматосенсорной информации в ЦНС можно разделить на три уровня: афферентный, интегративный и эфферентный. Афферентная подсистема включает в себя передачу и обработку сигналов в спинном мозге, стволе, таламусе, и коре больших полушарий. Интегративный уровень связан с ассоциативной и лимбической системами головного мозга; в первую очередь, он нужен для того, чтобы соотнести вновь поступившие сигналы с теми, которые хранятся у нас в памяти. Эфферентный уровень включает в себя двигательную и вегетативную системы. Его функция – обеспечить реакцию. В состав этой подсистемы входят несколько путей, передающих информацию параллельно и запускающие реакцию организма на поступающую информацию с учетом всех факторов.
Переработку соматосенсорной информации в ЦНС можно разделить на три уровня: афферентный, интегративный и эфферентный. Афферентная подсистема включает в себя передачу и обработку сигналов в спинном мозге, стволе, таламусе, и коре больших полушарий. Интегративный уровень связан с ассоциативной и лимбической системами головного мозга; в первую очередь, он нужен для того, чтобы соотнести вновь поступившие сигналы с теми, которые хранятся у нас в памяти. Эфферентный уровень включает в себя двигательную и вегетативную системы. Его функция – обеспечить реакцию. В состав этой подсистемы входят несколько путей, передающих информацию параллельно и запускающие реакцию организма на поступающую информацию с учетом всех факторов.
 Афферентные пути Существует несколько параллельных путей передачи") Схема лемнискового пути (проводящие тракты обозначены синим) Афферентные пути Существует несколько параллельных путей передачи сигнала от рецепторов тела в головной мозг. Основные пути - лемнисковый и спинно-таламический. Они сильно различаются как по строению, так и по функциям. Кроме того, есть несколько побочных, менее важных восходящих путей.
Схема лемнискового пути (проводящие тракты обозначены синим) Афферентные пути Существует несколько параллельных путей передачи сигнала от рецепторов тела в головной мозг. Основные пути - лемнисковый и спинно-таламический. Они сильно различаются как по строению, так и по функциям. Кроме того, есть несколько побочных, менее важных восходящих путей.
 Лемнисковый путь. Главная функция этого пути – быстрая и точная передача информации от тактильных рецепторов и проприорецепторов. Его проводящие волокна относительно толстые и миелинизированы на всех уровнях. Первые нейроны лемнискового пути находятся в спинальных ганглиях, их аксоны входят в состав задних столбов. Затем они образуют синапсы с нейронами в продолговатом мозге. Часть аксонов (в основном, от рецепторов мышц и суставов) входит в состав побочных путей или же оканчивается на мотонейронах, запуская спинномозговые рефлексы.
Лемнисковый путь. Главная функция этого пути – быстрая и точная передача информации от тактильных рецепторов и проприорецепторов. Его проводящие волокна относительно толстые и миелинизированы на всех уровнях. Первые нейроны лемнискового пути находятся в спинальных ганглиях, их аксоны входят в состав задних столбов. Затем они образуют синапсы с нейронами в продолговатом мозге. Часть аксонов (в основном, от рецепторов мышц и суставов) входит в состав побочных путей или же оканчивается на мотонейронах, запуская спинномозговые рефлексы.
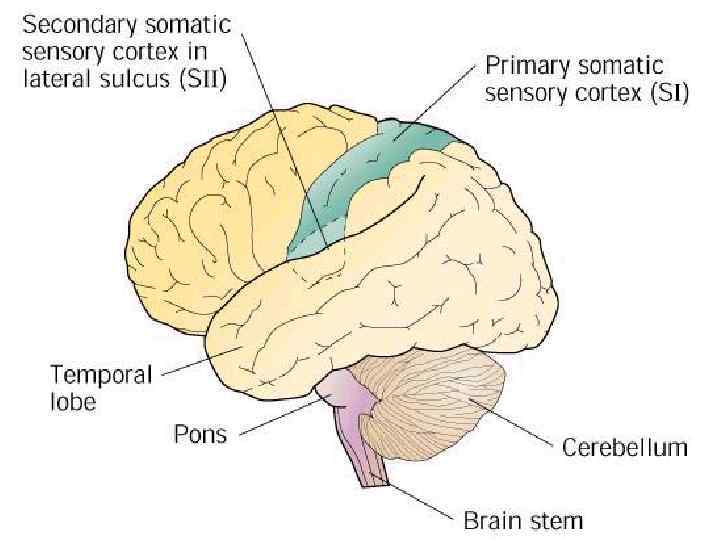
 Из продолговатого мозга, от вторых нейронов лемнискового пути, аксоны идут в таламус, при этом перекрещиваясь и совершая медиальную петлю – резкий изгиб пучка нервных волокон в стволе мозга (по-другому она называется медиальный лемниск, откуда и пошло название пути). В таламусе происходит передача сигнала на третьи нейроны, расположенные в специальных ядрах – вентробазальном ядерном комплексе. От них аксоны направляются в соматосенсорную кору головного мозга. Там она проецируется на соматосенсорную кору, причем сигналы от близлежащих рецепторов проецируются на соседние участки коры. Такая организация передачи сигнала называется соматотопической, т. е. отражающей взаимное положение иннервируемых участков.
Из продолговатого мозга, от вторых нейронов лемнискового пути, аксоны идут в таламус, при этом перекрещиваясь и совершая медиальную петлю – резкий изгиб пучка нервных волокон в стволе мозга (по-другому она называется медиальный лемниск, откуда и пошло название пути). В таламусе происходит передача сигнала на третьи нейроны, расположенные в специальных ядрах – вентробазальном ядерном комплексе. От них аксоны направляются в соматосенсорную кору головного мозга. Там она проецируется на соматосенсорную кору, причем сигналы от близлежащих рецепторов проецируются на соседние участки коры. Такая организация передачи сигнала называется соматотопической, т. е. отражающей взаимное положение иннервируемых участков.
 Разные участки кожи иннервированы поразному, на них отводятся разные по размеру части коры - получается проекция с сохраненным местоположением частей тела относительно друга, но с искаженными размерами (визуальное изображение распределения информации в соматосенсорной коре – т. н. сенсорный гомункулус). Искаженное геометрически проецирование характерно не только для коры, но и для других уровней лемнискового пути.
Разные участки кожи иннервированы поразному, на них отводятся разные по размеру части коры - получается проекция с сохраненным местоположением частей тела относительно друга, но с искаженными размерами (визуальное изображение распределения информации в соматосенсорной коре – т. н. сенсорный гомункулус). Искаженное геометрически проецирование характерно не только для коры, но и для других уровней лемнискового пути.



 Нейронная организация спинного мозга Серое вещество распределено на ядра, вытянутые по длине спинного мозга, и на поперечном разрезе располагается в форме буквы Н. В грудном отделе спинной мозг имеет еще и боковые рога. Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга. В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов.
Нейронная организация спинного мозга Серое вещество распределено на ядра, вытянутые по длине спинного мозга, и на поперечном разрезе располагается в форме буквы Н. В грудном отделе спинной мозг имеет еще и боковые рога. Задние рога выполняют главным образом сенсорные функции и содержат нейроны, передающие сигналы в вышележащие центры, в симметричные структуры противоположной стороны либо к передним рогам спинного мозга. В передних рогах находятся нейроны, дающие свои аксоны к мышцам. Все нисходящие пути центральной нервной системы, вызывающие двигательные реакции, заканчиваются на нейронах передних рогов.
 мотонейроны, или двигательные,") Функционально нейроны спинного мозга можно разделить на 4 основные группы: 1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки; 2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения; 3) симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков; 4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами.
Функционально нейроны спинного мозга можно разделить на 4 основные группы: 1) мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки; 2) интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах. Эти нейроны реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения; 3) симпатические, парасимпатические нейроны расположены преимущественно в боковых рогах. Аксоны этих нейронов выходят из спинного мозга в составе передних корешков; 4) ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами.
 спинного мозга имеется промежуточное") В средней зоне серого вещества (между задним и передним рогами) спинного мозга имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1— 2 сегмента и дают коллатерали на нейроны ипси- и контралатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога спинного мозга — эта сеть образует так называемое студенистое вещество (желатинозная субстанция Роланда) и выполняет функции ретикулярной формации спинного мозга.
В средней зоне серого вещества (между задним и передним рогами) спинного мозга имеется промежуточное ядро (ядро Кахаля) с клетками, аксоны которых идут вверх или вниз на 1— 2 сегмента и дают коллатерали на нейроны ипси- и контралатеральной стороны, образуя сеть. Подобная сеть имеется и на верхушке заднего рога спинного мозга — эта сеть образует так называемое студенистое вещество (желатинозная субстанция Роланда) и выполняет функции ретикулярной формации спинного мозга.
 Мотонейроны. Аксон мотонейрона своими терминалами иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Мотонейроны спинного мозга функционально делят на α- и γ-нейроны. α-мотонейроны образуют связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена. γ-мотонейроны иннервируют интрафузальные мышечные волокна мышечного веретена, получают информацию о его состоянии через промежуточные нейроны.
Мотонейроны. Аксон мотонейрона своими терминалами иннервирует сотни мышечных волокон, образуя мотонейронную единицу. Мотонейроны спинного мозга функционально делят на α- и γ-нейроны. α-мотонейроны образуют связи с чувствительными путями, идущими от экстрафузальных волокон мышечного веретена. γ-мотонейроны иннервируют интрафузальные мышечные волокна мышечного веретена, получают информацию о его состоянии через промежуточные нейроны.
 В случаях поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх.
В случаях поражения заднего рога утрачивается болевая и температурная чувствительность на стороне повреждения, а тактильная и проприоцептивная сохраняется, так как из заднего корешка аксоны температурной и болевой чувствительности идут в задний рог, а аксоны тактильной и проприоцептивной — прямо в задний столб и по проводящим путям поднимаются вверх.
 Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность.
Вследствие того, что аксоны вторых нейронов болевой и температурной чувствительности идут на противоположную сторону через переднюю серую спайку спинного мозга, при повреждении этой спайки на теле симметрично утрачивается болевая и температурная чувствительность.
 Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом.
Поражение переднего рога и переднего корешка спинного мозга приводит к параличу мышц, которые теряют тонус, атрофируются, при этом исчезают рефлексы, связанные с пораженным сегментом.


 The proprioceptive system
The proprioceptive system


 Колончатая организация соматосенсорной коры Согласно современным представлениям, соматосенсорная кора организована в элементарные функциональные единицы – колонки. Каждая колонка состоит из нескольких морфологических микромодулей, объединяющих пять-шесть гнездообразно расположенных нейронов. В его состав входит несколько пирамидных клеток, апикальные дендриты которых максимально сближены и образуют дендритный пучок; в пределах этого пучка возможны электротонические связи, которые обеспечивают, по всей вероятности, синхронную работу всего объединения. Структурный модуль сенсомоторной коры: III-YI -корковые слои; 1 -интернейроны, 2 - афферентный вход, контактирующий с интернейроном, 3 - афферентный вход, образующий терминали на пирамидных нейронах, 4 - возвратная коллатераль аксона, вступающая в контакт с тормозным интернейроном, 5 -пучок аксонов, выходящий за пределы колонки, 6 - возвратная коллатераль, обеспечивающая облегчающие влияния в пределах модуля, 7 - кортикоспинальные пирамидные клетки, 8 - кортикорубральная пирамидная клетка.
Колончатая организация соматосенсорной коры Согласно современным представлениям, соматосенсорная кора организована в элементарные функциональные единицы – колонки. Каждая колонка состоит из нескольких морфологических микромодулей, объединяющих пять-шесть гнездообразно расположенных нейронов. В его состав входит несколько пирамидных клеток, апикальные дендриты которых максимально сближены и образуют дендритный пучок; в пределах этого пучка возможны электротонические связи, которые обеспечивают, по всей вероятности, синхронную работу всего объединения. Структурный модуль сенсомоторной коры: III-YI -корковые слои; 1 -интернейроны, 2 - афферентный вход, контактирующий с интернейроном, 3 - афферентный вход, образующий терминали на пирамидных нейронах, 4 - возвратная коллатераль аксона, вступающая в контакт с тормозным интернейроном, 5 -пучок аксонов, выходящий за пределы колонки, 6 - возвратная коллатераль, обеспечивающая облегчающие влияния в пределах модуля, 7 - кортикоспинальные пирамидные клетки, 8 - кортикорубральная пирамидная клетка.
 Аксоны пирамидных клеток образуют возвратные коллатерали, которые могут обеспечивать и облегчающие влияния в пределах микромодуля, и тормозные взаимодействия между микромодулями, контактируя с тормозными интернейронами. К группе пирамидных клеток примыкают звездчатые клетки, с которыми контактируют приходящие таламокортикальные волокна. Некоторые из звездчатых клеток, выполняющие тормозную функцию, имеют длинные аксоны, распространяющиеся в горизонтальном направлении. Несколько структурных микромодулей, аксоны терминалей и отростков звездчатых клеток формируют колонку (или макромодуль), диаметр которой достигает 500 -1000 мкм. Каждая колонка характеризуется функциональным единством, которое проявляется в том, что нейроны колонки отвечают на раздражитель одной модальности.
Аксоны пирамидных клеток образуют возвратные коллатерали, которые могут обеспечивать и облегчающие влияния в пределах микромодуля, и тормозные взаимодействия между микромодулями, контактируя с тормозными интернейронами. К группе пирамидных клеток примыкают звездчатые клетки, с которыми контактируют приходящие таламокортикальные волокна. Некоторые из звездчатых клеток, выполняющие тормозную функцию, имеют длинные аксоны, распространяющиеся в горизонтальном направлении. Несколько структурных микромодулей, аксоны терминалей и отростков звездчатых клеток формируют колонку (или макромодуль), диаметр которой достигает 500 -1000 мкм. Каждая колонка характеризуется функциональным единством, которое проявляется в том, что нейроны колонки отвечают на раздражитель одной модальности.
 В пределах одной колонки нейроны можно дифференцировать по характеру их импульсных ответов на простые и сложные. У простых нейронов распределение импульсов в ответе на раздражение адекватного механорецептора идентично ответу самого рецептора. Сложные нейроны дают максимальный ответ только на определенные формы раздражения, например на движущиеся стимулы. По всей вероятности, сложные нейроны связаны с более поздней ступенью анализа тактильного восприятия. Нейрофизиологические исследования сенсомоторной коры показали, что обычно возбуждение одного из модулей сопровождается торможением соседних. Очевидно, этот факт свидетельствует о том, что процесс торможения препятствует иррадиации возбуждения на смежные модули и, таким образом, усиливает контрастное различение контуров тактильного раздражения.
В пределах одной колонки нейроны можно дифференцировать по характеру их импульсных ответов на простые и сложные. У простых нейронов распределение импульсов в ответе на раздражение адекватного механорецептора идентично ответу самого рецептора. Сложные нейроны дают максимальный ответ только на определенные формы раздражения, например на движущиеся стимулы. По всей вероятности, сложные нейроны связаны с более поздней ступенью анализа тактильного восприятия. Нейрофизиологические исследования сенсомоторной коры показали, что обычно возбуждение одного из модулей сопровождается торможением соседних. Очевидно, этот факт свидетельствует о том, что процесс торможения препятствует иррадиации возбуждения на смежные модули и, таким образом, усиливает контрастное различение контуров тактильного раздражения.