IV_lektsia_Adaptivny_immunitet.pptx
- Количество слайдов: 33



Адаптивный иммунитет Доцент И. В. Беляева
, селекция лимфоцитов. Костный мозг – продукция")
Центральные лимфоидные органы: пролиферация, дифференцировка (так называемая, антиген-независимая), селекция лимфоцитов. Костный мозг – продукция В-лимфоцитов. Тимус – развитие Т-лимфоцитов. Далее лимфоциты выселяются в периферические лимфоидные органы, постоянно рециркулируя: ткани – лимфа – лимфоидные ткани – кровь. При активации антигеном – клональная антигензависимая пролиферация и дифференцировка.

Т-ЛИМФОЦИТЫ Основная задача T-лимфоцитов - распознавание чужеродных или изменённых собственных антигенов в составе комплекса с молекулами MHC. Если на поверхности своих клеток представлены чужеродные или изменённые свои молекулы, T-лимфоцит запускает их уничтожение. В отличие от B-лимфоцитов, T-лимфоциты не продуцируют растворимых форм антигенраспознающих молекул (антител). Более того, большинство T-лимфоцитов не способны распознавать растворимые антигены.

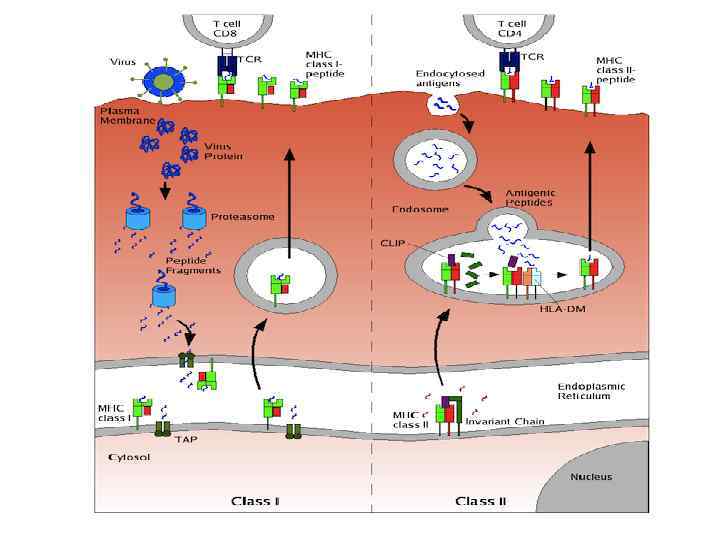
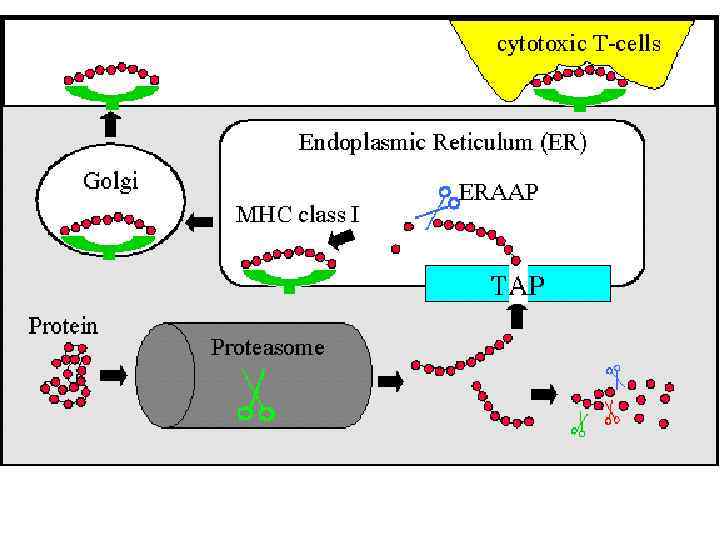
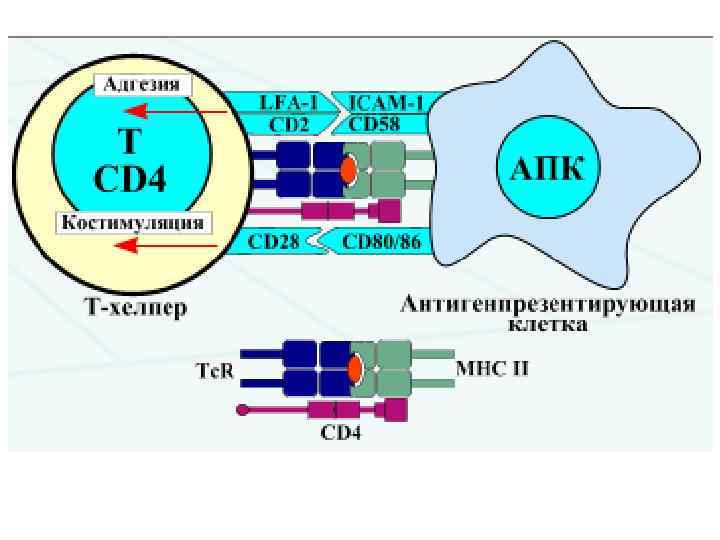
Чтобы T-лимфоцит «обратил на антиген своё внимание» , другие клетки должны «пропустить» антиген через себя и выставить его на своей мембране в комплексе с MHC-I или MHC-II. Это и есть процессинг и презентация антигена Tлимфоциту. Распознавание такого комплекса Tлимфоцитом - двойное распознавание, «своего» - молекул МНС (MHC-рестрикция Tлимфоцитов) и «чужого» – процессированного антигена (короткого пептидного фрагмента).
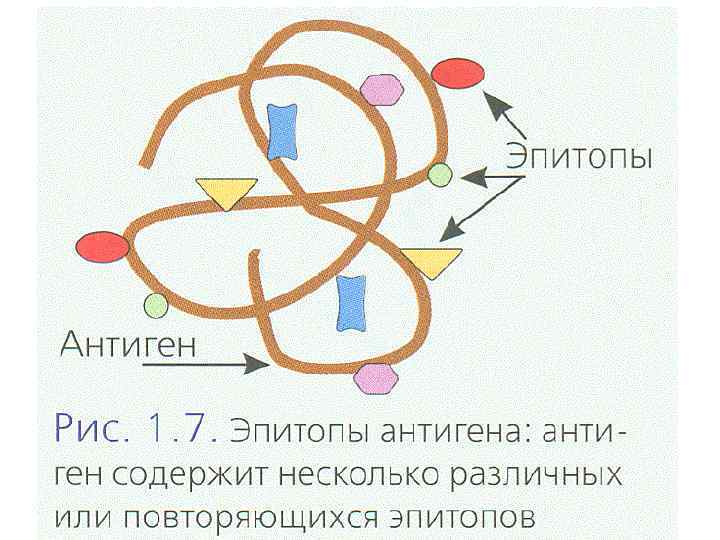

АНТИГЕНРАСПОЗНАЮЩИЙ РЕЦЕПТОР T-ЛИМФОЦИТОВ Антигенраспознающие рецепторы T-клеток - TCR состоят из цепей, принадлежащих к суперсемейству иммуноглобулинов

Антигенраспознающий участок TCR - гетеродимер, т. е. состоит из двух разных полипептидных цепей. Известны два варианта TCR, обозначаемые как αβTCR и γδTCR, они различаются составом полипептидных цепей антигенраспознающего участка. αβT-клетки были открыты раньше и изучены подробнее, чем γδT-лимфоциты. В связи с этим строение антигенраспознающего рецептора T-лимфоцитов удобнее описывать на примере αβTCR. Трансмембранно расположенный комплекс TCR состоит из 8 полипептидных цепей (α- и β-цепей собственно TCR, двух вспомогательных цепей ζ, а также по одному гетеродимеру ε/δ- и ε/γ-цепей молекулы СD 3)

Короткий цитоплазматический участок α- и β-цепи не может проводить сигнал внутрь клетки. Для этого служат 6 дополнительных полипептидных цепей: γ, δ, 2ε и 2ζ. • Комплекс CD 3. Цепи γ, δ, ε между собой образуют гетеродимеры γε и δε (вместе их называют комплекс CD 3). Этот комплекс необходим для экспрессии α- и β -цепей, их стабилизации и проведения сигнала внутрь клетки. • ζ-Цепи расположены в цитоплазме и осуществляют проведение сигнала внутрь клетки.

Корецепторные молекулы CD 4 и CD 8 • CD 4 связывается с инвариантной частью (β 2 доменом) молекулы MHC-II (принадлежит к суперсемейству иммуноглобулинов). При активации T-лимфоцита одну молекулу TCR «обслуживают» 2 молекулы CD 4: вероятно, происходит димеризация молекул CD 4.

• CD 8 принадлежит к суперсемейству иммуноглобулинов, связывается с инвариантной частью молекулы MHC-I. CD 8 - гетеродимер цепей α и β, соединённых дисульфидной связью. Во внеклеточной части каждая из цепей имеет по одному иммуноглобулиноподобному домену.

Гены T-клеточного рецептора Гены α-, β-, γ- и δ-цепей гомологичны генам иммуноглобулинов и претерпевают при дифференцировке T-лимфоцитов рекомбинацию ДНК, что теоретически обеспечивает генерацию порядка 1016 -1018 вариантов антигенсвязывающих рецепторов (реально это разнообразие ограничено числом лимфоцитов в организме до 109). Рекомбинация ДНК происходит при объединении V-, D- и J-сегментов и катализируется тем же комплексом рекомбиназ, что и при дифференцировке Bлимфоцитов.
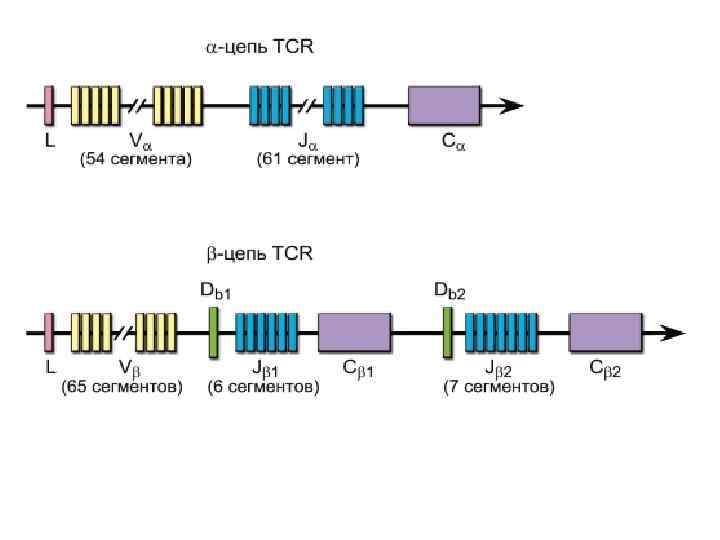

После перестройки VJ в генах α-цепи и VDJ в генах βцепи, а также после присоединения некодируемых N- и P-нуклеотидов с ДНК транскрибируется РНК. Гены α-цепи могут перестраиваться неоднократно при уже правильно перестроенных и экспрессированных генах β-цепи. Именно поэтому есть вероятность того, что одна клетка может нести более одного варианта TCR. • Соматическому гипермутагенезу гены TCR в отличие от генов иммуноглобулинов не подвергаются.

Развитие Т-лимфоцитов • Клетки-предшественники Т-лц мигрируют из костного мозга в субкапсулярную зону коры тимуса, где пролиферируют. • Затем клетки перемещаются вглубь коры тимуса. Начинается перестройка генов β-, γ- и δ-цепей TCR. Если гены γ- и δ-цепей успевают перестроиться раньше, чем гены β-цепи, то лимфоцит дифференцируется далее как γδT. В противном случае развивается αβТ. Клетки начинают пролиферировать. Накапливается масса клеток с уже готовой β-цепью, но с ещё не перестроенными генами α-цепи, что вносит свой вклад в разнообразие αβгетеродимеров. • Далее - рекомбинация вариабельных сегментов генов αцепи.

Селекция тимоцитов - Позитивная селекция: погибают тимоциты, не связавшие ни одного из доступных комплексов MHCпептид (не имеют CD 4 или CD 8). В результате позитивной селекции в тимусе погибает около 90% тимоцитов. - Негативная селекция уничтожает клоны тимоцитов, связывающих комплексы MHC-пептид со слишком высокой аффинностью (распознан «свой» пептид). - Тимоциты, связавшие какой-либо из комплексов MHC -пептид с правильной, т. е. средней по силе, аффинностью, получают сигнал к выживанию и продолжают дифференцировку.

 - они распознают и непосредственно убивают")
CD 8+ T-клетки играют роль цитотоксических Tлимфоцитов (ЦТЛ) - они распознают и непосредственно убивают клетки, модифицированные вирусом, опухолевые и другие «изменённые» клетки. CD 4+ T-клетки. Функциональная специализация CD 4+ Tлимфоцитов более разнообразна. Значительная часть CD 4+ T-лимфоцитов в процессе развития иммунного ответа становится T-хелперами (помощниками), взаимодействующими с В-лимфоцитами, Тлимфоцитами и другими клетками прямом контакте или через растворимые факторы (цитокины). В определённых случаях из них могут развиться CD 4+ ЦТЛ.

Как CD 8+ T-клетки убивают

Субпопуляции T-хелперов С конца 80 -х годов XX века было принято выделять 2 субпопуляции T-хелперов (в зависимости от того, какой набор цитокинов они продуцируют) - Th 1 и Th 2. В последние годы спектр субпопуляций CD 4+ Т-клеток продолжает расширяться. Обнаружены такие субпопуляции, как: Th 17, T-регуляторы, Tr 1, Th 3, Tfh и др.

• Th 0 - CD 4+ Т-лимфоциты на ранних стадиях развития иммунного ответа, они продуцируют только ИЛ-2 (митоген для всех лимфоцитов). • Th 1 - дифференцированная субпопуляция CD 4+ Тлимфоцитов (продукция ИФНγ, ФНОβ и ИЛ-2). Регуляция многих реакций клеточного иммунитета, включая гиперчувствительность замедленного типа (ГЗТ) и активацию ЦТЛ. Кроме того, Th 1 стимулируют продукцию В-лимфоцитами опсонизирующих антител класса Ig. G, запускающих каскад активации комплемента. Развитие избыточного воспаления с последующим повреждением тканей напрямую связано с активностью Th 1 -субпопуляции.

• Th 2 - дифференцированная субпопуляция CD 4+ Т-лимфоцитов, специализирующаяся на выработке ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13. Эта субпопуляция участвует в активации Влимфоцитов и способствует секреции ими больших количеств антител разных классов, особенно Ig. E. Кроме того, Th 2 -субпопуляция участвует в активации эозинофилов и развитии аллергических реакций.
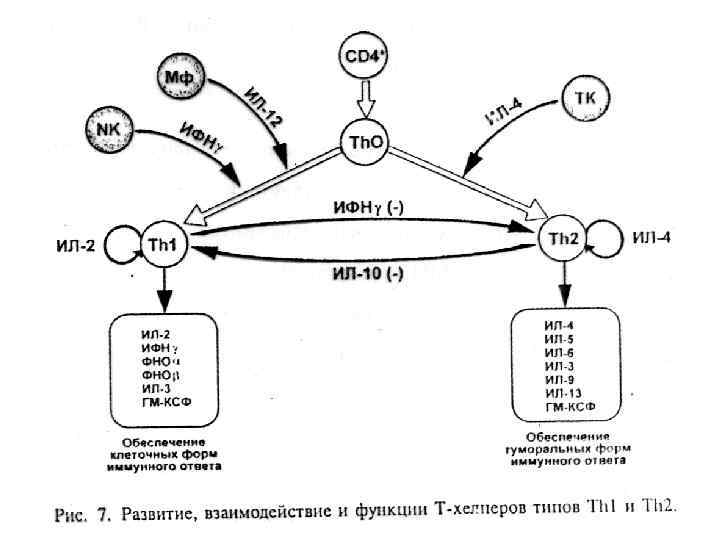

Th 17 - субпопуляция CD 4+ Т-лимфоцитов, специализирующаяся на образовании ИЛ-17. Эти клетки осуществляют противогрибковую и антимикробную защиту эпителиальных и слизистых барьеров, а также играют ключевую роль в патологии аутоиммунных заболеваний. В последнее время появляется описание всё новых популяций CD 4+ Т-лимфоцитов, классифицированных по типу преимущественно продуцируемого ими цитокина: Th 18, Th 9, Th 23
 находятся в лимфоидных фолликулах и осуществляют хелперную")
Tfh ( follicular helper - фолликулярный хелпер) находятся в лимфоидных фолликулах и осуществляют хелперную функцию для Влимфоцитов, секретируя ИЛ-21, вызывая их дифференцировку в плазматические клетки. Кроме ИЛ-21 Tfh могут также продуцировать ИЛ-6 и ИЛ-10, необходимые для дифференцировки В-лимфоцитов. Нарушение функций этой популяции приводит к развитию аутоиммунных заболеваний или иммунодефицитов.

Т-регуляторы - CD 4+ Т-лимфоциты, подавляющие активность других клеток иммунной системы: секретируя иммуносупрессорные цитокины - ИЛ-10 (ингибитор активности макрофагов и Th 1 -клеток) и ТФРβ - ингибитор пролиферации лимфоцитов. Ингибиторный эффект может также достигаться индукцией апоптоза. CD 4+ регуляторные Т-лимфоциты: естественные (Treg), созревающие в тимусе (CD 4+CD 25+, экспрессируют фактор транскрипции Foxp 3), и индуцированные - локализованные преимущественно в слизистых оболочках пищеварительного тракта и переключившиеся на образование ТФРβ (Th 3) или ИЛ-10 (Tr 1). Нормальное функционирование Т-регуляторов предотвращает развитие аутоиммунных заболеваний.
")
Адаптивные субпопуляции Т-клеток (клетки-партнеры, физиологические и патологические эффекты)
 T-лимфоцитов, проходящих лимфопоэз в тимусе, составляют αβTклетки; менее 1% -")
γδT-лимфоциты Подавляющее большинство (99%) T-лимфоцитов, проходящих лимфопоэз в тимусе, составляют αβTклетки; менее 1% - γδT-клетки. Последние в большинстве дифференцируются вне тимуса, в первую очередь в слизистых оболочках пищеварительного тракта. В коже, лёгких, пищеварительном и репродуктивном трактах они являются доминирующей субпопуляцией внутриэпителиальных лимфоцитов. Среди всех Tлимфоцитов организма γδT-клетки составляют от 10 до 50%. В эмбриогенезе γδT-клетки появляются раньше αβT-клеток.

γδT-клетки • не экспрессируют CD 4. • связывают нативные антигены независимо от классических молекул MHC - для них не обязателен или вовсе не нужен предварительный процессинг антигена АПК. • Разнообразие γδTCR меньше, чем αβTCR или иммуноглобулинов, γδT-клетки способны распознавать в основном фосфолипидные антигены микобактерий, углеводы, белки теплового шока.
 и тканей является Аг-неспецифической и")
Толерантность на уровне периферических органов иммуногенеза (миндалины, лимфатические узлы) и тканей является Аг-неспецифической и обеспечивается следующими путями: а) Клональная инактивация зрелых Т-клеток: если Тлимфоцит распознал Аг, но не получил костимуляторных сигналов от Аг-презентирующей клетки, то он инактивируется, т. е. переходит в состояние анергии. В норме аутореактивные Т-лимфоциты распознают ауто. Аг, которые в большом количестве экспрессируются в тканях, но не получают костимуляции и «обезвреживаются» . Таким же образом В-лимфоцит без активации Т-хелперами также переходит в состояние анергии.
 Супрессорный эффект Т-хелперов. Классическая модель баланса Т-супрессоров и Тхелперов была отвергнута несколько лет")
б) Супрессорный эффект Т-хелперов. Классическая модель баланса Т-супрессоров и Тхелперов была отвергнута несколько лет назад. Сейчас считается, что принципиальным является баланс между Т-хелперами 1 и 2 типов. Т-хелперы 2 типа подавляют функцию аутореактивных Тхелперов 1 типа путем секреции ИЛ 10 и трансформирующего фактора роста (? )
IV_lektsia_Adaptivny_immunitet.pptx