f5facbaaca344e3f8b99c21064208e16.ppt
- Количество слайдов: 42


 έχουν καταλυτική")
ΜΙΚΡΟΒΙΑΚΗ ΕΞΕΛΙΞΗ ΚΑΙ ΣΥΣΤΗΜΑΤΙΚΗ Λαμβάνοντας υπ’οψιν ότι ορισμένα είδη ριβονουκλεϊκού οξέος (RNA) έχουν καταλυτική δράση, πιστεύεται ότι οι πρώτες μορφές ζωής δεν διέθεταν καθόλου DNA, περιείχαν ελάχιστες έως καθόλου πρωτεΐνες, και αποτελούνταν κυρίως από RNA. Το αυτοαντιγραφόμενο RNA θα μπορούσε να εξελιχθεί σε αυτόνομες κυτταρικές οντότητες με την ενσωμάτωσή του σε κυστίδια λιποπρωτεϊνών. Με την πάροδο του χρόνου, οι πρωτεΐνες αντικατέστησαν τις καταλυτικές λειτουργίες του RNA και το DNA αντικατέστησε τις κωδικοποιητικές λειτουργίες του RNA. Αυτές οι μορφές ζωής με βάση το RNA είναι δυνατόν να εξελίχθηκαν στην πρώτη κυτταρική μορφή ζωής, όταν τα αυτοαντιγραφόμενα RNA βρέθηκαν εγκλωβισμένα σε λιποπρωτεϊνικά κυστίδια. Αυτό μπορεί να συνέβη αναρίθμητες φορές στην πρώιμη Γη χωρίς να ευοδωθεί, αλλά κάποια στιγμή βρέθηκαν μαζί τα κατάλληλα συστατικά που επέτρεψαν την εμφάνιση του πρωτόγονου κυτταρικού οργανισμού. Είναι πιθανόν οι πρωτεΐνες να εμφανίστηκαν αρχικά ως σύμπλοκα με το RNA, και καθώς η εξέλιξη επέλεγε ολοένα και ακριβέστερους βιοχημικούς καταλύτες, το RNA τελικά αντικαταστάθηκε από τις πρωτεΐνες στον ρόλο του κυτταρικού ενζύμου. Η καθιέρωση του DNA ως γονιδιώματος του κυττάρου προέκυψε από την εξελικτική πίεση προς την κατεύθυνση της καλλίτερης απόδοσης και πιστότητας στην αντιγραφή της γενετικής πληροφορίας (οι DNA πολυμεράσες είναι πολύ πιο ακριβείς από τις RNA πολυμεράσες). Κάποια στιγμή στα πρώτα στάδια της μικροβιακής εξέλιξης, το τριμερές σύστημα - DNA, RNA και πρωτεΐνες - παγιώθηκε στις κυτταρικές μορφές ζωής ως η βέλτιστη λύση για τη διαχείριση της βιολογικής πληροφορίας

Μεταβολισμός στους πρωτόγονους οργανισμούς Μέχρι την ανάπτυξη των κυανοβακτηρίων, το μοριακό οξυγόνο δεν υπήρχε σε αξιοσημείωτες ποσότητες στη Γη. Οι πρώιμοι οργανισμοί θα μπορούσαν να εκμεταλλευτούν μόνον εκείνους τους ενεργειακούς μηχανισμούς που μπορούν να δράσουν σε ανοξικές συνθήκες. Μια απλή χημική αντίδραση που χρησιμοποιεί δισθενή σίδηρο (ο οποίος αφθονούσε στην πρώιμη Γη) ίσως υπήρξε η μέθοδος την οποία χρησιμοποίησαν οι πρωτόγονοι οργανισμοί για να εξασφαλίσουν ενέργεια. Η αντίδραση 2 Fe. S + H 2 S Fe 2 S 3 + H 2 ΔG 0΄ = - 42 k. J/αντίδραση συντελείται εξώεργα, απελευθερώνοντας ενέργεια. Η αντίδραση παράγει επίσης H 2, και επομένως τα πρωτόγονα κύτταρα χρησιμοποίησαν αυτό το H 2 για να δημιουργήσουν μια πρωτονιεργετική δύναμη δια μέσου της μεμβράνης, από την οποία μια πρωτόγονη ΑΤΡάση θα μπορούσε να ανακτήσει χρήσιμη χημική ενέργεια υπό τη μορφή ΑΤΡ. Με το H 2 ως δότη ηλεκτρονίων, θα χρειαζόταν και ένας δέκτης ηλεκτρονίων, που θα μπορούσε να είναι το στοιχειακό θείο. Πολλά υπερθερμόφιλα Αρχαία ( οι πλησιέστεροι συγγενείς των πρωιμότερων οργανισμών της Γης που υπάρχουν σήμερα) μπορούν να εκτελέσουν την αντίδραση αυτή. Οι πρωτόγονοι οργανισμοί μπορεί να εξασφάλιζαν άνθρακα από ποικίλες πηγές, ακόμη και από CO 2, ένα αέριο που υπήρχε σε αφθονία στην πρώιμη Γη. Η αυτοτροφία αποτελεί χαρακτηριστικό αρκετών υπερθερμόφιλων με μικρό γονιδίωμα, που διαχωρίζονται κοντά στη ρίζα στο εξελικτικό δένδρο της ζωής

Οξυγόνωση της ατμόσφαιρας Κυανοβακτήρια: οι οργανισμοί αυτοί εμφανίστηκαν για πρώτη φορά στη Γη πιθανόν πριν από 3 δισεκατομμύρια χρόνια περίπου, αλλά το Ο 2 που παρήγαγαν δεν συσσωρεύθηκε στην ατμόσφαιρα. Αυτό συνέβη επειδή όλες οι αναγωγικές ενώσεις (όπως το Fe. S), που υπήρχαν τότε αντιδρούσαν αυθόρμητα με το Ο 2. Είναι πολύ πιθανό ότι τα κυανοβακτήρια εξελίχθηκαν μέσα από την ανάπτυξη ενός φωτοσυστήματος που θα μπορούσε να χρησιμοποιήσει Η 2Ο ως ηλεκτρονιοδότη για τη φωτοσυνθετική αναγωγή του CO 2, ελευθερώνοντας έτσι Ο 2 ως ένα δεύτερο προϊόν (CO 2 + Η 2Ο CH 2Ο + Ο 2). Η εξέλιξη της φωτοσύνθεσης που παράγει οξυγόνο είχε τεράστιες συνέπειες για το γήινο περιβάλλον, διότι με τη συσσώρευση του Ο 2, η ατμόσφαιρα βαθμιαία άλλαξε από ανοξική σε οξυγονούχο. Με το Ο 2 να είναι διαθέσιμο ως δέκτης ηλεκτρονίων, μπορούσαν πλέον να εξελιχθούν οι αερόβιοι οργανισμοί. Την εποχή περίπου η ατμόσφαιρα της Γης έγινε οξειδωτική σημειώθηκαν ταχύτατες εξελίξεις που οδήγησαν στην εμφάνιση ευκαρυωτικών οργανισμών με εσωτερικά οργανίδια και στην ταχεία διαφοροποίηση των πολυκύτταρων οργανισμών και εν τέλει στα ανώτερα ζώα και φυτά. Μια άλλη σημαντική συνέπεια της εμφάνισης του Ο 2 ήταν ο σχηματισμός του όζοντος (Ο 3) που δρα προστατευτικά κατά της έντονης υπεριώδους ακτινοβολίας που φθάνει στη Γη από τον ήλιο. Μέχρι την ανάπτυξη μιας ασπίδας όζοντος, η εξέλιξη μπορούσε να λειτουργεί μόνο σε περιβάλλοντα προστατευμένα από την άμεση ηλιακή ακτινοβολία, επειδή η έντονη υπεριώδης ακτινοβολία θα προκαλούσε θανάσιμες καταστροφές στο DNA.

Η ανάπτυξη του ευκαρυωτικού πυρήνα διευκόλυνε επίσης τη διαχείριση του τεράστιου γονιδιώματος και επέτρεψε επίσης τον ανασυνδυασμό των γονιδιωμάτων. Ενδοσυμβίωση Η ενδοσυμβιωτική θεωρία, υποστηρίζει ότι ένα αερόβιο βακτήριο παγίωσε τη θέση του στο κυτταρόπλασμα ενός πρωτόγονου ευκαρυώτη και του προμήθευε ενέργεια, διασφαλίζοντας ως αντάλλαγμα ένα προστατευμένο περιβάλλον και την απρόσκοπτη παροχή θρεπτικών υλικών. Αυτός ο συμβιώτης ήταν ο πρόδρομος του σημερινού μιτοχονδρίου. Παρομοίως, η ενδοσυμβιωτική ενσωμάτωση ενός φωτότροφου παραγωγού οξυγόνου θα είχε προσδώσει φωτοσυνθετικές ικανότητες σε έναν πρωτόγονο ευκαρυώτη, ούτως ώστε να πάψει να εξαρτάται από οργανικές ενώσεις για την παραγωγή ενέργειας. Αυτός ο φωτότροφος συμβιώτης ήταν ο πρόδρομος του σημερινού χλωροπλάστη. Τόσο οι χλωροπλάστες όσο και τα μιτοχόνδρια περιέχουν ριβοσώματα και μάλιστα προκαρυωτικού τύπου. Τα ριβοσώματα από τα οργανίδια αυτά παρουσιάζουν αλληλουχίες RNA που χαρακτηρίζουν συγκεκριμένα Βακτήρια, και η λειτουργία τους παρεμποδίζεται από τα ίδια αντιβιοτικά που επιδρούν στα ριβοσώματα αυτόνομων Βακτηρίων. Τα μιτοχόνδρια και οι χλωροπλάστες περιέχουν επίσης μικρές ποσότητες κυκλικού DNA, χαρακτηριστικού των προκαρυωτών.

Τα ριβοσωματικά RNA ως εξελικτικά χρονόμετρα Τα ριβοσωματικά RNA έχουν σταθερή λειτουργικότητα, είναι οικουμενικώς κατανεμημένα και έχουν μια μετρίως διατηρημένη αλληλουχία σε ένα ευρύ φάσμα φυλογενετικών αποστάσεων. Σε μεγάλα μόρια όπως το ριβοσωματικό RNA, η ομοιότητα μεταξύ δύο αλληλουχιών αποτελεί πάντοτε ένδειξη κάποιας φυλογενετικής σχέσης. Εκείνο που συνιστά ένδειξη για τη σχετική εξελικτική απόσταση είναι ο βαθμός της ομοιότητας των αλληλουχιών των ριβοσωματικών RNA μεταξύ δύο οργανισμών. Από συγκριτικές αναλύσεις μπορούν να κατασκευαστούν οι μοριακές γενεαλογίες οι οποίες οδηγούν στα φυλογενετικά δέντρα που δείχνουν τις πιθανότερες εξελικτικές θέσεις των οργανισμών, όταν τοποθετούνται ο ένας σε σχέση με τον άλλο. Υπάρχουν τρία μόρια ριβοσωματικού RNA, τα οποία στους προκαρυώτες έχουν μέγεθος 5 S, 16 S και 23 S. Τα μεγάλα βακτηριακά r. RNA, τα 16 S και 23 S (με περίπου 1500 και 2900 νουκλεοτίδια, αντιστοίχως), περιέχουν αρκετές περιοχές με συντηρημένες αλληλουχίες σε υψηλό βαθμό, χρήσιμες για την επίτευξη σωστών στοιχίσεων, αλλά χαρακτηρίζονται και από σημαντική μεταβλητότητα σε άλλες περιοχές του μορίου, ώστε να χρησιμεύουν ως εξαιρετικά φυλογενετικά χρονόμετρα. Η βάση δεδομένων των αλληλουχιών r. RNA (Ribosomal Database Project – RDP) είναι προσβάσιμη μέσω Διαδικτύου. Οι νεοπροσδιοριζόμενες αλληλουχίες συγκρίνονται με τις υπάρχουσες στο RDP και με αλληλουχίες από άλλες βάσεις, όπως η Gen. Bank (ΗΠΑ) ή αυτή του EMBL (Γερμανία).
 για")
Για την ανάλυση του ριβοσωματικού RNA χρησιμοποιείται η αλυσιδωτή αντίδραση της πολυμεράσης (PCR) για την ενίσχυση των γονιδίων που κωδικοποιούν το ριβοσωματικό RNA 16 S. Στη συνέχεια προσδιορίζεται η αλληλουχία του προϊόντος της PCR. Μόλις ολοκληρωθεί ο προσδιορισμός της αλληλουχίας, τα δεδομένα είναι έτοιμα για επεξεργασία με τη βοήθεια κατάλληλων λογισμικών (υπάρχουν αρκετοί διαφορετικοί αλγόριθμοι ανάλυσης αλληλουχιών και δημιουργίας φυλογενετικών δέντρων διαθέσιμοι για τη συγκριτική εξέταση του ριβοσωματικού RNA ). Ωστόσο, ανεξαρτήτως από το ποιο πρόγραμμα χρησιμοποιείται, οι αλληλουχίες πριν από την επεξεργασία πρέπει να «στοιχηθούν» . Οι στοιχισμένες αλληλουχίες εισάγονται κατόπιν στο κατάλληλο λογισμικό και γίνονται οι συγκριτικές αναλύσεις. Με την εφαρμογή μεθόδων «απόστασης» , οι αλληλουχίες αντιστοιχίζονται και ο υπολογιστής μετρά κάθε δυνατή θέση στα δεδομένα στην οποία υπάρχει διαφορά, για να υπολογίσει μια εξελικτική απόσταση (ΕD). Η εξελικτική απόσταση υπολογίζεται ως το ποσοστό των μη πανομοιότυπων αλληλουχιών μεταξύ των r. RNA δυο οργανισμών. Η διορθωμένη ΕD προκύπτει από την στατιστική διόρθωση αναγκαία για τον συνυπολογισμό ανάστροφων μεταλλάξεων που επανέφεραν τον αρχικό γονότυπο. Από τα δεδομένα αυτά, μπορεί να κατασκευαστεί ένας πίνακας που δείχνει την ΕD μεταξύ οιουδήποτε ζεύγους αλληλουχιών στο σύνολο των δεδομένων. Τέλος, κατασκευάζεται ένα φυλογενετικό δένδρο στο οποίο οι αποστάσεις των γραμμών είναι ανάλογες των εξελικτικών αποστάσεων.

Κατασκευή Δενδρογράμματος βάσει του % ομοιότητας μεταξύ 5 στοιχισμένων αλληλουχιών.

Με το ίδιο σκεπτικό μπορεί να κατασκευασθεί δενδρόγραμμα βάσει του συντελεστή «ανομοιότητας» ή εξελικτικής απόστασης
 Διορθωμένες")
Η εξελικτική απόσταση μπορεί να διορθωθεί βάσει του κατωτέρω τύπου (dissimilarity= εξελικτική απόσταση) Διορθωμένες τιμές εξελικτικής απόστασης

Φυλογενετική Ανάλυση χρησιμοποιώντας τον αλγόριθμο UPGMA

Φυλογενετική Ανάλυση χρησιμοποιώντας την μέθοδο “neighbor-joining” To ολικό μήκος των κλάδων που χωρίζουν δυο μικροοργανισμούς είναι ανάλογο προς την ΕD. Χρησιμοποιείται επίσης μια στατιστική διεργασία το «boot strapping» με την οποία δημιουργούνται πολλές εκδοχές του δένδρου ώστε να επιβεβαιωθεί ότι το τελικό δένδρο παρουσιάζει τη βέλτιστη συμφωνία των δεδομένων.

Αλληλουχίες – υπογραφή Πρόκειται για μικρές αλληλουχίες νουκλεοτιδίων που απαντούν αποκλειστικά σε ορισμένες ομάδες μικροοργανισμών. Π. χ. οι αλληλουχίες υπογραφές μας βοηθούν να τοποθετήσουμε στη σωστή φυλογενετική ομάδα οργανισμούς που απομονώθηκαν για πρώτη φορά. Ανιχνευτές κατάλληλης εξειδίκευσης μπορούν να σχεδιαστούν ούτως ώστε να υβριδοποιούνται μόνο με το RNA οργανισμών από τον «χώρο» των Βακτηρίων. Παρομοίως, είναι εφικτό να σχεδιαστούν ανιχνευτές με εξειδίκευση για είδη Αρχαίων ή Ευκαρύων.

Η υβριδοποίηση των ανιχνευτών σε κυτταρικά ριβοσώματα μπορεί να παρατηρηθεί στο μικροσκόπιο, εάν προσδεθεί στον ανιχνευτή μια φθορίζουσα χρωστική. Μετά τον υβριδισμό του ανιχνευτή με το ριβοσωματικό RNA, τα κύτταρα φθορίζουν και μπορούν να εξεταστούν σε μικροσκόπιο φθορισμού. Η τεχνική αυτή είναι γνωστή ως in situ φθορίζων υβριδισμός (FISH – fluorescent In Situ Hybridization).
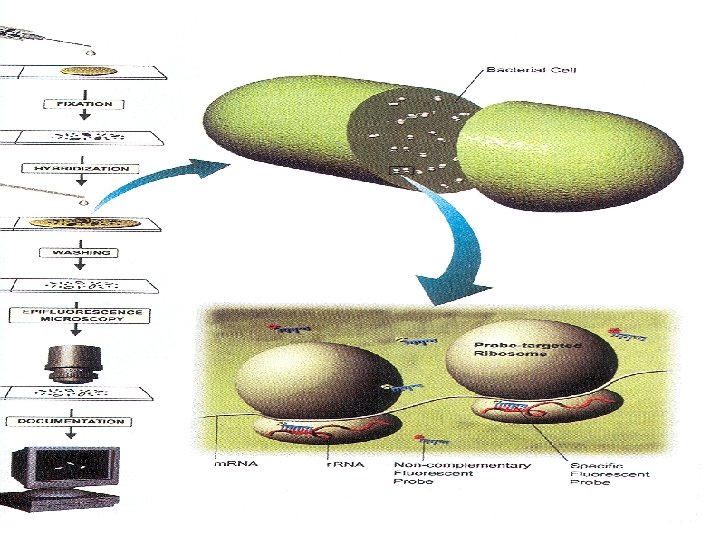

Η τεχνολογία FISH χρησιμοποιείται ευρέως στη μικροβιακή οικολογία και στην κλινική διαγνωστική. Στην οικολογία μπορεί να εφαρμοστεί στη μικροσκοπική ταυτοποίηση και την παρακολούθηση μικροοργανισμών απευθείας στο περιβάλλον. Στην κλινική διαγνωστική, η FISH έχει χρησιμεύσει στην ταχεία ταυτοποίηση συγκεκριμένων παθογόνων σε δείγματα από ασθενείς. Με την τεχνική αυτή παρακάμπτουμε την ανάγκη να καλλιεργήσουμε έναν οργανισμό στο εργαστήριο, γεγονός που επιτρέπει να αρχίσουμε μια θεραπευτική αγωγή ώρες ή ημέρες νωρίτερα απ’ ότι αν χρησιμοποιούσαμε τις κλασικές τεχνικές απομόνωσης και ταυτοποίησης. Το οικουμενικό δένδρο της ζωής Όλες οι υπάρχουσες μορφές ζωής είναι σύγχρονοι οργανισμοί, καλά προσαρμοσμένοι όσον αφορά τη διαβίωση μέσα στους οικολογικούς θώκους τους. Οι οργανισμοί Aquifex και Methanopyrus αναπτύσσονται σε πολύ υψηλές θερμοκρασίες, συνθήκη που πολύ πιθανόν αντιμετώπιζαν οι πρωτόγονοι συγγενείς αυτών των οργανισμών, οι οποίοι ζούσαν σε μια πιο θερμότερη Γη. Από το φυλογενετικό δένδρο φαίνεται πως το Aquifex και το Methanopyrus είναι πιο κοντά στους πρωτόγονους οργανισμούς απ’ ότι τα ακραία αλόφιλα (Αρχαία).

Οικουμενικό φυλογενετικό δένδρο βάσει των αλληλουχιών του r. RNA.
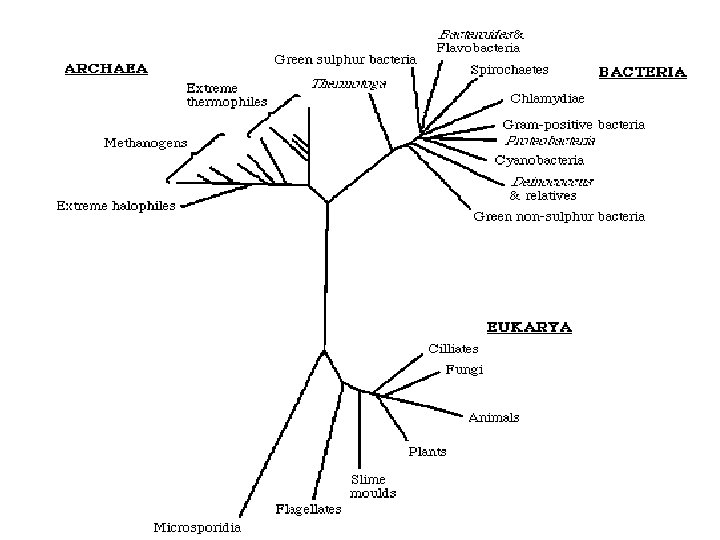
. Πολλά φύλα ορίζονται από")
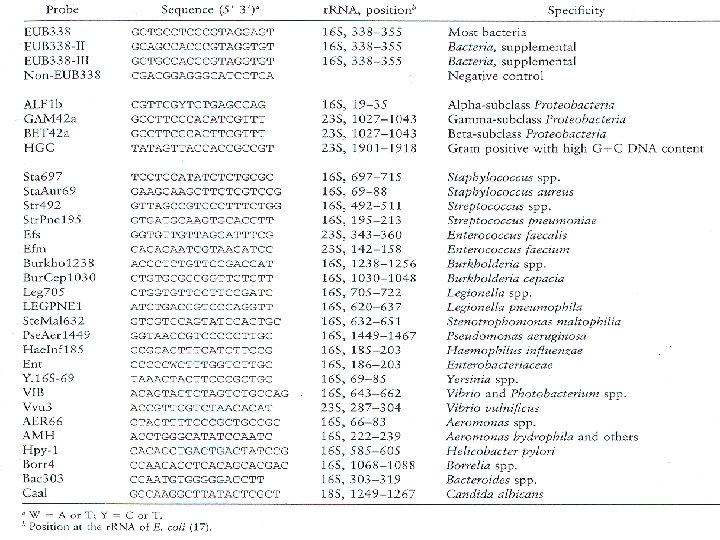
Βακτήρια Μεταξύ των Βακτηρίων έχουν ανακαλυφθεί τουλάχιστον 40 υποδιαιρέσεις (φύλα). Πολλά φύλα ορίζονται από την ανάλυση αλληλουχιών και μόνο. Τα περισσότερα φύλα απαρτίζονται από είδη τα οποία, αν και από φυλογενετική σκοπιά έχουν συσχετισμούς, στερούνται μιας ισχυρής φαινοτυπικής ομοιογένειας. Η φυσιολογία και η φυλογένεση δεν είναι απαραιτήτως συνδεδεμένες. Είναι φανερό ότι τα ευκαρυωτικά οργανίδια έχουν προέλθει από τον «χώρο» των Βακτηρίων. Όπως φαίνεται από το οικουμενικό δένδρο, τα μιτοχόνδρια προήλθαν από τα Πρωτεοβακτήρια. Ο χλωροπλάστης φαίνεται ότι προήλθε από το φύλο των κυανοβακτηρίων, ομάδα βακτηρίων ικανών για οξυγονοπαραγωγό φωτοσύνθεση. Αρχαία Ο «χώρος» των Αρχαίων απαρτίζεται από τρία μεγάλα φύλα, τα Κρεναρχαιωτικά, τα Ευρυαρχαιωτικά και μια τρίτη πιθανή εξελικτική γραμμή που ονομάζεται «Κοραρχαιωτικά» . Τα υπερθερμόφιλα Κρεναρχαιωτικά, όπως το Thermoproteus, το Pyrolobus και το Pyrodictium διακλαδίζονται πολύ κοντά στη ρίζα του οικουμενικού δένδρου. Ακολουθούνται από τα Ευρυαρχαιωτικά, τους μεθανιογόνους προκαρυώτες και τα είδη ακραίων αλόφιλων. Το Thermoplasma, ένας οξεόφιλος, θερμόφιλος προκαρυώτης χωρίς κυτταρικό τοίχωμα. Τα «Κοραρχαιωτικά» είναι υπερθερμόφιλα.

Κυτταρικά τοιχώματα Σχεδόν όλα τα Βακτήρια έχουν κυτταρικά τοιχώματα που περιέχουν πεπτιδογλυκάνη. Οι μόνες γνωστές εξαιρέσεις είναι τα μέλη της ομάδας Planctomyces-Pirella, των οποίων τα κυτταρικά τοιχώματα αποτελούνται από πρωτεΐνη, και τα μέλη των ομάδων Mycoplasma -Chlamydia, που στερούνται εξ ολοκλήρου κυτταρικών τοιχωμάτων. Η πεπτιδογλυκάνη επομένως θεωρείται ως μόριο – «υπογραφή» για πολλά είδη των Βακτηρίων και ουσιαστικά ανιχνεύουμε το μουραμικό οξύ επειδή εμφανίζεται αποκλειστικά και μόνο στην πεπτιδογλυκάνη.

Βακτηριακό τοίχωμα Ε. Coli. Πεπτιδογλυκάνη - επαναλαμβανόμενες μονάδες του τετραπεπτιδίου της γλυκάνης Τα Ευκάρυα και τα Αρχαία στερούνται πεπτιδογλυκάνης. Στους ευκαρυώτες, εάν υπάρχουν κυτταρικά τοιχώματα, αυτά είναι συνήθως κατασκευασμένα από κυτταρίνη ή χιτίνη. Στα Αρχαία, υπάρχουν διάφοροι τύποι κυτταρικών τοιχωμάτων, από το ανάλογο της πεπτιδογλυκάνης, την ψευδοπεπτιδογλυκάνη, μέχρι τα τοιχώματα που κατασκευάζονται από πολυσακχαρίτες, πρωτεΐνη ή γλυκοπρωτεΐνη.

Δομή της ψευδοπεπτιδογλυκάνης ( οι πεπτιδικές συνδέσεις γίνονται με ομάδες Νακετυλοταλοζαμινουρονικού –ΝΑΤ αντί του μουραμικού οξέος).

Λιπίδια Τα Βακτήρια και τα Ευκάρυα συνθέτουν μεμβρανικά λιπίδια, με σκελετό που αποτελείται από λιπαρά οξέα προσδεδεμένα με εστερικό δεσμό σε ένα μόριο γλυκερόλης. Αντιθέτως, τα λιπίδια των Αρχαίων αποτελούνται από μόρια με αιθερικούς δεσμούς (α) : εστερικός δεσμός R: (β) : αιθερικός δεσμός στα λιπίδια των Ευκαρύων και των Βακτηρίων , R= λιπαρά οξέα ενώ στα Αρχαία R =ισοπρένιο.

Στα λιπίδια με εστερικούς δεσμούς, τα λιπαρά οξέα είναι συνήθως μόρια με ευθείες αλυσίδες, ενώ στα Αρχαία, στη θέση των λιπαρών οξέων υπάρχουν υδρογονάνθρακες με μακρές διακλαδισμένες αλυσίδες, είτε φυτανύλιου, είτε του διφυτανύλιου, που συνδέονται με τη γλυκερόλη μέσω αιθερικών δεσμών.

RNA πολυμεράση Η μεταγραφή εκτελείται σε όλους τους οργανισμούς από DNA-εξαρτώμενες RNA πολυμεράσες. Τα κύτταρα των Βακτηρίων περιέχουν έναν απλό τύπο RNA πολυμεράσης με μάλλον απλή τεταρτοταγή δομή. Αυτός ο κλασικός τύπος RNA πολυμεράσης αποτελείται από τέσσερα πολυπεπτίδια, α, β, β΄, και σ, συνδυασμένα σε μια αναλογία 2: 1: 1: 1 αντιστοίχως. Σύνοψη διαφορών Βακτηρίων , Αρχαίων και Ευκαρύων Χαρακτηριστικά Βακτήρια Αρχαία Ευκάρυα Προκαρυωτική κυτταρική δομή Ναι Όχι DNA σε ομοιοπολικά κλειστή κυκλική μορφή Ναι Όχι Παρουσία ιστονών Όχι Ναι Μεμβρανοπερίκλειστος πυρήνας Όχι Ναι Κυτταρικό τοίχωμα Με μουραμικό οξύ Χωρίς μουραμικό οξύ Μεμβρανικά λιπίδια Με εστερικούς δεσμούς Με αιθερικούς δεσμούς Με εστερικούς δεσμούς Ριβοσώματα 70 S 80 S Εναρκτήριο t. RNA Φορμυλομεθειονίνη Μεθειονίνη Ιντρόνια στα περισσότερα γονίδια Όχι Ναι Οπερόνια Ναι Όχι Καλύπτρα και ουρά πολύ-Α του m. RNA Όχι Ναι Πλασμίδια Ναι Σπανίως Ριβοσώματα ευαίσθητα στην τοξίνη της διφθερίτιδας Όχι Ναι RNA πολυμεράσες Μία (4 υπομονάδες) Αρκετές (8 -12 υπομονάδες η καθεμιά) Τρεις (12 -14 υπομονάδες η καθεμιά)
 Πλαίσιο ΤΑΤΑ Ευαισθησία σε")
Δομή υποκινητών Αλληλουχίες – 10 και – 35 (πλαίσιο Pribnow) Πλαίσιο ΤΑΤΑ Ευαισθησία σε χλωραμφαινικόλη, στρεπτομυκίνη και καναμυκίνη Ναι Όχι Μεθανιογένηση Όχι Ναι Όχι Νιτροποίηση Ναι Όχι Απανίτρωση Ναι Όχι Δέσμευση αζώτου Ναι Όχι Φωτοσύνθεση βασισμένη στη χλωροφύλλη Ναι Ναι (στους χλωροπλάστες) Χημειολιθοτροφία (Fe, S, H 2) Ναι Όχι Αεροκυστίδια Ναι Όχι Σύνθεση κοκκίων αποθήκευσης άνθρακα, που συντίθενται από πολύ-β-υδροξυοξέα Ναι Όχι Ανάπτυξη πάνω από τους 110οC Ναι Όχι

Οι RNA πολυμεράσες των Αρχαίων είναι δομικώς πιο περίπλοκες από εκείνες των Βακτηρίων. Περιέχουν οκτώ ή περισσότερα πολυπεπτίδια, μοιάζοντας περισσότερο με τον τύπο που βρίσκουμε στα Ευκάρυα. Η κύρια RNA πολυμεράση των ευκαρυωτών (υπάρχουν τρεις) περιέχει 10 -12 πολυπεπτίδια, και τα σχετικά μεγέθη των πεπτιδίων συμπίπτουν σε μεγαλύτερο βαθμό με εκείνα των ειδών των υπερθερμόφιλων Αρχαίων. Από φυλογενετικής υπογραφής, η πολυμεράση (α 2ββ΄σ) αποτελεί διαγνωστικό στοιχείο για τα Βακτήρια. Χαρακτηριστικά της πρωτεϊνοσύνθεσης Στα Βακτήρια, το κωδικόνιο έναρξης (AUG) επιβάλλει την ενσωμάτωση ενός εναρκτήριου t. RNA που μεταφέρει μια τροποποιημένη ομάδα μεθειονίνης, τη φορμυλομεθειονίνη. Αντιθέτως, στους ευκαρυώτες και στα Αρχαία, το εναρκτήριο t. RNA μεταφέρει μη τροποποιημένη μεθειονίνη. Τα περισσότερα από τα αντιβιοτικά που επιδρούν εξειδικευμένα στη πρωτεϊνοσύνθεση των Βακτηρίων δεν επιδρούν στην πρωτεϊνοσύνθεση των Αρχαίων ή των ευκαρυωτών. Τα αποτελέσματα αυτά υποδεικνύουν ότι οι ριβοσωματικές πρωτεΐνες των Αρχαίων και των Ευκαρύων μοιάζουν από λειτουργική άποψη περισσότερο μεταξύ τους παρά με εκείνες των ριβοσωματικών πρωτεϊνών των Βακτηρίων. Στην κλασική βακτηριακή ταξινομία, μετρούνται αρκετά χαρακτηριστικά για να γίνει ομαδοποίηση των οργανισμών κατά ανιούσα φορά στην ταξινομική κλίμακα. Χαρακτηριστικά με ταξινομική αξία, που χρησιμοποιούνται ευρέως, είναι διάφορες πλευρές της μορφολογίας, της βιοχημείας, της διατροφής κλπ.

. Μορφολογία Σχήμα , μέγεθος , αντίδραση κατά Gram Κινητικότητα Κίνηση με μαστίγια , κίνηση με ολίσθηση , κίνηση με αεροκυστίδια, απουσία κίνησης Θρέψη και φυσιολογία Μηχανισμός διατήρησης ενέργειας (φωτότροφα, χημειοργανότροφα, χημειολιθότροφα) , σχέση με το οξυγόνο , απαιτήσεις/ανοχές ως προς τη θερμοκρασία, το p. H και τα άλατα , ικανότητα εκμετάλλευσης διαφόρων πηγών άνθρακα, αζώτου και θείου Άλλοι παράγοντες κυτταρικά έγκλειστα ή δομές επιφανείας , παθογένεια , ευαισθησία στα αντιβιοτικά

Ο καθορισμός της συνολικής περιεκτικότητας του γονιδιωματικού DNA ενός οργανισμού σε γουανίνη και κυτοσίνη ( η οποία αποκαλείται συνήθως αναλογία GC) , αποτελεί μέρος της κλασικής βακτηριακής ταξινομίας χωρίς ουσιαστική αξία. Οι αναλογίες GC κυμαίνονται σε ένα ευρύ πεδίο, με χαμηλές τιμές από το 20% και ψηλές μέχρι το 80%, στους προκαρυώτες, οι οποίοι έχουν κάπως ευρύτερο πεδίο τιμών απ’ ότι οι ευκαρυώτες. Η γνώση της αναλογίας GC ενός οργανισμού είναι ενίοτε χρήσιμη για την ταυτοποίησή του, αλλά σπανίως καθοριστική. Αυτό συμβαίνει επειδή δύο οργανισμοί είναι δυνατόν να έχουν την ίδια αναλογία GC και όμως να είναι εντελώς άσχετοι μεταξύ τους (τόσο ταξινομικά όσο και φυλογενετικά). Επομένως, οι αναλογίες GC έχουν συνήθως ελάχιστη αξία στον συνολικό ταξινομικό χαρακτηρισμό ενός οργανισμού. Μοριακή ταξινόμηση Υβριδισμός Αν δύο οργανισμοί έχουν τις ίδιες αλληλουχίες νουκλεοτιδίων στο DNA τους, τότε είναι πιθανό να περιέχουν πολλά παραπλήσια, αν όχι πανομοιότυπα, γονίδια. Θα ήταν αναμενόμενο, λοιπόν, δύο μόρια DNA να υβριδίζονται μεταξύ τους με ευκολία ανάλογη προς την ομοιότητα στην αλληλουχία των γονιδίων τους. Ετσι η υβριδοποίηση μετρά τον βαθμό της ομοιότητας των αλληλουχιών και είναι συνεπώς χρήσιμη για τη διάκριση οργανισμών που έχουν στενή συγγένεια. Για να θεωρηθούν δύο απομονωμένοι οργανισμοί ως ίδια είδη, απαιτούνται τιμές υβριδοποίησης μεγαλύτερες της τάξης του 70%, ενώ απαιτούνται για να υποστηριχθεί ότι δύο οργανισμοί ανήκουν στο ίδιο γένος τιμές της τάξης του 20 -30% τουλάχιστον ενώ η υβριδοποίηση μορίων DNA από εντελώς άσχετους οργανισμούς συμβαίνει σε ποσοστό μικρότερο του 10%.

Απεικόνηση της διαδικασίας υβριδισμού σε καθαρή καλλιέργεια βακτηρίων και σε απ’ευθείας ανίχνευση βακτηρίων από κλινικό δείγμα.

Οι φαινοτυπικές μέθοδοι είναι χρονοβόρες και όχι απόλυτα αποτελεσματικές για καθορισμό βακτηρίων τα οποία αναπτύσσονται δύσκολα. Κατά συνέπεια υπάρχει μεγάλη ανάγκη για ανάπτυξη γρηγορότερων και κυρίως πιο αποτελεσματικών μεθόδων Η ανάλυση αλληλουχίας DNA που αντιστοιχεί στο 16 S r. RNA των προκαρυωτικών (r. DNA) είναι η μέθοδος επιλογής καθώς : Το 16 S r. RNA είναι κατάλληλο για χαρακτηρισμό βακτηρίων διότι περιέχει → Συντηρημένες αλληλουχίες ανάμεσα στα είδη των βακτηρίων που μπορούν να χρησιμοποιηθούν για το σχεδιασμό γενικών εκκινητών ( primers) για PCR → Ενδιάμεσες μεταβλητές αλληλουχίες που παρέχουν τον πολυμορφισμό για χαρακτηρισμό σε επίπεδο είδους ή στελέχους Πειραματική διαδικασία: -Aπομόνωση βακτηριακού DNA - PCR για ενίσχυση τμήματος του 16 S r. RNA - ηλεκτροφόρηση των προϊόντων - αλληλούχιση της περιοχής που ενισχύθηκε - ταυτοποίηση μεσω της βάσης δεδομένων (BLAST) - ταξινόμηση

Broad-Range PCR Ένα μόνο ζεύγος primers μπορεί να χρησιμοποιηθεί για την ανίχνευση διαφορετικών, μοναδικών αλληλουχιών. Οι εκκινητές έχουν σχεδιασθεί πάνω σε σταθερές περιοχές του r. RNA οι οποίες περιβάλλονται από μεταβλητές περιοχές. Πλεονεκτεί έναντι των συμβατικών διαγνωστικών μεθόδων γιατί είναι δυνατόν να καθορίσει τη σύσταση ακόμη και πολυμικροβιακών συμπλεγμάτων, όπως σε περιβαλλοντικά δείγματα ή δείγματα μικροβιακής χλωρίδας του ανθρώπου και των ζώων

Συνήθεις εκκινητές οι οποίοι χρησιμοποιούνται για την ανίχνευση Αρχαίων , Βακτηρίων και Ευκαρύων.

Ταξινόμηση με βάση το γένος ή το είδος Η ταξινόμηση βασίζεται στο % ομοιότητας της αλληλουχίας που προκύπτει από την αλληλούχιση των βακτηρίων και σύγκριση των αλληλουχιών με εκείνες που έχουν ήδη καταχωρηθεί στη βάση δεδομένων. Oργανισμοί που ανήκουν στο ίδιο είδος έχουν ομοιότητα > 99% στη 16 S r. RNA. Καθορισμός ριβοτύπου Σε αντίθεση με τις μεθόδους της συγκριτικής ανάλυσης αλληλουχιών, ο καθορισμός του ριβοτύπου δεν περιλαμβάνει τον προσδιορισμό αλληλουχιών, αλλά βασίζεται στο ιδιαίτερο προφίλ ζωνών ηλεκτροφόρησης που παράγονται όταν το DNA ενός συγκεκριμένου οργανισμού υφίσταται τη δράση περιοριστικών ενζύμων. Τα αντίστοιχα θραύσματα διαχωρίζονται και υβριδοποιούνται με έναν ανιχνευτή ριβοσωματικού RNA. Επειδή οι διαφορές στις αλληλουχίες ριβοσωματικού RNA μεταξύ δύο οργανισμών μεταφράζονται σε παρουσία ή απουσία συγκεκριμένων σημείων δράσης των ενζύμων περιορισμού, το συνολικό προφίλ των θραυσμάτων δεδομένου βακτηριακού είδους είναι μοναδικό για το είδος αυτό. Στον καθορισμό του ριβοτύπου έχουμε ως υλικό αφετηρίας είτε ποσότητα DNA είτε DNA που κωδικοποιεί 16 S r. RNA και στην περίπτωση αυτή ενισχύεται με αλυσιδωτή αντίδραση της πολυμεράσης (PCR). Το DNA υφίσταται επεξεργασία πέψης με ένα ή περισσότερα ένζυμα περιορισμού, τα προϊόντα της πέψης διαχωρίζονται με ηλεκτροφόρηση και κατόπιν υβριδοποιούνται

με ειδικό ανιχνευτή. Στη συνέχεια, η εικόνα που σχηματίζουν τα θραύσματα του DNA στο πήκτωμα αναλύεται με το κατάλληλο λογισμικό και γίνεται σύγκριση με αντίστοιχους τύπους πέψης από οργανισμούς αναφοράς σε μια βάση δεδομένων. Ο καθορισμός του ριβοτύπου είναι μια μέθοδος ταυτοποίησης βακτηρίων γρήγορη (αφού παρακάμπτει τον καθαυτό προσδιορισμό αλληλουχιών ) και ειδική. Για τον λόγο αυτό βρίσκει πολλές εφαρμογές στην κλινική διαγνωστική και στις μικροβιακές αναλύσεις τροφίμων, ποτών και νερού. Πέψη του απομονωμένου DNA με περιοριστικές ενδονουκλεάσες Εco. RI, Bam. HI και Hind. III

Automated Ribotyping Riboprinter® Microbial Characterization System Πλήρως αυτοματοποιημένο σύστημα ribotyping που δίνει το γενετικό "αποτύπωμα " οποιουδήποτε βακτηρίου μέσα σε 8 ώρες Το σύστημα δίνει ένα Ribo. Print® πρότυπο, το οποίο το συγκρίνει με άλλα πρότυπα μέσω μιας βάσης δεδομένων για χαρακτηρισμό και ταυτοποίηση Το σύστημα μπορεί να επεξεργαστεί μέχρι 8 βακτηριακά δείγματα την φορά, ενώ μπορεί να δέχεται καινούργια δείγματα κάθε 2 ώρες.

Λήψη δείγματος Προετοιμασία DNA Απομόνωση DNA , πέψη με ένζυμα περιορισμού , Ηλεκτροφορετικός διαχωρισμός Μεταφορά DNA σε νιτροκυτταρίνη και υβριδισμός Επεξεργασία μεμβράνης. Ανίχνευση ζωνών και με το κατάλληλο λογισμικό , ταυτοποίηση στελεχών τα οποία μπορούν να αποθηκευτούν και να συγκριθούν με άλλα στελέχη οπουδήποτε στον κόσμο

Εφαρμογή του αυτόματου Ribotyping σε Gram θετικά και Gram αρνητικά με τα αντίστοιχα ένζυμα περιορισμού

Α. Ribotyping K. pneumoniae. Κλινικά δείγματα 1 -8. S μάρτυρας μοριακού βάρους Β. Η ίδια εικόνα μετά από επεξεργασία με το κατάλληλο λογισμικό. Φαίνεται σαφώς ότι τα κλινικά δείγματα 5 και 6 είναι πανομοιότυπα ενώ τα υπόλοιπα κλινικά στελέχη αν και είναι K. Pneumoniae δεν είναι πανομοιότυπα.

Η έννοια των ειδών στη μικροβιολογία Έχει διατυπωθεί η πρόταση ότι ένας προκαρυώτης του οποίου η αλληλουχία ριβοσωματικού 16 S r. RNA διαφέρει τουλάχιστον κατά 3% από εκείνη όλων των άλλων οργανισμών, θα πρέπει να θεωρείται ως νέο είδος. Η πρόταση αυτή υποστηρίζεται από την παρατήρηση ότι DNA από δύο βακτήρια των οποίων οι αλληλουχίες του ριβοσωματικού 16 S r. RNA είναι όμοιες λιγότερο από 97% κατά κανόνα υβριδοποιούνται σε ποσοστό μικρότερο από το 70%. Η έννοια του είδους είναι σημαντική στη μικροβιολογία, διότι δίνει στα απομονωμένα στελέχη μια τυπική ταξινομική ταυτότητα. Ομάδες ειδών συγκεντρώνονται σε γένη. Διαφορές μεγαλύτερες από 5 -7% (9395% ομοιότητα) στην αλληλουχία 16 S έχει θεωρηθεί ότι υποδεικνύουν ένα νέο γένος. Ομάδες από γένη συγκεντρώνονται σε οικογένειες, οι οικογένειες συγκεντρώνονται σε τάξεις και ούτω καθεξής έως το ανώτερο ταξινομικό επίπεδο. Πως προκύπτουν νέα είδη βακτηριών; Εάν τα κύτταρα του πληθυσμού μοιράζονται δεδομένο ζωτικό πόρο (π. χ. μια βασική θρεπτική ουσία), ο πληθυσμός των κυττάρων μπορεί να ονομασθεί οικότυπος. Στο εσωτερικό ενός δεδομένου οικότυπου, μόλις εμφανιστεί μια προσαρμοστική μετάλλαξη ακολουθεί περιοδική επιλογή και ο παλαιός οικότυπος εξαλείφεται από τον πληθυσμό του νέου οικότυπου. Με τον τρόπο αυτό, οι πληθυσμοί των κυττάρων απομακρύνονται ο ένας από τον άλλο και οι επαναλαμβανόμενοι κύκλοι μεταλλάξεων και επιλογών οδηγούν τελικά σε έναν νέο οικότυπο που είναι αρκετά διαφορετικός γενετικά από τον πρωταρχικό, έτσι ώστε να αναγνωρίζεται ως νέο είδος.
 γονιδιακή ροή –")
Η δημιουργία νέων ειδών βακτηρίων επηρεάζεται επίσης από την οριζόντια (πλευρική) γονιδιακή ροή – τη μεταφορά γονιδίων μεταξύ ειδών με σύζευξη, μεταγωγή και μετασχηματισμό. Ωστόσο πρακτικά, η δημιουργία νέων ειδών θεωρείται μάλλον αποτέλεσμα μετάλλαξης και περιοδικής επιλογής στα γονίδια που υπάρχουν στο γονιδίωμα ενός οργανισμού, παρά απόκτησης νέων γονιδίων με οριζόντια μεταφορά. Σε αντίθεση, τα γονίδια που αποκτώνται με οριζόντια μεταφορά είναι συνήθως αριθμητικά λίγα, κατά κανόνα προσφέρουν μόνον ένα παροδικό πλεονέκτημα και συχνά χάνονται αν μειωθεί η επιλεκτική πίεση για τη διατήρησή τους. Συλλογές καλλιεργειών και δημοσίευση νέων ταξινομικών μονάδων Για να αποκτήσει επίσημη ταξινομική υπόσταση το νέο γένος ή είδος, δημοσιεύεται μια λεπτομερής περιγραφή του απομονωμένου οργανισμού, καθώς και η προτεινόμενη ονομασία τους, ενώ μια βιώσιμη καλλιέργειά του κατατίθεται σε μια συλλογή καλλιεργειών, όπως π. χ. στην ATCC [American Type Culture Collection]. Τα κατατιθέμενα στελέχη αναγνωρίζονται ως τυπικά στελέχη των νέων ειδών και ως το πρότυπο στο οποίο μπορούν να συγκριθούν άλλα στελέχη. Οι συλλογές καλλιεργειών διατηρούν την κατατεθειμένη καλλιέργεια εν γένει καταψύχοντάς την σε πολύ χαμηλές θερμοκρασίες (από – 80ο ως – 196οC) ή με λυοφιλοποίηση (ψυχρή αφυδάτωση).

Στο πεδίο της μικροβιακής ταξινομίας, το Εγχειρίδιο Συστηματικής Βακτηριολογίας του Bergey αποτελεί έργο αναφοράς. Το Εγχειρίδιο του Bergey είναι ένας κατάλογος προτύπων και μοριακών πληροφοριών για όλα τα αναγνωρισμένα είδη προκαρυωτών, κατά τον χρόνο της δημοσίευσής του, και περιέχει πίνακες, εικόνες και άλλες πληροφορίες σχετικές με τη Συστηματική, χρήσιμες για τις ταυτοποιήσεις ενσωματώνοντας πολλές από τις έννοιες που προέκυψαν με την ανάλυση αλληλουχιών του ριβοσωματικού RNA, τις οποίες συμπληρώνει με μια πληθώρα κλασικών ταξινομικών πληροφοριών. Ένα δεύτερο πολύτομο έργο αναφοράς είναι « Οι Προκαρυώτες» (The Prokaryotes). Το έργο αυτό, υπάρχει και σε μορφή προσπελάσιμη από το Διαδίκτυο (http: //www. prokaryotes. com), η οποία ανανεώνεται περιοδικά ώστε να ανταποκρίνεται στον ταχύ ρυθμό με τον οποίο παράγονται νέες πληροφορίες για την ταξινόμηση και τη φυλογένεση των προκαρυωτών.
f5facbaaca344e3f8b99c21064208e16.ppt