

 внутрь") • 1. . В основе процесса деполяризации лежит движение ионов: • а) внутрь клетки Na+ • б) внутрь клетки K+ • в) внутрь клетки Cl • г) из клетки Na+ • д) из клетки K+ • е) из клетки Cl-
• 1. . В основе процесса деполяризации лежит движение ионов: • а) внутрь клетки Na+ • б) внутрь клетки K+ • в) внутрь клетки Cl • г) из клетки Na+ • д) из клетки K+ • е) из клетки Cl-
 работы K+/ Na+ насоса •") • Потенциал действия возникает за счет: • а) работы K+/ Na+ насоса • б) движения ионов K+ • в) движения ионов Na+
• Потенциал действия возникает за счет: • а) работы K+/ Na+ насоса • б) движения ионов K+ • в) движения ионов Na+
 деполяризации • б) реполяризации •") • Относительная рефрактерность наблюдается во время: • а) деполяризации • б) реполяризации • в) гиперполяризации
• Относительная рефрактерность наблюдается во время: • а) деполяризации • б) реполяризации • в) гиперполяризации
 увеличение амплитуды локального ответа •") • При увеличении силы стимула происходит: • а) увеличение амплитуды локального ответа • б) увеличение амплитуды ПД • в) увеличение амплитуды и локального ответа, и ПД • д) увеличение длительности ПД • е) увеличение длительности и локального ответа, и ПД
• При увеличении силы стимула происходит: • а) увеличение амплитуды локального ответа • б) увеличение амплитуды ПД • в) увеличение амплитуды и локального ответа, и ПД • д) увеличение длительности ПД • е) увеличение длительности и локального ответа, и ПД
 уменьшается во время локального ответа • б)") • Возбудимость мембраны нейрона: • а) уменьшается во время локального ответа • б) уменьшается во время ПД • в) уменьшается во время и локального ответа, и ПД
• Возбудимость мембраны нейрона: • а) уменьшается во время локального ответа • б) уменьшается во время ПД • в) уменьшается во время и локального ответа, и ПД
 деполяризации •") • В тормозных синапсах на постсинаптической мембране возникает процесс: • а) деполяризации • б) реполяризации • в) гиперполяризации • г) поляризации
• В тормозных синапсах на постсинаптической мембране возникает процесс: • а) деполяризации • б) реполяризации • в) гиперполяризации • г) поляризации
 постсинаптической части • б)") • В состоянии покоя медиатор находиться в • а) постсинаптической части • б) пресинаптической части • в) межсинаптической щели • г) в везикулах
• В состоянии покоя медиатор находиться в • а) постсинаптической части • б) пресинаптической части • в) межсинаптической щели • г) в везикулах
 деполяризации •") • В возбуждающих синапсах на постсинаптической мембране возникает процесс • а) деполяризации • б) реполяризации • в) гиперполяризации • г) поляризации
• В возбуждающих синапсах на постсинаптической мембране возникает процесс • а) деполяризации • б) реполяризации • в) гиперполяризации • г) поляризации
 омического сопротивления") • Скорость проведения ПД по нервному волокну зависит от: • а) омического сопротивления аксона • б) сопротивления внеклеточной среды • в) диаметра волокна • г) наличия миелина
• Скорость проведения ПД по нервному волокну зависит от: • а) омического сопротивления аксона • б) сопротивления внеклеточной среды • в) диаметра волокна • г) наличия миелина
 Ствол мозга Часть головного мозга, включающая в себя v продолговатый мозг, v Варолиев мост, v средний мозг,
Ствол мозга Часть головного мозга, включающая в себя v продолговатый мозг, v Варолиев мост, v средний мозг,
 Медиальная поверхность
Медиальная поверхность
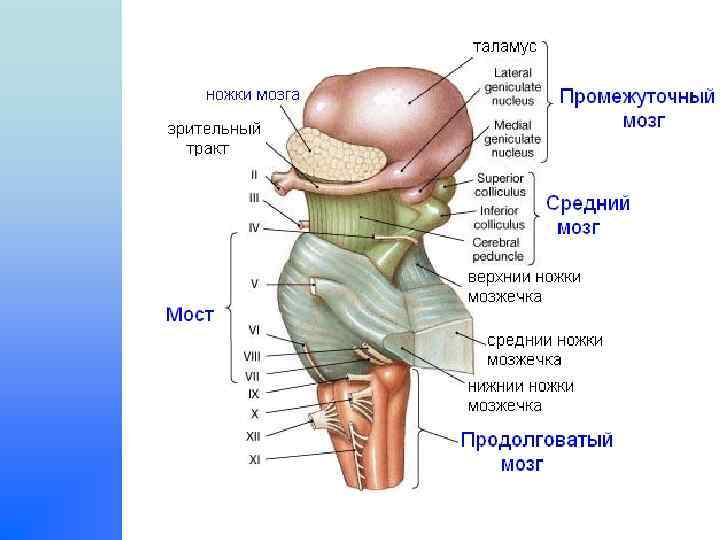


 Дорзальная поверхность
Дорзальная поверхность
 Медиальный срез
Медиальный срез
 Средний мозг v. Рефлекторная функция v. Проводящая функция ШНо не только
Средний мозг v. Рефлекторная функция v. Проводящая функция ШНо не только
 • Эволюция среднего мозга связана с развитием зрения.
• Эволюция среднего мозга связана с развитием зрения.
 Рефлексы
Рефлексы
 Ядро") Зрачковый рефлекс § Парасимпатические влияния –кольцевые мышцы – сужение ( близко расположенные предметы) Ядро Якубовича § Симпатические – радиальные мышцы- расширение ( при асфиксии)
Зрачковый рефлекс § Парасимпатические влияния –кольцевые мышцы – сужение ( близко расположенные предметы) Ядро Якубовича § Симпатические – радиальные мышцы- расширение ( при асфиксии)
 • Ближайшая точка ясного видения - 10 см
• Ближайшая точка ясного видения - 10 см
") Аккомодация • изменение кривизны хрусталика (близко – выпуклый)
Аккомодация • изменение кривизны хрусталика (близко – выпуклый)
 Саккады • быстрые, строго согласованные движения глаз, происходящие одновременно и в одном направлении
Саккады • быстрые, строго согласованные движения глаз, происходящие одновременно и в одном направлении
 Характеристика саккад • • Интервалы Амплитуда Ориентация Длительность – 10 - 80 мс
Характеристика саккад • • Интервалы Амплитуда Ориентация Длительность – 10 - 80 мс





 Организация движения
Организация движения
 Черная субстанция • очень древняя, жевание, глотание, пластический тонус, древняя, • координирует акты жевания и глотания, участвует в регуляции пластического тонуса, а у человека — и в мелких движениях пальцев рук. • Патология - паркинсон - обеднение содружественных движений – лицо маска
Черная субстанция • очень древняя, жевание, глотание, пластический тонус, древняя, • координирует акты жевания и глотания, участвует в регуляции пластического тонуса, а у человека — и в мелких движениях пальцев рук. • Патология - паркинсон - обеднение содружественных движений – лицо маска

 • Гибель дофаминергических нейронов черной субстанции приводит к снижению уровня дофамина в стриатуме и растормаживанию вставочных холинергических нейронов полосатого тела.
• Гибель дофаминергических нейронов черной субстанции приводит к снижению уровня дофамина в стриатуме и растормаживанию вставочных холинергических нейронов полосатого тела.
 • Тремор покоя, • мышечная ригидность • гипокинезия
• Тремор покоя, • мышечная ригидность • гипокинезия
 • Нейрогенез • Снижение дофамина и серотонина снижает нейрогенез
• Нейрогенез • Снижение дофамина и серотонина снижает нейрогенез
 Красное ядро • У рептилий • Участие в перераспределении мышечного тонуса и осуществлении и координировании тонических рефлексов. • Напр. Выпрямительных (участвуют рецепторы лабиринтов, шейных мышц, и кожной поверхности тела • Тонические рефлексы
Красное ядро • У рептилий • Участие в перераспределении мышечного тонуса и осуществлении и координировании тонических рефлексов. • Напр. Выпрямительных (участвуют рецепторы лабиринтов, шейных мышц, и кожной поверхности тела • Тонические рефлексы
 Децеребрационная ригидность • Рассечение ствола мозга у четвероногих каудальнее красного ядра • красное ядро осуществляет тормозное влияние на мышцы разгибатели
Децеребрационная ригидность • Рассечение ствола мозга у четвероногих каудальнее красного ядра • красное ядро осуществляет тормозное влияние на мышцы разгибатели



 • Двусторонее разрушение голубого пятна приводит к полному исчезновению сна с быстрыми движениями глаз
• Двусторонее разрушение голубого пятна приводит к полному исчезновению сна с быстрыми движениями глаз
 Мезенцефалическое сенсорное ядро тройничного нерва.
Мезенцефалическое сенсорное ядро тройничного нерва.
 ЦСВ • Восприятие боли • Вокализации
ЦСВ • Восприятие боли • Вокализации
 ЦСВ • У самок млекопитающих регулируют реакцию лордоза
ЦСВ • У самок млекопитающих регулируют реакцию лордоза
 • однако у материнской любви есть и свой специфический подкорковый участок, который не имеет отношения к страстной любви, — околоводопроводное серое вещество. Этот участок тесно связан с эмоциональными центрами лимбической системы и содержит много вазопрессиновых рецепторов, которые очень важны для формирования материнской привязанности. Прослеживается и еще одна связь между ОСВ и материнской любовью: этот участок мозга имеет отношение к подавлению чувства боли при сильных эмоциональных переживаниях, в том числе во время родов.
• однако у материнской любви есть и свой специфический подкорковый участок, который не имеет отношения к страстной любви, — околоводопроводное серое вещество. Этот участок тесно связан с эмоциональными центрами лимбической системы и содержит много вазопрессиновых рецепторов, которые очень важны для формирования материнской привязанности. Прослеживается и еще одна связь между ОСВ и материнской любовью: этот участок мозга имеет отношение к подавлению чувства боли при сильных эмоциональных переживаниях, в том числе во время родов.
 Сенсорные системы v. Слуховая v. Зрительная v. Соматическая
Сенсорные системы v. Слуховая v. Зрительная v. Соматическая
 Ретикулярная формация v. Восходящие влияния v. Нисходящие влияния
Ретикулярная формация v. Восходящие влияния v. Нисходящие влияния
 Ретикулярная формация v облегчающие восходящие влияния на кору v участие в организации сна v уменьшение частоты разряда во время сна перерезка – медленные волны в коре стимуляция - реакция пробуждения v на спинной мозг и активирующее и тормозное влияние
Ретикулярная формация v облегчающие восходящие влияния на кору v участие в организации сна v уменьшение частоты разряда во время сна перерезка – медленные волны в коре стимуляция - реакция пробуждения v на спинной мозг и активирующее и тормозное влияние
 • Возбудительный тонус в ретикулярных нейронах может поддерживаться благодаря циркуляции импульсов по нейронным замкнутым кольцевым цепям • Это определяет готовность к деятельности этих отделов центральной нервной системы, обеспечивая их быстрое реагирование и включение реакции настораживания.
• Возбудительный тонус в ретикулярных нейронах может поддерживаться благодаря циркуляции импульсов по нейронным замкнутым кольцевым цепям • Это определяет готовность к деятельности этих отделов центральной нервной системы, обеспечивая их быстрое реагирование и включение реакции настораживания.
 Ориентировочный рефлекс ЧТО ТАКОЕ?
Ориентировочный рефлекс ЧТО ТАКОЕ?
 Ориентировочный рефлекс ЧТО ТАКОЕ?
Ориентировочный рефлекс ЧТО ТАКОЕ?
 Мезолимбическая система
Мезолимбическая система
 Мезолимбическая система • Источники дофаминергических проекций, т. е. тела нейронов этой системы, расположены в вентральном поле покрышки среднего мозга и частично в компактной части черной субстанции. • Их отростки идут в • поясную извилину, • энториальную кору, • миндалину, • обонятельный бугорок, • аккумбентное ядро, • гиппокамп, • парагиппокампальную извилину, • перегородку и. • участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
Мезолимбическая система • Источники дофаминергических проекций, т. е. тела нейронов этой системы, расположены в вентральном поле покрышки среднего мозга и частично в компактной части черной субстанции. • Их отростки идут в • поясную извилину, • энториальную кору, • миндалину, • обонятельный бугорок, • аккумбентное ядро, • гиппокамп, • парагиппокампальную извилину, • перегородку и. • участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
 Мезолимбическая система • участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
Мезолимбическая система • участвует в механизмах памяти, эмоций, обучения и нейроэндокринной регуляции.
 Мезолимбическая система • Чувства радости и удовольствия зависят от активности дофаминовых нейронов вентральной области покрышки среднего мозга (VTA)
Мезолимбическая система • Чувства радости и удовольствия зависят от активности дофаминовых нейронов вентральной области покрышки среднего мозга (VTA)
 — ключевой компонент") Вентральная область покрышки • Вентральная область покрышки (ventral tegmental area, VTA) — ключевой компонент «системы вознаграждения» (также известной как «система внутреннего подкрепления» , reward system) в мозге млекопитающих. • Примерно 55– 65% нейронов VTA — это дофаминовые (дофаминэргические) нейроны, Дофамин играет в мозге роль «вещества удовольствия» ? ) Дофаминовые сигналы из VTA поступают 1. в префронтальную кору, 2. в гиппокамп 3. в прилежащее ядро (nucleus accumbens), которое можно назвать «главным центром удовольствия» и которое отвечает за мотивацию, привязанности и зависимости, 4. в миндалину (amygdala) • • •
Вентральная область покрышки • Вентральная область покрышки (ventral tegmental area, VTA) — ключевой компонент «системы вознаграждения» (также известной как «система внутреннего подкрепления» , reward system) в мозге млекопитающих. • Примерно 55– 65% нейронов VTA — это дофаминовые (дофаминэргические) нейроны, Дофамин играет в мозге роль «вещества удовольствия» ? ) Дофаминовые сигналы из VTA поступают 1. в префронтальную кору, 2. в гиппокамп 3. в прилежащее ядро (nucleus accumbens), которое можно назвать «главным центром удовольствия» и которое отвечает за мотивацию, привязанности и зависимости, 4. в миндалину (amygdala) • • •
 и наших") • Их активность, в свою очередь, определяется соотношением награды (положительного стимула) и наших ожиданий • П. В. Симонов
• Их активность, в свою очередь, определяется соотношением награды (положительного стимула) и наших ожиданий • П. В. Симонов
 Часть нейронов VTA специализируется на кодировании ожиданий, не реагируя на саму награду. Эти нейроны — тормозные (ГАМК‑эргические). Их регулярное возбуждение снижает активность дофаминовых нейронов VTA и тем самым притупляет чувство радости от «хороших новостей» , которые были известны животному заранее. (оптогенетика)
Часть нейронов VTA специализируется на кодировании ожиданий, не реагируя на саму награду. Эти нейроны — тормозные (ГАМК‑эргические). Их регулярное возбуждение снижает активность дофаминовых нейронов VTA и тем самым притупляет чувство радости от «хороших новостей» , которые были известны животному заранее. (оптогенетика)
 • 1. Дофаминовые нейроны VTA активируются в ответ на положительные стимулы (информацию о которых они получают от других отделов мозга, в том числе от коры), а также на условные стимулы, предвещающие награду. • 2. Например, если животное знает, что после звукового сигнала ему дают что-нибудь вкусненькое, дофаминовые нейроны VTA будут возбуждаться в ответ на звуковой сигнал. 3. Сама же награда ( «безусловный стимул» ) активирует их в зависимости от своей предсказуемости: неожиданная награда вызывает сильную активацию, но если животное точно знает, что угощение всегда появляется после звонка, то нейроны энергично ответят только на звонок, а на саму награду отреагируют слабее или не отреагируют вовсе. 4. Ну а если после звонка положенная награда не появится, активность дофаминовых нейронов VTA отобразит разочарование, опустившись ниже базового ( «спокойного» ) уровня. • • • Таким образом, работа этих нейронов кодирует не хорошие новости как таковые, а, скорее, степень их соответствия ожиданиям, «ошибку предсказания награды» (reward prediction error, RPE)
• 1. Дофаминовые нейроны VTA активируются в ответ на положительные стимулы (информацию о которых они получают от других отделов мозга, в том числе от коры), а также на условные стимулы, предвещающие награду. • 2. Например, если животное знает, что после звукового сигнала ему дают что-нибудь вкусненькое, дофаминовые нейроны VTA будут возбуждаться в ответ на звуковой сигнал. 3. Сама же награда ( «безусловный стимул» ) активирует их в зависимости от своей предсказуемости: неожиданная награда вызывает сильную активацию, но если животное точно знает, что угощение всегда появляется после звонка, то нейроны энергично ответят только на звонок, а на саму награду отреагируют слабее или не отреагируют вовсе. 4. Ну а если после звонка положенная награда не появится, активность дофаминовых нейронов VTA отобразит разочарование, опустившись ниже базового ( «спокойного» ) уровня. • • • Таким образом, работа этих нейронов кодирует не хорошие новости как таковые, а, скорее, степень их соответствия ожиданиям, «ошибку предсказания награды» (reward prediction error, RPE)
, предвещающий награду, но не на саму награду (капля") Нейрон реагирует на условный стимул (CS), предвещающий награду, но не на саму награду (капля воды). Хотя он получает два возбуждающих входных сигнала (об условном стимуле и о награде, черные линии) Он также получает тормозящий сигнал от неких нейронов, активность которых кодирует ожидание награды (красная линия)
Нейрон реагирует на условный стимул (CS), предвещающий награду, но не на саму награду (капля воды). Хотя он получает два возбуждающих входных сигнала (об условном стимуле и о награде, черные линии) Он также получает тормозящий сигнал от неких нейронов, активность которых кодирует ожидание награды (красная линия)
 показал, что") Анализ активности нейронов в четырех ситуациях (большая награда, малая награда, ничего, наказание) показал, что нейроны VTA четко подразделяются на три типа. • Нейроны типа I — самые многочисленные. Они активируются в ответ на условный стимул, предвещающий награду. Затем их активность быстро возвращается к базовому уровню и остается низкой всё время, пока мышь ждет награду. На саму награду, появление которой было «предсказано» условным стимулом, реагирует только часть нейронов первого типа. При этом реакция на ожидаемую награду снижается день ото дня. Иными словами, по мере того, как мышь всё более привыкает к тому, что после данного условного стимула обязательно появляется награда, реакция на нее у нейронов первого типа уменьшается (хотя реакция на условный стимул остается сильной). • Таким образом, нейроны типа I ведут себя в точности так, как положено вести себя дофаминовым нейронам VTA:
Анализ активности нейронов в четырех ситуациях (большая награда, малая награда, ничего, наказание) показал, что нейроны VTA четко подразделяются на три типа. • Нейроны типа I — самые многочисленные. Они активируются в ответ на условный стимул, предвещающий награду. Затем их активность быстро возвращается к базовому уровню и остается низкой всё время, пока мышь ждет награду. На саму награду, появление которой было «предсказано» условным стимулом, реагирует только часть нейронов первого типа. При этом реакция на ожидаемую награду снижается день ото дня. Иными словами, по мере того, как мышь всё более привыкает к тому, что после данного условного стимула обязательно появляется награда, реакция на нее у нейронов первого типа уменьшается (хотя реакция на условный стимул остается сильной). • Таким образом, нейроны типа I ведут себя в точности так, как положено вести себя дофаминовым нейронам VTA:
 • Нейроны типа II тоже увеличивают свою активность в ответ на условный стимул, предвещающий награду, • Но! в отличие от нейронов типа I, они продолжают активно работать в течение всего периода ожидания. При этом их активность коррелирует с размером ожидаемой награды. Реакция на саму награду выражена слабо. • • На отсутствие награды после положительного условного стимула ( «разочарование» ) эти нейроны не реагируют. • Это оказались ГАМК нейроны
• Нейроны типа II тоже увеличивают свою активность в ответ на условный стимул, предвещающий награду, • Но! в отличие от нейронов типа I, они продолжают активно работать в течение всего периода ожидания. При этом их активность коррелирует с размером ожидаемой награды. Реакция на саму награду выражена слабо. • • На отсутствие награды после положительного условного стимула ( «разочарование» ) эти нейроны не реагируют. • Это оказались ГАМК нейроны
 • Нейроны типа III уменьшают свою активность в ответ на «хорошие» условные стимулы и немного увеличивают в ответ на «плохие» . • природа и функции этих нейронов остались неизвестными (((.
• Нейроны типа III уменьшают свою активность в ответ на «хорошие» условные стимулы и немного увеличивают в ответ на «плохие» . • природа и функции этих нейронов остались неизвестными (((.
 и негативных") VTA получает возбуждающие и тормозящие входные сигналы о позитивных ожиданиях (Expectation) и негативных стимулах (Aversion) из префронтальной коры , латеральной уздечки , педункулопонтийного ядра , стриатума. Сигналы дофаминовых нейронов VTA, несущие информацию об ошибке предсказания награды (RPE), поступают в кору (Сtx), стриатум, миндалину (Amy), гиппокамп (Hipp)
VTA получает возбуждающие и тормозящие входные сигналы о позитивных ожиданиях (Expectation) и негативных стимулах (Aversion) из префронтальной коры , латеральной уздечки , педункулопонтийного ядра , стриатума. Сигналы дофаминовых нейронов VTA, несущие информацию об ошибке предсказания награды (RPE), поступают в кору (Сtx), стриатум, миндалину (Amy), гиппокамп (Hipp)
 • На самом деле все сложнее • Jeremiah Y. Cohen, Sebastian Haesler, Linh Vong, Bradford B. Lowell, Naoshige Uchida. Neuron-type-specific signals for reward and punishment in the ventral tegmental area // Nature. 2012. V. 482. P. 85– 88.
• На самом деле все сложнее • Jeremiah Y. Cohen, Sebastian Haesler, Linh Vong, Bradford B. Lowell, Naoshige Uchida. Neuron-type-specific signals for reward and punishment in the ventral tegmental area // Nature. 2012. V. 482. P. 85– 88.

