

Chloroplasts 2011.ppt
- Количество слайдов: 75
 Ш Хлоропласты: строение, деление и их происхождение Ш Запасные") Ш Пластиды (хлоропласты и амилопласты) Ш Хлоропласты: строение, деление и их происхождение Ш Запасные вещества Ш Пиреноиды и карбоксисомы
Ш Пластиды (хлоропласты и амилопласты) Ш Хлоропласты: строение, деление и их происхождение Ш Запасные вещества Ш Пиреноиды и карбоксисомы
 • У эукариотических водорослей встречается два типа пластид:") Пластиды (клеточные органеллы, несущие собственный геном) • У эукариотических водорослей встречается два типа пластид: хлоропласты (хроматофоры) и амилопласты • Амилопласты представляют собой Амилопласты непигментированные органеллы, присутствующие в некоторых клетках водорослей. Амилопласты отвечают за синтез и последующее хранение гранул крахмала путём полимеризации глюкозы
Пластиды (клеточные органеллы, несущие собственный геном) • У эукариотических водорослей встречается два типа пластид: хлоропласты (хроматофоры) и амилопласты • Амилопласты представляют собой Амилопласты непигментированные органеллы, присутствующие в некоторых клетках водорослей. Амилопласты отвечают за синтез и последующее хранение гранул крахмала путём полимеризации глюкозы
 СТРОЕНИЕ ХЛОРОПЛАСТОВ 1. 2. 1– хлоропласты у разных представителей зеленых водорослей, 2 - строение хлоропласта зеленых водорослей
СТРОЕНИЕ ХЛОРОПЛАСТОВ 1. 2. 1– хлоропласты у разных представителей зеленых водорослей, 2 - строение хлоропласта зеленых водорослей
 • Хлоропла сты — зелёные пластиды, которые встречаются в клетках растений и эукариотических водорослей. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл и являются двумембранными органеллами. Под двойной мембраной локализованы тилакоиды (мембранные образования, в которых находится электрон-транспортная цепь хлоропластов). • Пространство между оболочкой хлоропласта и тилакоидами называется стромой. В строме содержатся хлоропластные молекулы РНК, пластидная ДНК, рибосомы прокариотического типа — 70 S рибосомы, могут откладываться крахмальные зёрна
• Хлоропла сты — зелёные пластиды, которые встречаются в клетках растений и эукариотических водорослей. С их помощью происходит фотосинтез. Хлоропласты содержат хлорофилл и являются двумембранными органеллами. Под двойной мембраной локализованы тилакоиды (мембранные образования, в которых находится электрон-транспортная цепь хлоропластов). • Пространство между оболочкой хлоропласта и тилакоидами называется стромой. В строме содержатся хлоропластные молекулы РНК, пластидная ДНК, рибосомы прокариотического типа — 70 S рибосомы, могут откладываться крахмальные зёрна
 • Хлоропласты обладают собственным геномом, в котором хранятся гены некоторых тилакоидных белков. Однако в процессе эволюции пластид из их предшественников — эндосимбиотических цианобактерий — произошел перенос большого количества генов из хлоропластного генома в ядро клетки. В результате этого четыре основных тилакоидных белковых комплекса частично кодируются в геноме хлоропласта, а частично — ядерным геномом • Для образования тилакоидов требуется белок VIPP 1 (белок пластид 1, индуцирующий везикулярный транспорт, англ. vesicle-inducing protein in plastids 1).
• Хлоропласты обладают собственным геномом, в котором хранятся гены некоторых тилакоидных белков. Однако в процессе эволюции пластид из их предшественников — эндосимбиотических цианобактерий — произошел перенос большого количества генов из хлоропластного генома в ядро клетки. В результате этого четыре основных тилакоидных белковых комплекса частично кодируются в геноме хлоропласта, а частично — ядерным геномом • Для образования тилакоидов требуется белок VIPP 1 (белок пластид 1, индуцирующий везикулярный транспорт, англ. vesicle-inducing protein in plastids 1).
. Геном") Размеры хлоропластной ДНК варьируют от 130 до 160 тыс пар оснований (130 генов). Геном хлоропластов кодирует около половины белков тилакоидной мембраны, ген большой субъединицы ключевого фермента фотосинтеза – Ру. Бис. Ко)
Размеры хлоропластной ДНК варьируют от 130 до 160 тыс пар оснований (130 генов). Геном хлоропластов кодирует около половины белков тилакоидной мембраны, ген большой субъединицы ключевого фермента фотосинтеза – Ру. Бис. Ко)
 и напоминают соответствующие") Тилакоиды • Мембраны тилакоидов похожи на мембраны прокариот (наличием кислых липидов) и напоминают соответствующие органеллы у цианобактерий. На мембранах тилакоидов протекают световые реакции фотосинтеза. Мембрана тилакоидов цианобактерий состоит из складок плазматической мембраны, которые лежат в цитоплазме. • Тилакоиды зеленых водорослей группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл.
Тилакоиды • Мембраны тилакоидов похожи на мембраны прокариот (наличием кислых липидов) и напоминают соответствующие органеллы у цианобактерий. На мембранах тилакоидов протекают световые реакции фотосинтеза. Мембрана тилакоидов цианобактерий состоит из складок плазматической мембраны, которые лежат в цитоплазме. • Тилакоиды зеленых водорослей группируются в граны, которые представляют собой стопки сплюснутых и тесно прижатых друг к другу тилакоидов, имеющих форму дисков. Соединяются граны с помощью ламелл.
 Строение тилакоида Тилакоиды — ограниченные мембраной компартменты, которые находятся внутри хлоропластов эукариотических водорослей или в цитоплазме цианобактерий. Слово «тилакоид» происходит от греческого слова thylakos, означающего «мешочек» . Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Мембрана тилакоида собственно и является тем местом, где протекают светозависимые реакции фотосинтеза. Эти реакции идут при участии фотосинтетических пигментов, расположенных непосредственно на мембране в виде перемежающихся темных и светлых полос шириной около 1 нм. Тилакоиды у хлоропластов зеленых водорослей часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами (от латинского «Granum» стопка монет). Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.
Строение тилакоида Тилакоиды — ограниченные мембраной компартменты, которые находятся внутри хлоропластов эукариотических водорослей или в цитоплазме цианобактерий. Слово «тилакоид» происходит от греческого слова thylakos, означающего «мешочек» . Тилакоиды состоят из мембраны, окружающей просвет тилакоида. Мембрана тилакоида собственно и является тем местом, где протекают светозависимые реакции фотосинтеза. Эти реакции идут при участии фотосинтетических пигментов, расположенных непосредственно на мембране в виде перемежающихся темных и светлых полос шириной около 1 нм. Тилакоиды у хлоропластов зеленых водорослей часто имеют структуру, напоминающую стопку дисков. Эти стопки называют гранами (от латинского «Granum» стопка монет). Граны соединены межграновыми или строматическими тилакоидами (ламеллами) в единое функциональное пространство.
 Тилакоидные мембраны содержат интегральные белки, играющие важную роль в захвате светового фотона в светозависимых фотосинтетических реакциях. На мембране есть четыре основных белковых комплекса: фотосистемы I и II Цитохромный комплекс b 6 f АТФ-синтаза • Фотосистема II в основном встречается у грановых тилакоидов, тогда как фотосистема I и АТФ-синтаза — у строматических тилакоидов, а также у внешних слоев гран. Цитохромный комплекс b 6 f распределен равномерно по всей мембране. • Поскольку две фотосистемы пространственно разделены на тилакоидной мембране, для обмена электронами между ними необходимы подвижные переносчики. В роли таких переносчиков выступают пластохинон и пластоциан. Пластохинон переносит электроны от фотосистемы II к цитохромному комплексу b 6 f, тогда как пластоцианин переносит их от цитохромного комплекса b 6 f к фотосистеме I. • Все вместе эти белки преобразуют энергию света для работы электронтранспортных цепей, которые создают электрохимический потенциал через тилакоидную мембрану, а также синтезируют фосфат никотинамид-аденинового динуклеотида (НАДФ) — продукт конечной окислительно-восстановительной реакции. АТФ-синтаза использует этот электрохимический потенциал для синтеза АТФ в процессе фотофосфорилирования.
Тилакоидные мембраны содержат интегральные белки, играющие важную роль в захвате светового фотона в светозависимых фотосинтетических реакциях. На мембране есть четыре основных белковых комплекса: фотосистемы I и II Цитохромный комплекс b 6 f АТФ-синтаза • Фотосистема II в основном встречается у грановых тилакоидов, тогда как фотосистема I и АТФ-синтаза — у строматических тилакоидов, а также у внешних слоев гран. Цитохромный комплекс b 6 f распределен равномерно по всей мембране. • Поскольку две фотосистемы пространственно разделены на тилакоидной мембране, для обмена электронами между ними необходимы подвижные переносчики. В роли таких переносчиков выступают пластохинон и пластоциан. Пластохинон переносит электроны от фотосистемы II к цитохромному комплексу b 6 f, тогда как пластоцианин переносит их от цитохромного комплекса b 6 f к фотосистеме I. • Все вместе эти белки преобразуют энергию света для работы электронтранспортных цепей, которые создают электрохимический потенциал через тилакоидную мембрану, а также синтезируют фосфат никотинамид-аденинового динуклеотида (НАДФ) — продукт конечной окислительно-восстановительной реакции. АТФ-синтаза использует этот электрохимический потенциал для синтеза АТФ в процессе фотофосфорилирования.
 • Фотосистемы тилакоида — центры осуществления окислительно-восстановительных светозависимых реакций. Каждая фотосистема содержит антенный комплекс, который улавливает свет различных длин волн с использованием хлорофилла и вспомогательных фотосинтетических пигментов, таких как каротиноиды и фикобилипротеины. На антенном комплексе имеется от 250 до 400 молекул пигмента. Поглощаемая ими энергия за счет резонансного переноса передается специализированному хлорофиллу a, расположенному в реакционном центре каждой фотосистемы. Когда любая из двух молекул хлорофилла a в реакционном центре получает энергию, электрон передается молекулеакцептору.
• Фотосистемы тилакоида — центры осуществления окислительно-восстановительных светозависимых реакций. Каждая фотосистема содержит антенный комплекс, который улавливает свет различных длин волн с использованием хлорофилла и вспомогательных фотосинтетических пигментов, таких как каротиноиды и фикобилипротеины. На антенном комплексе имеется от 250 до 400 молекул пигмента. Поглощаемая ими энергия за счет резонансного переноса передается специализированному хлорофиллу a, расположенному в реакционном центре каждой фотосистемы. Когда любая из двух молекул хлорофилла a в реакционном центре получает энергию, электрон передается молекулеакцептору.
 ФОТОСИНТЕЗ
ФОТОСИНТЕЗ
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ ХЛОРОФИЛЛЫ У водорослей существует 4 хлорофилла: a, b, c (с в трех формах – с1, с2 и с3) и d. Структура хлорофиллов (Рис. 1). Спектр поглощения: от 650 до 750 нм (оранжевый, красный). Расположение: на фотосинтетических мембранах. Функция: поглощение света и участие в процессе фотосинтеза. 1, 2 – формулы хлорофиллов 2.
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ ХЛОРОФИЛЛЫ У водорослей существует 4 хлорофилла: a, b, c (с в трех формах – с1, с2 и с3) и d. Структура хлорофиллов (Рис. 1). Спектр поглощения: от 650 до 750 нм (оранжевый, красный). Расположение: на фотосинтетических мембранах. Функция: поглощение света и участие в процессе фотосинтеза. 1, 2 – формулы хлорофиллов 2.
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ КАРОТИНОИДЫ Различают 2 основные группы – каротины и ксантофиллы. Структура каротиноидов: (С 40 Н 56) (Рис. 1). Спектр поглощения: от 450 до 500 нм (синий, голубой). Расположение: на фотосинтетических мембранах, рядом с хлорофиллами. Функции: 1. Главная функция состоит в том, чтобы предотвратить образование триплетного состояния хлорофилла и предотвратить образования опасного синглетного кислорода. 2. Каротиноиды улавливают свет более короткой длины волны по сравнению с хлорофиллами и осуществляют передачу возбуждения на фоторецептор. 1. 1– формулы каротиноидов
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ КАРОТИНОИДЫ Различают 2 основные группы – каротины и ксантофиллы. Структура каротиноидов: (С 40 Н 56) (Рис. 1). Спектр поглощения: от 450 до 500 нм (синий, голубой). Расположение: на фотосинтетических мембранах, рядом с хлорофиллами. Функции: 1. Главная функция состоит в том, чтобы предотвратить образование триплетного состояния хлорофилла и предотвратить образования опасного синглетного кислорода. 2. Каротиноиды улавливают свет более короткой длины волны по сравнению с хлорофиллами и осуществляют передачу возбуждения на фоторецептор. 1. 1– формулы каротиноидов
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ ФИКОБИЛИПРОТЕИНЫ Различают 3 основные группы фикобилинов: фикоэритрин, фикоцианин и аллофикоцианин. Структура фикобилинов (Рис. 1). Спектр поглощения: от 545 до 675 нм (зеленый, желтый, красный). Расположение: на фотосинтетических мембранах (у красных и синезеленых водорослей образуют фикобилисомы (Рис. 2)). Функция: Улавливание коротковолновой части спектра и передача возбуждения на хлорофиллы. 1– формулы фикобилинов, 2 - строение фикобилисомы 1. 2.
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ ФИКОБИЛИПРОТЕИНЫ Различают 3 основные группы фикобилинов: фикоэритрин, фикоцианин и аллофикоцианин. Структура фикобилинов (Рис. 1). Спектр поглощения: от 545 до 675 нм (зеленый, желтый, красный). Расположение: на фотосинтетических мембранах (у красных и синезеленых водорослей образуют фикобилисомы (Рис. 2)). Функция: Улавливание коротковолновой части спектра и передача возбуждения на хлорофиллы. 1– формулы фикобилинов, 2 - строение фикобилисомы 1. 2.
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ
 1. ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ ДИНОФИТОВЫХ ВОДОРОСЛЕЙ 2. 4. 3. 1– 2 - структуры хлорофиллов, 3 - структура перидинина 4 - хлоропласт Glenodinium (CE- оболочка хлоропласта, TH- тилакоиды, L- ламелла, PY- пиреноид, PC- запасной продукт)
1. ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ ДИНОФИТОВЫХ ВОДОРОСЛЕЙ 2. 4. 3. 1– 2 - структуры хлорофиллов, 3 - структура перидинина 4 - хлоропласт Glenodinium (CE- оболочка хлоропласта, TH- тилакоиды, L- ламелла, PY- пиреноид, PC- запасной продукт)
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ ОХРОФИТОВЫХ ВОДОРОСЛЕЙ 1. 2. 5. 3. 4. 5 - хлоропласт Ectocarpus (CE- оболочка хлоропласта, CERхлоропластный ЭПР, GLопоясывающая ламелла, L- 3 -х тилакоидная ламелла, PY- пиреноид) 1– 2 - структуры хлорофиллов, 3 - структура фукоксантина, 4 - структура вошериаксантина
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ ОХРОФИТОВЫХ ВОДОРОСЛЕЙ 1. 2. 5. 3. 4. 5 - хлоропласт Ectocarpus (CE- оболочка хлоропласта, CERхлоропластный ЭПР, GLопоясывающая ламелла, L- 3 -х тилакоидная ламелла, PY- пиреноид) 1– 2 - структуры хлорофиллов, 3 - структура фукоксантина, 4 - структура вошериаксантина
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ 1. 4. 2. 3. 1– структуры хлорофиллов, 2 - структура неоксантина, 3 - структура β-каротина 4 - хлоропласт Euglena (CER- оболочка хлоропласта, Py- пиреноид, Pa- парамилон)
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ 1. 4. 2. 3. 1– структуры хлорофиллов, 2 - структура неоксантина, 3 - структура β-каротина 4 - хлоропласт Euglena (CER- оболочка хлоропласта, Py- пиреноид, Pa- парамилон)
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ ЗЕЛЕНЫХ ВОДОРОСЛЕЙ 1. 2. 3. 1 - строение хлорофиллов a и b, 2 -3 - схемы возможных путей синтеза ксантофиллов зеленых водорослей
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ ЗЕЛЕНЫХ ВОДОРОСЛЕЙ 1. 2. 3. 1 - строение хлорофиллов a и b, 2 -3 - схемы возможных путей синтеза ксантофиллов зеленых водорослей
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ФОТОСИНТЕТИЧЕСКИЕ МЕМБРАНЫ CYANOPHYTA 1. 4. 2. 3. 6. 5. 1 - Synechocystis, 2 - Prochloron, 3 - Acaryochloris marina, 4 - одиночные тилакоиды и фикобилисомы, 5 - строение фикобилисомы, 6 - формулы хлорофиллов а и d и их спектры поглощения
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ФОТОСИНТЕТИЧЕСКИЕ МЕМБРАНЫ CYANOPHYTA 1. 4. 2. 3. 6. 5. 1 - Synechocystis, 2 - Prochloron, 3 - Acaryochloris marina, 4 - одиночные тилакоиды и фикобилисомы, 5 - строение фикобилисомы, 6 - формулы хлорофиллов а и d и их спектры поглощения
 Мембраны тилакоидов цианобактерий Цианобактерии — фотосинтетические прокариоты, обладающие высокодифференцированными мембранными системами. У этих бактерий имеется внутренняя система тилакоидных мембран, на которых расположены все компоненты действующих электронтранспортных цепей фотосинтеза и дыхания. Для цианобактерий характерно наличие на внешней стороне мембран тилакоидов сложных белковых комплексов — фикобилисом, которые состоят в основном из белков-фикобилинов фикоциана и фикоэритрина. Из эукариот фикобилисомы есть у красных водорослей и глаукофитовых. Фикобилины поглощают световую энергию и передают её к хлорофиллу а фотосистемы II. За счет наличия таких систем фотосинтетический аппарат этих бактерий сложнее, чем у других групп фотобактерий. Им нужно уметь управлять этими мембранами, синтезировать специфические мембранные липиды, а также обеспечивать правильный транспорт белков. Плазматическая мембрана, окружающая клетку, и мембраны тилакоидов играют разные роли в клетках цианобактерий. Организация, функции, состав и динамика белков мембранных систем остается одной из наиболее активно изучаемых областей в биологии цианобактерий. Строение фикобилисомы
Мембраны тилакоидов цианобактерий Цианобактерии — фотосинтетические прокариоты, обладающие высокодифференцированными мембранными системами. У этих бактерий имеется внутренняя система тилакоидных мембран, на которых расположены все компоненты действующих электронтранспортных цепей фотосинтеза и дыхания. Для цианобактерий характерно наличие на внешней стороне мембран тилакоидов сложных белковых комплексов — фикобилисом, которые состоят в основном из белков-фикобилинов фикоциана и фикоэритрина. Из эукариот фикобилисомы есть у красных водорослей и глаукофитовых. Фикобилины поглощают световую энергию и передают её к хлорофиллу а фотосистемы II. За счет наличия таких систем фотосинтетический аппарат этих бактерий сложнее, чем у других групп фотобактерий. Им нужно уметь управлять этими мембранами, синтезировать специфические мембранные липиды, а также обеспечивать правильный транспорт белков. Плазматическая мембрана, окружающая клетку, и мембраны тилакоидов играют разные роли в клетках цианобактерий. Организация, функции, состав и динамика белков мембранных систем остается одной из наиболее активно изучаемых областей в биологии цианобактерий. Строение фикобилисомы
 СТРОЕНИЕ ХЛОРОПЛАСТОВ
СТРОЕНИЕ ХЛОРОПЛАСТОВ
 Империя Eubacteria Отдел Cyanophyta или Cyanobacteria Название") ЦИАНОБАКТЕРИИ или СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ (CYANOPHYTA или CYANOBACTERIA) Империя Eubacteria Отдел Cyanophyta или Cyanobacteria Название отдела происходит от греч. cyanos - синий
ЦИАНОБАКТЕРИИ или СИНЕЗЕЛЕНЫЕ ВОДОРОСЛИ (CYANOPHYTA или CYANOBACTERIA) Империя Eubacteria Отдел Cyanophyta или Cyanobacteria Название отдела происходит от греч. cyanos - синий
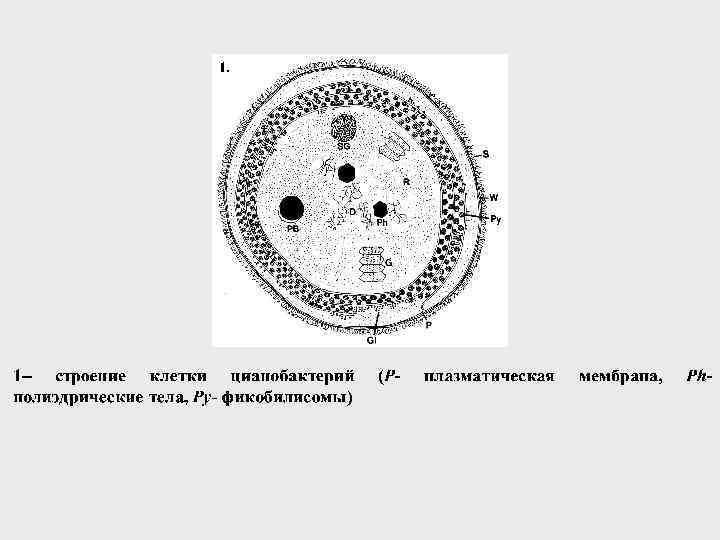
 Империя Plantae Отдел Rhodophyta sensu lato") КРАСНЫЕ ВОДОРОСЛИ или БАГРЯНКИ (RHODOPHYTA S. L. ) Империя Plantae Отдел Rhodophyta sensu lato Название отдела происходит от греч. rhodon - розовый
КРАСНЫЕ ВОДОРОСЛИ или БАГРЯНКИ (RHODOPHYTA S. L. ) Империя Plantae Отдел Rhodophyta sensu lato Название отдела происходит от греч. rhodon - розовый
 ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ 1. 2. 4. 3. 3. 1 -2 - строение хлоропласта красных водорослей: CHL- хлоропласт, CE- оболочка хлоропласта, PTH- периферические тилакоиды (стрелками на Рис. 2 показаны фикобилисомы), 3 - строение фикобилисомы 4 - схема хлоропластов красных водорослей
ФОТОСИНТЕТИЧЕСКИЕ ПИГМЕНТЫ И ХЛОРОПЛАСТЫ 1. 2. 4. 3. 3. 1 -2 - строение хлоропласта красных водорослей: CHL- хлоропласт, CE- оболочка хлоропласта, PTH- периферические тилакоиды (стрелками на Рис. 2 показаны фикобилисомы), 3 - строение фикобилисомы 4 - схема хлоропластов красных водорослей
 и при освещениии (B).") Ультраструктура клеток Porphyra в темноте (A) и при освещениии (B).
Ультраструктура клеток Porphyra в темноте (A) и при освещениии (B).
 Империя Chromalveolates Отдел Cryptophyta Cryptomonas") КРИПТОФИТОВЫЕ ВОДОРОСЛИ или КРИПТОМОНАДЫ (CRYPTOPHYTA) Империя Chromalveolates Отдел Cryptophyta Cryptomonas
КРИПТОФИТОВЫЕ ВОДОРОСЛИ или КРИПТОМОНАДЫ (CRYPTOPHYTA) Империя Chromalveolates Отдел Cryptophyta Cryptomonas
 Хлоропласт криптофитовых • Хлорофилл а и с2 • Каротиноиды и фикобилипротеиновые пигменты, локализованные внутри тилакоидов. В клетках присутствует или фикоциан или фикоэритрин • Запасной продукт – крахмал, откладывается в перипластидном пространстве • Хлоропласт одет 4 мембранами, ламеллы двух- или трехтилакоидные. ДНКхл рассеяна в строме хлоропласта
Хлоропласт криптофитовых • Хлорофилл а и с2 • Каротиноиды и фикобилипротеиновые пигменты, локализованные внутри тилакоидов. В клетках присутствует или фикоциан или фикоэритрин • Запасной продукт – крахмал, откладывается в перипластидном пространстве • Хлоропласт одет 4 мембранами, ламеллы двух- или трехтилакоидные. ДНКхл рассеяна в строме хлоропласта
 Империя Excavates Отдел Euglenophyta Название отдела происходит от греч.") ЭВГЛЕНОВЫЕ ВОДОРОСЛИ (EUGLENOPHYTA или EUGLENOZOA) Империя Excavates Отдел Euglenophyta Название отдела происходит от греч. eu – хороший, glene - глаз
ЭВГЛЕНОВЫЕ ВОДОРОСЛИ (EUGLENOPHYTA или EUGLENOZOA) Империя Excavates Отдел Euglenophyta Название отдела происходит от греч. eu – хороший, glene - глаз
 Империя Chromalveolates Отдел Dinophyta Название отдела происходит от") ДИНОФИТОВЫЕ ВОДОРОСЛИ, ДИНОФИТЫ или ДИНОФЛАГЕЛЛАТЫ (DINOPHYTA) Империя Chromalveolates Отдел Dinophyta Название отдела происходит от греч. dineo – вращаться
ДИНОФИТОВЫЕ ВОДОРОСЛИ, ДИНОФИТЫ или ДИНОФЛАГЕЛЛАТЫ (DINOPHYTA) Империя Chromalveolates Отдел Dinophyta Название отдела происходит от греч. dineo – вращаться
 Хлоропласт эвгленовых и динофитовых
Хлоропласт эвгленовых и динофитовых
 Хлоропласты в клетках Euglena cuneata P - пиреноид Pa - парамилон
Хлоропласты в клетках Euglena cuneata P - пиреноид Pa - парамилон
 Хлоропласты в клетках Euglena deses
Хлоропласты в клетках Euglena deses
 Хлоропласты в клетках Euglena mutabilis
Хлоропласты в клетках Euglena mutabilis
 Хлоропласты охрофитовых 3 1. Таллом Laminaria. 2. Таллом Fucus 3
Хлоропласты охрофитовых 3 1. Таллом Laminaria. 2. Таллом Fucus 3
 BROWN ALGA EGREGIA MENZIESII
BROWN ALGA EGREGIA MENZIESII
 Империя Plantae Отделы Chlorophyta и Charophyta Название") ЗЕЛЕНЫЕ и ХАРОВЫЕ ВОДОРОСЛИ (CLOROPHYTA и CHAROPHYTA) Империя Plantae Отделы Chlorophyta и Charophyta Название отдела происходит от греч. chloro - зеленый
ЗЕЛЕНЫЕ и ХАРОВЫЕ ВОДОРОСЛИ (CLOROPHYTA и CHAROPHYTA) Империя Plantae Отделы Chlorophyta и Charophyta Название отдела происходит от греч. chloro - зеленый
 Схема строения хлоропласта зеленых водорослей
Схема строения хлоропласта зеленых водорослей
 Chlamydomonas и Oocystis marssonii
Chlamydomonas и Oocystis marssonii
 ДНК ядер и хлоропластов у хламидомонады Фельген Акридиновый оранжевый
ДНК ядер и хлоропластов у хламидомонады Фельген Акридиновый оранжевый
 Хлоропластная сеть Acetabularia Окрашивание Di. OC 6. Заметно свечение оболочки хлоропласта и тонкие связующие филаменты между хлоропластами (стрелки). Bar: 10 мкм
Хлоропластная сеть Acetabularia Окрашивание Di. OC 6. Заметно свечение оболочки хлоропласта и тонкие связующие филаменты между хлоропластами (стрелки). Bar: 10 мкм
") Продольные срезы клеток Scherffelia dubia (CHLOROPHYTA)
Продольные срезы клеток Scherffelia dubia (CHLOROPHYTA)
 и на свету (B)") Ультраструктура клеток Ulva, культивируемых в темноте (A) и на свету (B)
Ультраструктура клеток Ulva, культивируемых в темноте (A) и на свету (B)
 ПРОИСХОЖДЕНИЕ ХЛОРОПЛАСТОВ 1. 1– возникновение хлоропластов в результате первичного и вторичного эндосимбиозов
ПРОИСХОЖДЕНИЕ ХЛОРОПЛАСТОВ 1. 1– возникновение хлоропластов в результате первичного и вторичного эндосимбиозов
 ПРОИСХОЖДЕНИЕ ХЛОРОПЛАСТОВ Жирным шрифтом выделены отделы водорослей, которые изучают на малом практикуме
ПРОИСХОЖДЕНИЕ ХЛОРОПЛАСТОВ Жирным шрифтом выделены отделы водорослей, которые изучают на малом практикуме
 Происхождение хлоропластов у разных групп водорослей
Происхождение хлоропластов у разных групп водорослей
 Симбиоз и эволюция эукариотических водорослей. Представлены примеры типов эндосимбиотических событий, ответственных за множественное приобретение фотосинтеза у эукариот показано в виде схем. A, Происхождение симбиотических событий, по-видимому, происходили между примитивным жгутиковым протистов (n 1 представленным ядром протиста) и цианобактерией (cy). Потеря генов цианобактерии и перенос части генов в геном ядра привели к формированию облигатного внутриклеточного хлоропласта (ch). B, Гипотетический предшественник гетеротрофный страменопил (ядро представлено n 2) поглощает фотоавтотрофную эукариотическую водоросль. Происходит перенос генов между n 1 и n 2, приводит к потере n 1 и становится хромофитом. C, Гипотетический гетеротрофный динофлагеллят (n 3) поглощает водоросль. В этом примере n 2 остается. В других случаях симбиоза происходит перенос генов между n 2 и n 3, что приводит к редукции (нуклеоморф) или полной утрате n 2 D, Симбиоз макроводоросли Vausheria litorea с Elysia chlorotica является результатом сосания молодых морских личинок на филаментах водоросли. V. litorea утрачивает ядра (n 2), и остаются только хлоропласты (ch) в цитоплазме клеток моллюска.
Симбиоз и эволюция эукариотических водорослей. Представлены примеры типов эндосимбиотических событий, ответственных за множественное приобретение фотосинтеза у эукариот показано в виде схем. A, Происхождение симбиотических событий, по-видимому, происходили между примитивным жгутиковым протистов (n 1 представленным ядром протиста) и цианобактерией (cy). Потеря генов цианобактерии и перенос части генов в геном ядра привели к формированию облигатного внутриклеточного хлоропласта (ch). B, Гипотетический предшественник гетеротрофный страменопил (ядро представлено n 2) поглощает фотоавтотрофную эукариотическую водоросль. Происходит перенос генов между n 1 и n 2, приводит к потере n 1 и становится хромофитом. C, Гипотетический гетеротрофный динофлагеллят (n 3) поглощает водоросль. В этом примере n 2 остается. В других случаях симбиоза происходит перенос генов между n 2 и n 3, что приводит к редукции (нуклеоморф) или полной утрате n 2 D, Симбиоз макроводоросли Vausheria litorea с Elysia chlorotica является результатом сосания молодых морских личинок на филаментах водоросли. V. litorea утрачивает ядра (n 2), и остаются только хлоропласты (ch) в цитоплазме клеток моллюска.

 Жизнь и смерть Elysia chlototica • Фотография слева это морская личинка рода Elysia, которая выглядит как маленький лист. Зеленый цвет, который создает камуфляж связан с присутствием хлоропластов водоросли, которые попадают во время питания личинки. Использование пигментов хлоропластов для приобретения зеленого окрашивания распространенное явление среди морских личинок. В тоже время у большинства морских личинок хлоропласты перестают работать в течение несколких дней или недель после захвата, в то время как у Elysia chlorotica они могут оставаться стабильными и активными в течение 10 месяцев. В результате личинка представляет собой редкий пример фотосинтетического животного и благодаря этому они могут долгое время жить на свету и СО 2 долгое время.
Жизнь и смерть Elysia chlototica • Фотография слева это морская личинка рода Elysia, которая выглядит как маленький лист. Зеленый цвет, который создает камуфляж связан с присутствием хлоропластов водоросли, которые попадают во время питания личинки. Использование пигментов хлоропластов для приобретения зеленого окрашивания распространенное явление среди морских личинок. В тоже время у большинства морских личинок хлоропласты перестают работать в течение несколких дней или недель после захвата, в то время как у Elysia chlorotica они могут оставаться стабильными и активными в течение 10 месяцев. В результате личинка представляет собой редкий пример фотосинтетического животного и благодаря этому они могут долгое время жить на свету и СО 2 долгое время.
 Деление хлоропласта мхи папортники Эволюция машины деления хлоропласта. Комплекс деления водоросли и наземных растений включают Fts. Z и ARC 6, оба являются участниками машины деления цианобактерий. DR P 5 B добавляется после эндосимбиоза. PDV 2 был приобретен общим предком наземных растений. PDV 1 и PARC 6 появляются при дупликации гена PDV 2 и ARC 6, соответственно, у общего предка сосудистых растений (папортников и семенных растений). Место кольца Fts. Z детерминируется белками Min. D и Min. E которые берут происхождение от цианобактерий, белок ARC 3 является специфичным для зеленых водорослей и наземных растений, и специфичный для наземных растений MCD 1. Вставочная диаграмма показывает возникновение комплекса деления хлоропласта у зеленых водорослей и наземных растений.
Деление хлоропласта мхи папортники Эволюция машины деления хлоропласта. Комплекс деления водоросли и наземных растений включают Fts. Z и ARC 6, оба являются участниками машины деления цианобактерий. DR P 5 B добавляется после эндосимбиоза. PDV 2 был приобретен общим предком наземных растений. PDV 1 и PARC 6 появляются при дупликации гена PDV 2 и ARC 6, соответственно, у общего предка сосудистых растений (папортников и семенных растений). Место кольца Fts. Z детерминируется белками Min. D и Min. E которые берут происхождение от цианобактерий, белок ARC 3 является специфичным для зеленых водорослей и наземных растений, и специфичный для наземных растений MCD 1. Вставочная диаграмма показывает возникновение комплекса деления хлоропласта у зеленых водорослей и наземных растений.
 Хлоропласты возникли из цианобактериального эндосимбионта и остались в эукариотических клетках более чем на один биллион лет благодаря делению хлоропластов и наследованию дочерними клетками в процессе деления. Последние исследования относительно деления хлоропласта показали, что деление хлоропласта представлено крупным белковым комплексом в месте деления, охватывающие как внутреннюю, так и наружную мембраны оболочки. Комплекс деления имеет несколько белков компонентов машины деления цианобактерий, остальные компоненты из клеток хозяина. На основе информации о комплексе деления можно попытаться понять как комплекс деления эволюционировал и как клетки эукариотического хозяина регулируют деление хлоропласта в процессе пролиферации и дифференциации.
Хлоропласты возникли из цианобактериального эндосимбионта и остались в эукариотических клетках более чем на один биллион лет благодаря делению хлоропластов и наследованию дочерними клетками в процессе деления. Последние исследования относительно деления хлоропласта показали, что деление хлоропласта представлено крупным белковым комплексом в месте деления, охватывающие как внутреннюю, так и наружную мембраны оболочки. Комплекс деления имеет несколько белков компонентов машины деления цианобактерий, остальные компоненты из клеток хозяина. На основе информации о комплексе деления можно попытаться понять как комплекс деления эволюционировал и как клетки эукариотического хозяина регулируют деление хлоропласта в процессе пролиферации и дифференциации.
 Часть машины деления, которая происходит от эукариот вовлекает специфические белки члены семейства динаминов. Каждый член семейства динаминов вовлечен в деление или слияние разных эукариотических мембранных систем. Взглянув на тип предшественника динаминового белка и активность эукариотических мембран, которые вовлечены в регуляцию деления хлоропласта, были исследованы функции белков динаминов, которые тесно связаны с белками деления хлоропластов. Эти белки у амебы Dictyostelium discoideum и Arabidopsis thaliana локализованы в местах клеточного деления, где они вовлечены в цитокинез. Наши результаты предполагают, что динамин для деления хлоропласта происходит из машины цитокинеза эукариот. Поэтому машина деления хлоропласта является смесью компонентов цитокинеза бактерий и эукариот с поздним ключевым фактором в синхронизации деления эндосимбионта и деления хозяина.
Часть машины деления, которая происходит от эукариот вовлекает специфические белки члены семейства динаминов. Каждый член семейства динаминов вовлечен в деление или слияние разных эукариотических мембранных систем. Взглянув на тип предшественника динаминового белка и активность эукариотических мембран, которые вовлечены в регуляцию деления хлоропласта, были исследованы функции белков динаминов, которые тесно связаны с белками деления хлоропластов. Эти белки у амебы Dictyostelium discoideum и Arabidopsis thaliana локализованы в местах клеточного деления, где они вовлечены в цитокинез. Наши результаты предполагают, что динамин для деления хлоропласта происходит из машины цитокинеза эукариот. Поэтому машина деления хлоропласта является смесью компонентов цитокинеза бактерий и эукариот с поздним ключевым фактором в синхронизации деления эндосимбионта и деления хозяина.
, chloroplasts of Cyanophora paradoxa NIES-547") Делящиеся цианобактерии и хлоропласты. Aphanothece sp. CMP 2529 (cyanobacterium), chloroplasts of Cyanophora paradoxa NIES-547 (unicellular glaucophyte alga), Chlorella vulgaris C-27 (unicellular green alga), Ulva sp. (multicellulargreenalga), and Physcomitrella patens (мох, наземное растение). Arrowheads indicate the dividing chloroplasts of Ulva and Chlorella. Scalebars=10 mm.
Делящиеся цианобактерии и хлоропласты. Aphanothece sp. CMP 2529 (cyanobacterium), chloroplasts of Cyanophora paradoxa NIES-547 (unicellular glaucophyte alga), Chlorella vulgaris C-27 (unicellular green alga), Ulva sp. (multicellulargreenalga), and Physcomitrella patens (мох, наземное растение). Arrowheads indicate the dividing chloroplasts of Ulva and Chlorella. Scalebars=10 mm.
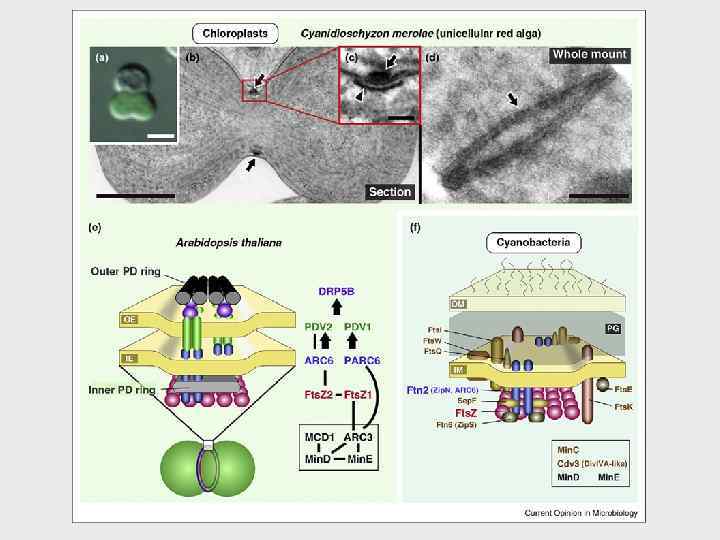
–(d) Микрофотографии показывают деление хлоропласта и деление хлоропласта") Деление комплексов хлоропластов и цианобактерий. (a)–(d) Микрофотографии показывают деление хлоропласта и деление хлоропласта одноклеточной красной водоросли Cyanidioschyzon merolae. (a) Клетка содержит делящийся хлоропласт. (b) Поперечный срез делящегося хлоропласта. (c) Увеличенное фото среза хлоропласта деления комплекса. (d) Фото полученное негативным окрашиванием наружного PD кольца. (e) Диаграмма показывающая способ деления комплекса хлоропласта у Arabidopsis thaliana. Показаны только известные компоненты машины деления. Порядок набора показан стрелками и полоска показывает, что непосредственная связь между двумя белками показана экспериментально. Показано взаимоотношение между кольцом PD и другими компонентами у C. merolae. Другие белки (MSL, GC 1 and CDT 1) предположительно также вовлечены в деление хлоропласта, но их взаимоотношения с комплексом деления пока неизвестны. (f) Подобная диаграмма, показывающая деление комплекса цианобактерии. Локализация Ftn 2, Sep. F, Fts. Z, и Ftn 6 в месте деления показана экспериментально. Локализация Fts. E, Fts. I, Fts. K, Fts. Q, и Fts. W не определены у цианобактерии, но эти белки вовлечены в комплекс деления у других видов бактерий. Arrow, outer PD ring; arrow head, inner PD ring. Scale bars=2 mm (a), 500 nm(b, d), and 50 nm(c).
Деление комплексов хлоропластов и цианобактерий. (a)–(d) Микрофотографии показывают деление хлоропласта и деление хлоропласта одноклеточной красной водоросли Cyanidioschyzon merolae. (a) Клетка содержит делящийся хлоропласт. (b) Поперечный срез делящегося хлоропласта. (c) Увеличенное фото среза хлоропласта деления комплекса. (d) Фото полученное негативным окрашиванием наружного PD кольца. (e) Диаграмма показывающая способ деления комплекса хлоропласта у Arabidopsis thaliana. Показаны только известные компоненты машины деления. Порядок набора показан стрелками и полоска показывает, что непосредственная связь между двумя белками показана экспериментально. Показано взаимоотношение между кольцом PD и другими компонентами у C. merolae. Другие белки (MSL, GC 1 and CDT 1) предположительно также вовлечены в деление хлоропласта, но их взаимоотношения с комплексом деления пока неизвестны. (f) Подобная диаграмма, показывающая деление комплекса цианобактерии. Локализация Ftn 2, Sep. F, Fts. Z, и Ftn 6 в месте деления показана экспериментально. Локализация Fts. E, Fts. I, Fts. K, Fts. Q, и Fts. W не определены у цианобактерии, но эти белки вовлечены в комплекс деления у других видов бактерий. Arrow, outer PD ring; arrow head, inner PD ring. Scale bars=2 mm (a), 500 nm(b, d), and 50 nm(c).
 Распространение известных компонентов машины деления цианобактерий и хлоропластов
Распространение известных компонентов машины деления цианобактерий и хлоропластов
 зависимое от клеточного цикла деление") Регуляция деления хлоропластов у водорослей и наземных растений а-б) зависимое от клеточного цикла деление хлоропласта у клеток водорослей с-d) Регуляция деления хлоропласта PDV белками у наземных растений. а) деление хлоропласта красной водоросли C. merolae. с) деление хлоропласта Arabidopsis thaliana.
Регуляция деления хлоропластов у водорослей и наземных растений а-б) зависимое от клеточного цикла деление хлоропласта у клеток водорослей с-d) Регуляция деления хлоропласта PDV белками у наземных растений. а) деление хлоропласта красной водоросли C. merolae. с) деление хлоропласта Arabidopsis thaliana.
 Хлоропласт делится один раз во") Регуляция деления хлоропласта у водорослей и наземных растений. (A) Хлоропласт делится один раз во время деления клеточного цикла хозяина у водорослей. (B) Скорость деления хлоропласта изменяется во время клеточной дифференциации наземных растений. Пропластиды активно делятся в меристематичеких клетках, постепенно скорость деления снижается по мере созревания хлоропластов. Уровень PDV, но не других компонентов деления снижается параллельно скорости их деления.
Регуляция деления хлоропласта у водорослей и наземных растений. (A) Хлоропласт делится один раз во время деления клеточного цикла хозяина у водорослей. (B) Скорость деления хлоропласта изменяется во время клеточной дифференциации наземных растений. Пропластиды активно делятся в меристематичеких клетках, постепенно скорость деления снижается по мере созревания хлоропластов. Уровень PDV, но не других компонентов деления снижается параллельно скорости их деления.
") Fts. Z – прокариотический цитоскелетный белок, локализованный в строме хлоропласта (близок к тубулину)
Fts. Z – прокариотический цитоскелетный белок, локализованный в строме хлоропласта (близок к тубулину)
 ЗАПАСНЫЕ ВЕЩЕСТВА ЗЕЛЕНЫХ ВОДОРОСЛЕЙ 1. 2. 1. 3. 1 - хлоропласт зеленых водорослей (S- крахмал), 2 - структура крахмала, 3 - структура фруктана
ЗАПАСНЫЕ ВЕЩЕСТВА ЗЕЛЕНЫХ ВОДОРОСЛЕЙ 1. 2. 1. 3. 1 - хлоропласт зеленых водорослей (S- крахмал), 2 - структура крахмала, 3 - структура фруктана
 ЗАПАСНЫЕ ВЕЩЕСТВА ОХРОФИТОВЫХ ВОДОРОСЛЕЙ 1. 2. 1 - клетка охрофитовых водорослей (Ch- хлоропласт, CV- хризоламинарин), 2 - структура полисахарида хризоламинарина
ЗАПАСНЫЕ ВЕЩЕСТВА ОХРОФИТОВЫХ ВОДОРОСЛЕЙ 1. 2. 1 - клетка охрофитовых водорослей (Ch- хлоропласт, CV- хризоламинарин), 2 - структура полисахарида хризоламинарина
 ЗАПАСНЫЕ ВЕЩЕСТВА Euglenophyta 1. 2. 3. 1– запасной продукт в клетке Phacus, 2 - структура парамилона, производное глюкозы, вещество близкое по структуре к крахмалу 3 - гранулы парамилона
ЗАПАСНЫЕ ВЕЩЕСТВА Euglenophyta 1. 2. 3. 1– запасной продукт в клетке Phacus, 2 - структура парамилона, производное глюкозы, вещество близкое по структуре к крахмалу 3 - гранулы парамилона
 ЗАПАСНЫЕ ВЕЩЕСТВА Cyanophyta 1. 2. 1– клетка цианобактерий: CS- цианофициновый крахмал, CY- цианофициновые гранулы, L- капли масла, 2 - формула цианофицинового крахмала (резервный полисахарид, близкий к гликогену)
ЗАПАСНЫЕ ВЕЩЕСТВА Cyanophyta 1. 2. 1– клетка цианобактерий: CS- цианофициновый крахмал, CY- цианофициновые гранулы, L- капли масла, 2 - формула цианофицинового крахмала (резервный полисахарид, близкий к гликогену)
 ЗАПАСНЫЕ ВЕЩЕСТВА Rhodophyta 1. 2. 1 - клетка красных водорослей: C- хлоропласт, S- запасной продукт, 2 - структура багрянкового крахмала, близкий к амилопектину и гликогену
ЗАПАСНЫЕ ВЕЩЕСТВА Rhodophyta 1. 2. 1 - клетка красных водорослей: C- хлоропласт, S- запасной продукт, 2 - структура багрянкового крахмала, близкий к амилопектину и гликогену
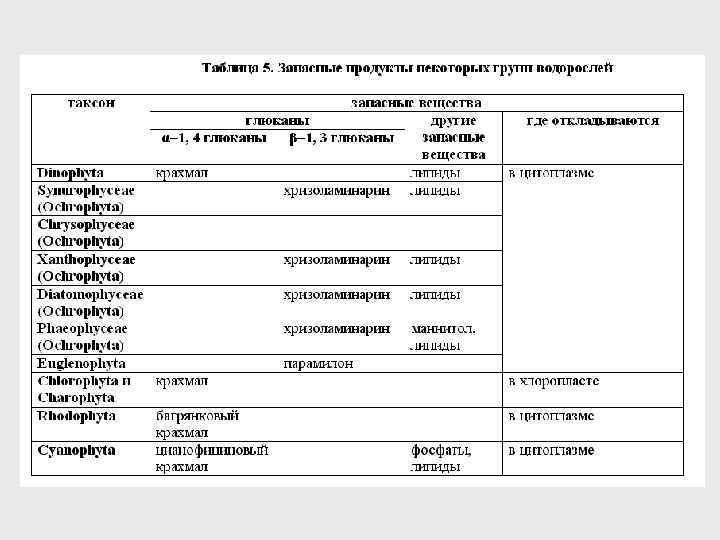
 ЗАПАСНЫЕ ВЕЩЕСТВА 1. 1– формулы глюканов
ЗАПАСНЫЕ ВЕЩЕСТВА 1. 1– формулы глюканов
 Пиреноид • Диференцированная область внутри хлоропласта, которая плотнее стромы и в которую могут заходить тилакоиды. • Пиреноид содержит фермент рибулезо 1, 5 -бисфосфаткарбоксилазуоксигеназу (Рубиско), который фиксирует двуокись углерода. • Рубиско у водорослей известна в двух формах – I и II
Пиреноид • Диференцированная область внутри хлоропласта, которая плотнее стромы и в которую могут заходить тилакоиды. • Пиреноид содержит фермент рибулезо 1, 5 -бисфосфаткарбоксилазуоксигеназу (Рубиско), который фиксирует двуокись углерода. • Рубиско у водорослей известна в двух формах – I и II
 • Форма I присутствует у некоторых бактерий, цианобактерий, всех водорослей, исключая динофитовых и высшие растения. • Состоит из восьми больших (54 к. Да кодируются хлоропластным геномом) и восьми малых субъединиц (14 к. Да, кодируется или в хлоропластном геноме или в ядре).
• Форма I присутствует у некоторых бактерий, цианобактерий, всех водорослей, исключая динофитовых и высшие растения. • Состоит из восьми больших (54 к. Да кодируются хлоропластным геномом) и восьми малых субъединиц (14 к. Да, кодируется или в хлоропластном геноме или в ядре).
; DIC (B, E); overlay of Rubisco fluorescence") Иммунолокализация Rubisco FITC окрашивание Rubisco (A, D); DIC (B, E); overlay of Rubisco fluorescence and DIC image (C, F). Rubisco is localized to a pyrenoid Chlamydomonas reinhardtii
Иммунолокализация Rubisco FITC окрашивание Rubisco (A, D); DIC (B, E); overlay of Rubisco fluorescence and DIC image (C, F). Rubisco is localized to a pyrenoid Chlamydomonas reinhardtii
 красные, желтозеленые, диатомовые эвгленовые, золотистые, зеленые эвгленовые, бурые, динофитовые, зеленые, празинофитовые Типы пиреноидов (Dodge, 1972)
красные, желтозеленые, диатомовые эвгленовые, золотистые, зеленые эвгленовые, бурые, динофитовые, зеленые, празинофитовые Типы пиреноидов (Dodge, 1972)
 хлореллы") Пиреноид (Рубиско) хлореллы
Пиреноид (Рубиско) хлореллы
 • Форма II встречается у некоторых эубактерий и большинства динофитовых водорослей • Состоит из двух больших субъединиц и кожируется ядерным геномом
• Форма II встречается у некоторых эубактерий и большинства динофитовых водорослей • Состоит из двух больших субъединиц и кожируется ядерным геномом

") Карбоксисомы цианобактерий (Рубиско)
Карбоксисомы цианобактерий (Рубиско)