

лекция 3- 1 В -ЛФ - копия.ppt
- Количество слайдов: 97
 Развитие В - клеток Функции В – клеток Гуморальный ответ
Развитие В - клеток Функции В – клеток Гуморальный ответ
 ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА 1954 - Bruce Glick, США Изучение функции Фабрициевой сумки (bursa Fabricius), лимфоидного органа в области клоаки у курицы Бурсэктомия у кур не приводила к видимым эффектам Бурсэктомированных цыплят использовали в экспериментах по получению антител к антигенам Salmonella Ни у одного из бурсэктомирован ных цыплят не было обнаружено антител против Salmonella Было установлено, что бурса – это орган, в котором развиваются антителообразующие клетки – поэтому их назвали B клетками у млекопитающих bursa Fabricius отсутствует
ОТКРЫТИЕ В - КЛЕТОЧНОГО ИММУНИТЕТА 1954 - Bruce Glick, США Изучение функции Фабрициевой сумки (bursa Fabricius), лимфоидного органа в области клоаки у курицы Бурсэктомия у кур не приводила к видимым эффектам Бурсэктомированных цыплят использовали в экспериментах по получению антител к антигенам Salmonella Ни у одного из бурсэктомирован ных цыплят не было обнаружено антител против Salmonella Было установлено, что бурса – это орган, в котором развиваются антителообразующие клетки – поэтому их назвали B клетками у млекопитающих bursa Fabricius отсутствует
 Происхождение В клеток и орган, в котором созревают В-клетки Перенос меченых клеток фетальной печени В крови есть зрелые В-клетки Нормальный костный мозг Зрелые В-клетки отсутствуют Дефектный костный мозг B клетки начинают развиваться в фетальной печени После рождения их развитие продолжается в костном мозге
Происхождение В клеток и орган, в котором созревают В-клетки Перенос меченых клеток фетальной печени В крови есть зрелые В-клетки Нормальный костный мозг Зрелые В-клетки отсутствуют Дефектный костный мозг B клетки начинают развиваться в фетальной печени После рождения их развитие продолжается в костном мозге
 Развитие В клеток в костном мозге B Регуляция созревания В рецепторов B Каждая В – клетка - моноспецифична B Уничтожение аутореактивных В - клеток B Выход полноценных, но еще незрелых В клеток на периферию (сначала в селезенку, затем - в лимфатические узлы) Костный мозг обеспечивает МИКРООКРУЖЕНИЕ ДЛЯ СОЗРЕВАНИЯ , ДИФФЕРЕНЦИРОВКИ И РАЗВИТИЯ В КЛЕТОК
Развитие В клеток в костном мозге B Регуляция созревания В рецепторов B Каждая В – клетка - моноспецифична B Уничтожение аутореактивных В - клеток B Выход полноценных, но еще незрелых В клеток на периферию (сначала в селезенку, затем - в лимфатические узлы) Костный мозг обеспечивает МИКРООКРУЖЕНИЕ ДЛЯ СОЗРЕВАНИЯ , ДИФФЕРЕНЦИРОВКИ И РАЗВИТИЯ В КЛЕТОК
 Схема развития В – клеток в костном мозге предшественники Э н д ос Т К О С Т Н О Й П Л А С Т И Н К и Клетки стромы Незрелые и зрелые В клетки Пре -B X X X макрофаг Центральный синус
Схема развития В – клеток в костном мозге предшественники Э н д ос Т К О С Т Н О Й П Л А С Т И Н К и Клетки стромы Незрелые и зрелые В клетки Пре -B X X X макрофаг Центральный синус
 Развитие В клеток в костном мозге • Выжившие клетки продолжают созревать и достигают центрального венозного синуса. • На всех этапах созревания В-клеток важную роль играет связь В-клеток с клетками стромы (микроокружение) и присутствие цитокинов – в частности, интерлейкина -7 (ИЛ-7).
Развитие В клеток в костном мозге • Выжившие клетки продолжают созревать и достигают центрального венозного синуса. • На всех этапах созревания В-клеток важную роль играет связь В-клеток с клетками стромы (микроокружение) и присутствие цитокинов – в частности, интерлейкина -7 (ИЛ-7).
 Развитие В клеток в костном мозге • Более 75% созревающих в костном мозге Вклеток не попадает в кровоток, а погибает путем апоптоза и поглощается костномозговыми макрофагами. • + селекция происходит при взаимодействии В - клеток и клеток стромы – остаются Вклетки с продуктивной перестройкой генов иммуноглобулинов (Ig).
Развитие В клеток в костном мозге • Более 75% созревающих в костном мозге Вклеток не попадает в кровоток, а погибает путем апоптоза и поглощается костномозговыми макрофагами. • + селекция происходит при взаимодействии В - клеток и клеток стромы – остаются Вклетки с продуктивной перестройкой генов иммуноглобулинов (Ig).
 Постулаты теории клональной селекции • Каждый В лимфоцит имеет рецептор уникальной специфичности. • Высокоаффинное (прочное) взаимодействие рецептора с антигеном приводит к активации лимфоцита. • Специфичность рецептора сохраняется в процессе пролиферации и дифференцировки лимфоцита. • Лимфоциты с рецепторами, специфичными к собственным антигенам, удаляются на ранней стадии дифференцировки.
Постулаты теории клональной селекции • Каждый В лимфоцит имеет рецептор уникальной специфичности. • Высокоаффинное (прочное) взаимодействие рецептора с антигеном приводит к активации лимфоцита. • Специфичность рецептора сохраняется в процессе пролиферации и дифференцировки лимфоцита. • Лимфоциты с рецепторами, специфичными к собственным антигенам, удаляются на ранней стадии дифференцировки.
 Созревающие В клетки Клетки стромы
Созревающие В клетки Клетки стромы
 B B Клетка стромы
B B Клетка стромы
 Стадии развития В клеток Стволовая клетка Ранняя про - В Поздняя про - В Большая пре - В периферическая Малая пре -В Незрелая В клетка На каждой стадии развития происходит реаранжировка генов тяжелых и легких цепей Ig, экспрессия поверхностных Ig, экспрессия адгезионных молекул и рецепторов к цитокинам
Стадии развития В клеток Стволовая клетка Ранняя про - В Поздняя про - В Большая пре - В периферическая Малая пре -В Незрелая В клетка На каждой стадии развития происходит реаранжировка генов тяжелых и легких цепей Ig, экспрессия поверхностных Ig, экспрессия адгезионных молекул и рецепторов к цитокинам
 Развитие В-лимфоцитов • Из костного мозга уже отобранные В-лимфоциты попадают с током крови в первичные фолликулы селезенки. • В селезенке происходит ряд этапов функционального «дозревания» В лимфоцитов, включая экспрессию МНС II класса на их поверхностной мембране. • Далее В лимфоциты мигрируют в лимфатические узлы – для встречи с комплементарным их рецептору антигеном. • До встречи с антигеном В лимфоцит называется «наивным» .
Развитие В-лимфоцитов • Из костного мозга уже отобранные В-лимфоциты попадают с током крови в первичные фолликулы селезенки. • В селезенке происходит ряд этапов функционального «дозревания» В лимфоцитов, включая экспрессию МНС II класса на их поверхностной мембране. • Далее В лимфоциты мигрируют в лимфатические узлы – для встречи с комплементарным их рецептору антигеном. • До встречи с антигеном В лимфоцит называется «наивным» .
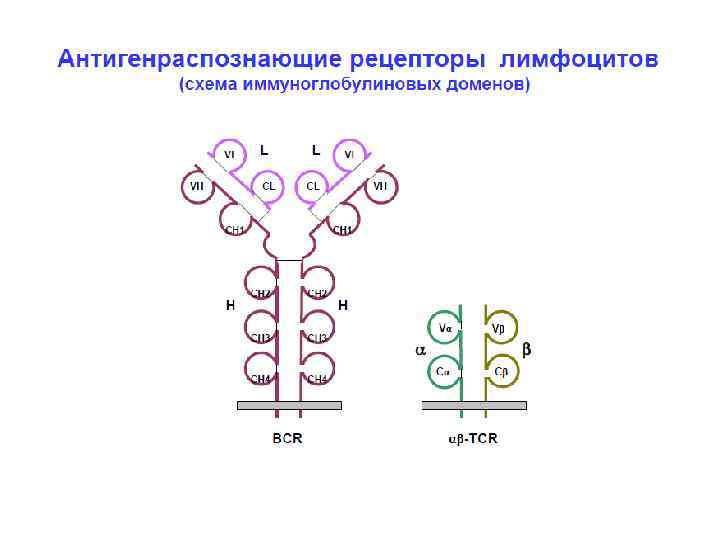
 и двух") гены Ig молекул • Каждая молекула Ig состоит из 2 тяжелых (Н) и двух легких (L) цепей, в каждой из этих цепей присутствуют константные (С) и вариабельные (V) области. • Вариабельные (V) и константные (С) области иммуноглобулиновых молекул кодируются отдельными генами.
гены Ig молекул • Каждая молекула Ig состоит из 2 тяжелых (Н) и двух легких (L) цепей, в каждой из этих цепей присутствуют константные (С) и вариабельные (V) области. • Вариабельные (V) и константные (С) области иммуноглобулиновых молекул кодируются отдельными генами.
, а") гены Ig молекул • Для вариабельных областей существует множество генов (V 1 -Vn), а для константой части молеулы Ig – один С-ген. • Легкие цепи иммуноглобулинов кодируются генными сегментами V и J. • Тяжелые цепи кодируются сегментами V и J, а также дополнительным сегментом (D).
гены Ig молекул • Для вариабельных областей существует множество генов (V 1 -Vn), а для константой части молеулы Ig – один С-ген. • Легкие цепи иммуноглобулинов кодируются генными сегментами V и J. • Тяжелые цепи кодируются сегментами V и J, а также дополнительным сегментом (D).
 Стадии дифференцировки определяются реаранжировкой генов Ig Стадии созревания конфигурация генов Ig. H Стволовая клетка Ранний про-В От Поздний про-В DH до JH От VH до DHJH Большой пре-В VHDHJH Пре-В клетка экспрес сирует рецептор Гены легких цепей Ig пока не реаранжируются
Стадии дифференцировки определяются реаранжировкой генов Ig Стадии созревания конфигурация генов Ig. H Стволовая клетка Ранний про-В От Поздний про-В DH до JH От VH до DHJH Большой пре-В VHDHJH Пре-В клетка экспрес сирует рецептор Гены легких цепей Ig пока не реаранжируются
 гены Ig молекул • Рекомбинации ограниченного числа генных сегментов V, D и J создают бесконечное число вариабельных доменов разной специфичности. • После антигенной стимуляции в генах легких и тяжелых цепей иммуноглобулинов происходят точечные соматические мутации (более тонкая «подгонка» АТ к АГ).
гены Ig молекул • Рекомбинации ограниченного числа генных сегментов V, D и J создают бесконечное число вариабельных доменов разной специфичности. • После антигенной стимуляции в генах легких и тяжелых цепей иммуноглобулинов происходят точечные соматические мутации (более тонкая «подгонка» АТ к АГ).
 5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 1. Многочисленность гаметных генов. Имеется большое число отдельных гаметных неперестроенных генов (V 1 - Vn), каждый из которых кодирует V –домен отдельной специфичности. 2. Соматический мутагенез. В онтогенезе В - клеток в результате мутаций гаметного V – гена в разных В - клеточных клонах возникают различные V – гены.
5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 1. Многочисленность гаметных генов. Имеется большое число отдельных гаметных неперестроенных генов (V 1 - Vn), каждый из которых кодирует V –домен отдельной специфичности. 2. Соматический мутагенез. В онтогенезе В - клеток в результате мутаций гаметного V – гена в разных В - клеточных клонах возникают различные V – гены.
 5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 3. Соматическая рекомбинация. В онтогенезе В-клеток происходит рекомбинация ряда генных сегментов (J 1 – Jn), соединяющихся с основной частью V – гена. В результате синтезируется белок, отдельные элементы которого кодируются разными генными сегментами.
5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 3. Соматическая рекомбинация. В онтогенезе В-клеток происходит рекомбинация ряда генных сегментов (J 1 – Jn), соединяющихся с основной частью V – гена. В результате синтезируется белок, отдельные элементы которого кодируются разными генными сегментами.
 5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 4. Генная конверсия. Отрезки ДНК, принадлежащие ряду псевдо - V – генов, могут копироваться в функциональном V – гене, меняя его исходную нуклеотидную последовательность.
5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 4. Генная конверсия. Отрезки ДНК, принадлежащие ряду псевдо - V – генов, могут копироваться в функциональном V – гене, меняя его исходную нуклеотидную последовательность.
 5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 5. Вставка добавочных нуклеотидов. При рекомбинации перед присоединением вырезанных V и J – сегментов ДНК возможно встраивание между ними добавочных нуклеотидов, кодирующих дополнительные аминокислотные остатки V-областей. Все эти 5 механизмов служат источником разнообразия антител у млекопитающих.
5 источников разнообразия Vобластей Н- и L -цепей молекул Ig 5. Вставка добавочных нуклеотидов. При рекомбинации перед присоединением вырезанных V и J – сегментов ДНК возможно встраивание между ними добавочных нуклеотидов, кодирующих дополнительные аминокислотные остатки V-областей. Все эти 5 механизмов служат источником разнообразия антител у млекопитающих.
 генов, кодирующих") Реаранжировка генов, кодирующих легкие цепи молекулы Ig • После завершения перестройки (реаранжировки) генов, кодирующих тяжелые цепи молекулы Ig, начинается перестройка генов легких цепей. Всего существует 2 типа легких цепей –либо каппа, либо лямбда. После этого на поверхности незрелого В лимфоцита появляется В - клеточный рецептор, состоящий из двух тяжелых цепей(H) и двух легких (L).
Реаранжировка генов, кодирующих легкие цепи молекулы Ig • После завершения перестройки (реаранжировки) генов, кодирующих тяжелые цепи молекулы Ig, начинается перестройка генов легких цепей. Всего существует 2 типа легких цепей –либо каппа, либо лямбда. После этого на поверхности незрелого В лимфоцита появляется В - клеточный рецептор, состоящий из двух тяжелых цепей(H) и двух легких (L).
 Общая схема перестройки генов тяжелых и легких цепей Ig в В - лимфоцитах Ранний про-B DH-JH на 1 хромо соме нет DH-JH на 2 хромо соме Поздний про- B Пре- B VH-DJH на 1 хромо соме k на 1 хромо соме да да нет VH-DJH на 2 хромосоме да нет да k на 2 хромо соме B да да Ig. M k Y B нет l на 1 хромо соме нет нет незрелый B l на 2 хромо соме нет да да Ig. M l Y B
Общая схема перестройки генов тяжелых и легких цепей Ig в В - лимфоцитах Ранний про-B DH-JH на 1 хромо соме нет DH-JH на 2 хромо соме Поздний про- B Пре- B VH-DJH на 1 хромо соме k на 1 хромо соме да да нет VH-DJH на 2 хромосоме да нет да k на 2 хромо соме B да да Ig. M k Y B нет l на 1 хромо соме нет нет незрелый B l на 2 хромо соме нет да да Ig. M l Y B
 Положительная и отрицательная селекция В лимфоцитов в костном мозге • + селекция происходит при взаимодействии В клеток и клеток стромы – остаются В-клетки с продуктивной перестройкой генов иммуноглобулинов (Ig), остальные – уничтожаются апоптозом. • - селекция – уничтожение аутореактивных Влимфоцитов может происходить и в костном мозге, и в селезенке – в органе, в который мигрирует большинство новообразованных В – клеток в период внутриутробного развития.
Положительная и отрицательная селекция В лимфоцитов в костном мозге • + селекция происходит при взаимодействии В клеток и клеток стромы – остаются В-клетки с продуктивной перестройкой генов иммуноглобулинов (Ig), остальные – уничтожаются апоптозом. • - селекция – уничтожение аутореактивных Влимфоцитов может происходить и в костном мозге, и в селезенке – в органе, в который мигрирует большинство новообразованных В – клеток в период внутриутробного развития.
 Выход зрелых В клеток на периферию • Из костного мозга выходят только те В – лимфоциты, у которых успешно произошла реаранжировка генов тяжелых и легких цепей Ig молекул, причем эти В –лимфоциты не активируются в ответ на аутоантигены –то есть не являются аутореактивными. Все остальные в клетки погибают в костном мозге путем апоптоза. • На поверхности отобранных В лимфоцитов экспрессируются Ig. M и Ig. D –Ig рецепторы, которые синтезируются с одной ДНК путем альтернативного сплайсинга.
Выход зрелых В клеток на периферию • Из костного мозга выходят только те В – лимфоциты, у которых успешно произошла реаранжировка генов тяжелых и легких цепей Ig молекул, причем эти В –лимфоциты не активируются в ответ на аутоантигены –то есть не являются аутореактивными. Все остальные в клетки погибают в костном мозге путем апоптоза. • На поверхности отобранных В лимфоцитов экспрессируются Ig. M и Ig. D –Ig рецепторы, которые синтезируются с одной ДНК путем альтернативного сплайсинга.
 В - клеточная аутотолерантность – выход зрелых В клеток из костного мозга B YY Незрелый В Ig. D Ig. M Малый пре –В не несет рецепторы Незрелый Влимфоцит не распознает ауто Аг B YY YY Малый пре-В YY Ig. D и Ig. M рецепторы Ig. D Ig. M Ig. D Зрелый В-лимфоцит выходит на периферию
В - клеточная аутотолерантность – выход зрелых В клеток из костного мозга B YY Незрелый В Ig. D Ig. M Малый пре –В не несет рецепторы Незрелый Влимфоцит не распознает ауто Аг B YY YY Малый пре-В YY Ig. D и Ig. M рецепторы Ig. D Ig. M Ig. D Зрелый В-лимфоцит выходит на периферию
 Функции вторичных лимфоидных органов • Захватывать патогены «на входе» в макроорганизм и тем самым ограничивать их распространение. • Способствовать контакту антигенпрезентирующих клеток и лимфоцитов. • Способствовать контакту Т и В лимфоцитов. • Обеспечивать ниши для дифференцировки эффекторных клеток.
Функции вторичных лимфоидных органов • Захватывать патогены «на входе» в макроорганизм и тем самым ограничивать их распространение. • Способствовать контакту антигенпрезентирующих клеток и лимфоцитов. • Способствовать контакту Т и В лимфоцитов. • Обеспечивать ниши для дифференцировки эффекторных клеток.
 Этапы дифференцировки В клеток на периферии • После того, как В лимфоцит встретится со «своим» антигеном в лимфоузле, он превратится в плазматическую клетку, синтезирующую антитела. • Плазматическая клетка –конечный этап дифференцировки В –лимфоцита.
Этапы дифференцировки В клеток на периферии • После того, как В лимфоцит встретится со «своим» антигеном в лимфоузле, он превратится в плазматическую клетку, синтезирующую антитела. • Плазматическая клетка –конечный этап дифференцировки В –лимфоцита.
 Фазы развития гуморального иммунного ответа
Фазы развития гуморального иммунного ответа
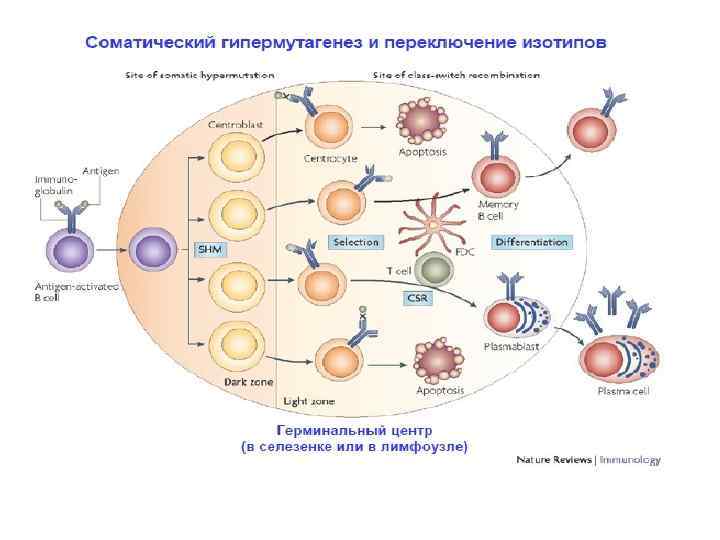
 События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном • После встречи наивного В лимфоцита с антигеном и контакта с Т-лимфоцитом-хелперов, В-лимфоцит входит в цикл пролиферации. Его потомки, несущие идентичные рецепторы, комплементарные к антигену, во вторичных фолликулах образуют зародышевые центры. • В зародышевых центрах интенсивная пролиферация сочетается с соматическими гипермутациями реаранжированных ранее V –генов и селекцией клеток, несущих высокоаффинные для данного антигена рецепторы.
События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном • После встречи наивного В лимфоцита с антигеном и контакта с Т-лимфоцитом-хелперов, В-лимфоцит входит в цикл пролиферации. Его потомки, несущие идентичные рецепторы, комплементарные к антигену, во вторичных фолликулах образуют зародышевые центры. • В зародышевых центрах интенсивная пролиферация сочетается с соматическими гипермутациями реаранжированных ранее V –генов и селекцией клеток, несущих высокоаффинные для данного антигена рецепторы.
 События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном • Гипермутации с частотой 2 -4% происходят в генах V –областей Ig. G и Ig. A, реже - Ig. M и характеризуются заменой одного нуклеотида (так называемые SNPs). • Биологический смысл: в результате таких гипермутаций повышается аффинность секретируемых антител – то есть сила их связи с антигеном. • В результате происходит более полная «подгонка» антител к антигену.
События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном • Гипермутации с частотой 2 -4% происходят в генах V –областей Ig. G и Ig. A, реже - Ig. M и характеризуются заменой одного нуклеотида (так называемые SNPs). • Биологический смысл: в результате таких гипермутаций повышается аффинность секретируемых антител – то есть сила их связи с антигеном. • В результате происходит более полная «подгонка» антител к антигену.
 События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном После соматических гипермутаций в V-областях генов иммуноглобулинов происходит отбор клеток, наиболее комплементарных к антигену- т. е. – антигензависимая селекция В-лимфоцитов. • После ее завершения для отобранных клеток есть 2 пути развития : • 1. Превращение в короткоживущие плазматические клетки, синтезирующие антитела. • 2. Превращение в долгоживущие В -клетки памяти, которые при повторном попадании антигена будут превращаться в плазматические клетки, сразу синтезирующие высокоаффинные антитела.
События в лимфоузле – посте встречи В лимфоцита с комплементарным его рецептору антигеном После соматических гипермутаций в V-областях генов иммуноглобулинов происходит отбор клеток, наиболее комплементарных к антигену- т. е. – антигензависимая селекция В-лимфоцитов. • После ее завершения для отобранных клеток есть 2 пути развития : • 1. Превращение в короткоживущие плазматические клетки, синтезирующие антитела. • 2. Превращение в долгоживущие В -клетки памяти, которые при повторном попадании антигена будут превращаться в плазматические клетки, сразу синтезирующие высокоаффинные антитела.
 в ЛУ и селезенке • B клетки экспрессируют поверхностные") Формирование зародышевых центров (центров размножения) в ЛУ и селезенке • B клетки экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК • Происходит апоптоз «несовершенных» В клеток : выживают только те В клетки, у которых рецепторы к данного антигену являются высокоаффинными, остальные –погибают путем апоптоза. • Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток
Формирование зародышевых центров (центров размножения) в ЛУ и селезенке • B клетки экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК • Происходит апоптоз «несовершенных» В клеток : выживают только те В клетки, у которых рецепторы к данного антигену являются высокоаффинными, остальные –погибают путем апоптоза. • Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток
 экспрессируют поверхностные Ig, прекращают делиться и получают") Микроанатомия зародышевого центра 2. B клетки (центроциты) экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК 4. Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток Primary Follicles become secondary follicles B Темная зона Светлая зона when germinal centres develop T ДК «отбирают» В лимфоциты 3. Апоптоз аутореактивных В клеток 1. B клетки (центробласты) пролиферируют, в них происходит соматическая гипермутация генов Ig созревание высокоаффинных Врецепторов
Микроанатомия зародышевого центра 2. B клетки (центроциты) экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК 4. Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток Primary Follicles become secondary follicles B Темная зона Светлая зона when germinal centres develop T ДК «отбирают» В лимфоциты 3. Апоптоз аутореактивных В клеток 1. B клетки (центробласты) пролиферируют, в них происходит соматическая гипермутация генов Ig созревание высокоаффинных Врецепторов
 Расположение, строение и функция лимфатических узлов • Лимфоузлы находятся на стыке лимфатических и кровеносных сосудов: по лимфе из тканей поступают клетки, готовые к презентации антигена, а также свободный антиген. • Из кровотока поступают клетки: Т лимфоциты для возможной встречи с антигеном, презентируемым дендритными клетками (ДК), и В клетки – для возможной встречи со свободным антигеном. • Т клетки выходят с лимфой и возвращаются в кровь через грудной проток (рециркуляция).
Расположение, строение и функция лимфатических узлов • Лимфоузлы находятся на стыке лимфатических и кровеносных сосудов: по лимфе из тканей поступают клетки, готовые к презентации антигена, а также свободный антиген. • Из кровотока поступают клетки: Т лимфоциты для возможной встречи с антигеном, презентируемым дендритными клетками (ДК), и В клетки – для возможной встречи со свободным антигеном. • Т клетки выходят с лимфой и возвращаются в кровь через грудной проток (рециркуляция).
 Развитие Т-хелперов различных типов под влиянием цитокиновых сигналов от дендритных клеток IL-6 IL-4 IFNg, IL-12 TGFb, IL-6 TGFb J J O’Shea, and W E Paul Science 2010; 327: 1098 -1102
Развитие Т-хелперов различных типов под влиянием цитокиновых сигналов от дендритных клеток IL-6 IL-4 IFNg, IL-12 TGFb, IL-6 TGFb J J O’Shea, and W E Paul Science 2010; 327: 1098 -1102
 и эфферентный (выносящий) лимфатические сосуды. • Снаружи капсула, под") Строение ЛУ • Афферентный (приносящий) и эфферентный (выносящий) лимфатические сосуды. • Снаружи капсула, под капсулой - кора – Вклеточная зона, в ней потом сформируются зародышевые центры –места интенсивного размножения клеток. • Паракортикальная зона –Т- клеточная зона.
Строение ЛУ • Афферентный (приносящий) и эфферентный (выносящий) лимфатические сосуды. • Снаружи капсула, под капсулой - кора – Вклеточная зона, в ней потом сформируются зародышевые центры –места интенсивного размножения клеток. • Паракортикальная зона –Т- клеточная зона.
 Строение ЛУ B клетки крови T клеточная зона B клеточн ая зона Эфферентная лимфа
Строение ЛУ B клетки крови T клеточная зона B клеточн ая зона Эфферентная лимфа
 Строение ЛУ • В клетки покидают сосуды и входят в ЛУ через ВЭВ (венулы с высоким эндотелием). • АГ попадает в ЛУ через афферентный приносящий лимфатический сосуд. • После встречи с антигеном В клетки быстро пролиферируют в зародышевых центрах. • Из зародышевого центра выходят В клетки, которые дифференцировались в плазматические клетки.
Строение ЛУ • В клетки покидают сосуды и входят в ЛУ через ВЭВ (венулы с высоким эндотелием). • АГ попадает в ЛУ через афферентный приносящий лимфатический сосуд. • После встречи с антигеном В клетки быстро пролиферируют в зародышевых центрах. • Из зародышевого центра выходят В клетки, которые дифференцировались в плазматические клетки.
 Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле В клетки покидают сосуды и входят в ЛУ через ВЭВ В клетки быстро пролиферируют Аг попадает в ЛУ через афферентный YY Y YY Y Зародышевый центр Интенсивная пролиферация Из зародышевого центра YY выходят В клетки, Y которые дифференци ровались в плазматичес кие клетки
Рециркулирующие В – клетки встречаются со «своим» антигеном в лимфоузле В клетки покидают сосуды и входят в ЛУ через ВЭВ В клетки быстро пролиферируют Аг попадает в ЛУ через афферентный YY Y YY Y Зародышевый центр Интенсивная пролиферация Из зародышевого центра YY выходят В клетки, Y которые дифференци ровались в плазматичес кие клетки
 YY Y YY Зрелый периферический В -лимфоцит B YY Распознал неауто-АГ на периферии B Y YY YY Дифференцировка В – клеток на периферии Ig - секретирующая плазматическая клетка
YY Y YY Зрелый периферический В -лимфоцит B YY Распознал неауто-АГ на периферии B Y YY YY Дифференцировка В – клеток на периферии Ig - секретирующая плазматическая клетка

 Плазматические клетки уровень рост экспрессии Ig B MHC II Высокий Да Ig секреция нет соматические переключение мутации изотипов да да нет да Зрелый В лимфоцит B низкий Плазматическая клетка нет да нет
Плазматические клетки уровень рост экспрессии Ig B MHC II Высокий Да Ig секреция нет соматические переключение мутации изотипов да да нет да Зрелый В лимфоцит B низкий Плазматическая клетка нет да нет
 СТРУКТУРА ДОМЕНОВ ИММУНОГЛОБУЛИНОВ Домены –компактно уложенные структуры, устойчивые к действию протеаз Fab Fc S S C L VL C H 3 S S CH 2 Домены тяжелых цепей , d, e, , или m CH 1 S VH S F(ab)2 Места отрыва пепсином Места отрыва папаином - 1 x (Fab)2 и 1 x Fc - 2 x Fab 1 x Fc С домены легких цепей k или l
СТРУКТУРА ДОМЕНОВ ИММУНОГЛОБУЛИНОВ Домены –компактно уложенные структуры, устойчивые к действию протеаз Fab Fc S S C L VL C H 3 S S CH 2 Домены тяжелых цепей , d, e, , или m CH 1 S VH S F(ab)2 Места отрыва пепсином Места отрыва папаином - 1 x (Fab)2 и 1 x Fc - 2 x Fab 1 x Fc С домены легких цепей k или l
 Fb Fb Fv Fv Fv Гибкость иммуноглобули новых молекул Fv CH 2 3 CH C H 3 CH 2 Fb Fv
Fb Fb Fv Fv Fv Гибкость иммуноглобули новых молекул Fv CH 2 3 CH C H 3 CH 2 Fb Fv
 фрагментов в молекулах Ig (Fab)2 фрагмент способен: • Распознавать") функции (Fab – antigen binding) фрагментов в молекулах Ig (Fab)2 фрагмент способен: • Распознавать антиген • Преципитировать антиген Блокировать активные сайты токсинов или патоген - ассоциированных молекул Блокировать взаимодействие между организмом –хозяином и чужеродными молекулами Но (Fab)2 не может активировать: • Воспаление и эффекторные функции клеток • Воспаление и эффекторные функции комплемента • Запускать процессинг антигенов
функции (Fab – antigen binding) фрагментов в молекулах Ig (Fab)2 фрагмент способен: • Распознавать антиген • Преципитировать антиген Блокировать активные сайты токсинов или патоген - ассоциированных молекул Блокировать взаимодействие между организмом –хозяином и чужеродными молекулами Но (Fab)2 не может активировать: • Воспаление и эффекторные функции клеток • Воспаление и эффекторные функции комплемента • Запускать процессинг антигенов
 Мономерный Ig. M На поверхности в клетки Ig. M существует только как мономер C m 4 мономерный Ig. M имеет очень низкую аффинность к антигену C m 3 Cm 2 Cm Показаны только константные области молекулы 1
Мономерный Ig. M На поверхности в клетки Ig. M существует только как мономер C m 4 мономерный Ig. M имеет очень низкую аффинность к антигену C m 3 Cm 2 Cm Показаны только константные области молекулы 1
 Cm") МУЛЬТИМЕРИЗАЦИЯ Ig. M Cm 2 1. 2 мономера Ig. M (показаны только Fc) Cm 2 C Cm 4 C Cm 3 s C C C CC ss C CC C C Cm 4 ss Cm 4 Cm 3 5. Цикл повторяется 2 Cm s. C C Cm 4 4. присоединяется J цепь другой мономерной молекулы Ig. M Cm 3 C m 2 6. Формируется пентамер. C дисульфидными мостиками C ss 3. J цепи соединяются m 4 Cm 3 формируют дисульфидные мостики с цистеинами каждого мономера 3 Cm 2. Цистеины в J цепях C m 2
МУЛЬТИМЕРИЗАЦИЯ Ig. M Cm 2 1. 2 мономера Ig. M (показаны только Fc) Cm 2 C Cm 4 C Cm 3 s C C C CC ss C CC C C Cm 4 ss Cm 4 Cm 3 5. Цикл повторяется 2 Cm s. C C Cm 4 4. присоединяется J цепь другой мономерной молекулы Ig. M Cm 3 C m 2 6. Формируется пентамер. C дисульфидными мостиками C ss 3. J цепи соединяются m 4 Cm 3 формируют дисульфидные мостики с цистеинами каждого мономера 3 Cm 2. Цистеины в J цепях C m 2
 Изменения конформации молекулы Ig. M, вызванные антигеном Форма «снежинки» Не фиксирует комплемент Форма «краба» Ig. M Конформация, вызванная связыванием с АГ Эффективно фиксирует комплемент
Изменения конформации молекулы Ig. M, вызванные антигеном Форма «снежинки» Не фиксирует комплемент Форма «краба» Ig. M Конформация, вызванная связыванием с АГ Эффективно фиксирует комплемент
 Ig. M m - Mю Тяжелая цепь: Время полужизни: 5 - 10 дней % от Ig сыворотки: Уровень в ыворотке 10 0. 25 - 3. 1 ( мг мл-1): Активация комплемента: ++++ классический путь Взаимодействие с клетками: фагоцитоз через C 3 b рецепторы эпителиальные клетки = полимерный Ig. R Перенос через плаценту : Аффинниость к Аг: нет мономерный Ig. M –– валентность = 2 пентамерный Ig. M - валентность = 10
Ig. M m - Mю Тяжелая цепь: Время полужизни: 5 - 10 дней % от Ig сыворотки: Уровень в ыворотке 10 0. 25 - 3. 1 ( мг мл-1): Активация комплемента: ++++ классический путь Взаимодействие с клетками: фагоцитоз через C 3 b рецепторы эпителиальные клетки = полимерный Ig. R Перенос через плаценту : Аффинниость к Аг: нет мономерный Ig. M –– валентность = 2 пентамерный Ig. M - валентность = 10
 Ig. D Тяжелая цепь: d - дельта Время полужизни: 2 - 8 дней % Ig в сыворотке: 0. 2 уровень (мг мл-1): 0. 03 - 0. 4 Активация комплемента: нет взаимодействие: T клетки через лектиноподобный Ig. D R Перенос через плаценту: нет Ig. D ко-экспрессируется вместе с Ig. M на В клеткк в результате альтернативного РНК - сплайсинга Ig. D –плазматические клетки обнаружены в назальной слизистой– однако функции этого класса иммуноглобулинов не ясны
Ig. D Тяжелая цепь: d - дельта Время полужизни: 2 - 8 дней % Ig в сыворотке: 0. 2 уровень (мг мл-1): 0. 03 - 0. 4 Активация комплемента: нет взаимодействие: T клетки через лектиноподобный Ig. D R Перенос через плаценту: нет Ig. D ко-экспрессируется вместе с Ig. M на В клеткк в результате альтернативного РНК - сплайсинга Ig. D –плазматические клетки обнаружены в назальной слизистой– однако функции этого класса иммуноглобулинов не ясны
 Ig. G Тяжелые цепи: g 1 g 2 g 3 g 4 - гамма 1 - 4 Время полужизни : Ig. G 1 Ig. G 3 21 - 24 дней 7 - 8 дней Ig. G 2 Ig. G 4 21 - 24 дней уровень (мг мл-1): Ig. G 1 Ig. G 3 5 - 12 0. 5 - 1 Ig. G 2 Ig. G 4 2 -6 0. 2 - 1 % от Ig сыворотки: Ig. G 1 Ig. G 3 45 - 53 3 -6 Ig. G 2 Ig. G 4 11 - 15 1 -4 Активация комплемента: Ig. G 1 Ig. G 3 Взаимодействие с клетками: ++++ Ig. G 2 Ig. G 4 + нет все субклассы через Ig. G рецепторы на МФ Перенос через плаценту: Ig. G 1 Ig. G 3 ++ ++ Ig. G 2 Ig. G 4 + ++
Ig. G Тяжелые цепи: g 1 g 2 g 3 g 4 - гамма 1 - 4 Время полужизни : Ig. G 1 Ig. G 3 21 - 24 дней 7 - 8 дней Ig. G 2 Ig. G 4 21 - 24 дней уровень (мг мл-1): Ig. G 1 Ig. G 3 5 - 12 0. 5 - 1 Ig. G 2 Ig. G 4 2 -6 0. 2 - 1 % от Ig сыворотки: Ig. G 1 Ig. G 3 45 - 53 3 -6 Ig. G 2 Ig. G 4 11 - 15 1 -4 Активация комплемента: Ig. G 1 Ig. G 3 Взаимодействие с клетками: ++++ Ig. G 2 Ig. G 4 + нет все субклассы через Ig. G рецепторы на МФ Перенос через плаценту: Ig. G 1 Ig. G 3 ++ ++ Ig. G 2 Ig. G 4 + ++
 Ig. A - димеризация и секреция Ig. A –основной изотип антител, секретируемых слизистыми В сыворотке существует в виде мономера, но обычно - как димер, связанный J цепью , который образуется сходным образом с Ig. M пентамерами S S S C J C C ss C C C S S S Существует 2 субкласса Ig. A 1 в основном присутствует в сыворотке и продуцируется костномозговыми B клетками Ig. A 2 в основном присутствует в секретах, молозиве и молоке и продуцируется B клетками, локализованными в слизистых
Ig. A - димеризация и секреция Ig. A –основной изотип антител, секретируемых слизистыми В сыворотке существует в виде мономера, но обычно - как димер, связанный J цепью , который образуется сходным образом с Ig. M пентамерами S S S C J C C ss C C C S S S Существует 2 субкласса Ig. A 1 в основном присутствует в сыворотке и продуцируется костномозговыми B клетками Ig. A 2 в основном присутствует в секретах, молозиве и молоке и продуцируется B клетками, локализованными в слизистых
 секреторный Ig. A и трансцитоз S S S S C J C C C C ss C S S S B C J C C ss C S S Эпителиаль ная клетка p. Ig. R и Ig. A интер нализируются S S B клетки подслизистого слоя продуцируют димерные Ig. A S S CC CJC Cs C s S S S S C J C C ss C Ig. A и p. Ig. R транспортирую тся к апикальной поверхности в везикулах S S комплекс p. Ig. R разрушается, освобождается Ig. A с частью p. Ig. R - секреторным компонентом полимерные Ig- рецепторы экспрессируются на базолатеральной поверхности эпителиальных клеток, захватывают Ig. A , продуцируемые в слизистой
секреторный Ig. A и трансцитоз S S S S C J C C C C ss C S S S B C J C C ss C S S Эпителиаль ная клетка p. Ig. R и Ig. A интер нализируются S S B клетки подслизистого слоя продуцируют димерные Ig. A S S CC CJC Cs C s S S S S C J C C ss C Ig. A и p. Ig. R транспортирую тся к апикальной поверхности в везикулах S S комплекс p. Ig. R разрушается, освобождается Ig. A с частью p. Ig. R - секреторным компонентом полимерные Ig- рецепторы экспрессируются на базолатеральной поверхности эпителиальных клеток, захватывают Ig. A , продуцируемые в слизистой
 Ig. A Тяжелые цепи: a 1 или a 2 - альфа 1 or 2 Время полужизни: Ig. A 1 5 - 7 дней Ig. A 2 4 - 6 дней Уровень в сыворотке Ig. A 1 1. 4 - 4. 2 (мг мл-1): Ig. A 2 0. 2 - 0. 5 % от Ig сыворотки : Ig. A 1 11 - 14 Ig. A 2 1 - 4 Активация комплемента: Ig. A 1 – альтернативный и лектиновый путь Ig. A 2 - нет Взаимодействие с клетками: эпителий - через p. Ig. R фагоциты -через Ig. AR перенос через плаценту : нет
Ig. A Тяжелые цепи: a 1 или a 2 - альфа 1 or 2 Время полужизни: Ig. A 1 5 - 7 дней Ig. A 2 4 - 6 дней Уровень в сыворотке Ig. A 1 1. 4 - 4. 2 (мг мл-1): Ig. A 2 0. 2 - 0. 5 % от Ig сыворотки : Ig. A 1 11 - 14 Ig. A 2 1 - 4 Активация комплемента: Ig. A 1 – альтернативный и лектиновый путь Ig. A 2 - нет Взаимодействие с клетками: эпителий - через p. Ig. R фагоциты -через Ig. AR перенос через плаценту : нет
 Ig. E Тяжелые цепи: e - эпсилон Время полужизни: 1 - 5 дней Уровень в сыворотк 0. 0001 - 0. 0002 % от Ig сыворотки : 0. 004 Активация комплемента : нет Взаимодействие : через высокоаффинные Ig. E R тучных клеток, эозинофилов, базофилов, клеток Лангерганса через низкоаффинные Ig. E R В лф , моноцитов перенос через плаценту : нет Ig. E появился позже в эволюции в связи с его ролью в защите от паразитарных инфекций Большинство Ig. E – молекул абсорбируется на Ig. E рецепторах эффекторных клеток Ig. E также тесно связан с аллергией
Ig. E Тяжелые цепи: e - эпсилон Время полужизни: 1 - 5 дней Уровень в сыворотк 0. 0001 - 0. 0002 % от Ig сыворотки : 0. 004 Активация комплемента : нет Взаимодействие : через высокоаффинные Ig. E R тучных клеток, эозинофилов, базофилов, клеток Лангерганса через низкоаффинные Ig. E R В лф , моноцитов перенос через плаценту : нет Ig. E появился позже в эволюции в связи с его ролью в защите от паразитарных инфекций Большинство Ig. E – молекул абсорбируется на Ig. E рецепторах эффекторных клеток Ig. E также тесно связан с аллергией
 Различия в аминокислотной последовательности молекул антител • 1. Изотипические различия – у всех изотипов одинаковая аминокислотная последовательность в антигенсвязывающем центре (Ig. M, Ig. D, Ig. G, Ig. A, Ig. E). • Аллотипические различия – разные последовательности аминокислот в константных областях тяжелых цепей у разных особей 1 вида. • Идиотипы – различия в аминокислотных последовательностях в антигенсвязывающих центрах молекул Ig (в гипервариабельных участках).
Различия в аминокислотной последовательности молекул антител • 1. Изотипические различия – у всех изотипов одинаковая аминокислотная последовательность в антигенсвязывающем центре (Ig. M, Ig. D, Ig. G, Ig. A, Ig. E). • Аллотипические различия – разные последовательности аминокислот в константных областях тяжелых цепей у разных особей 1 вида. • Идиотипы – различия в аминокислотных последовательностях в антигенсвязывающих центрах молекул Ig (в гипервариабельных участках).
 в активации В лимфоцитов при ответе на Т- зависимые антигены") Роль антигенпрезентирующих клеток (АПК) в активации В лимфоцитов при ответе на Т- зависимые антигены (большинство белковых антигенов) • Т- зависимые антигены одновременно распознаются и Т- и В-лимфоцитами, при этом Влимфоциты распознают эпитопы антигенов, а Тлимфоциты –белки-носители антигена. • Информацию об антигене Т- и В-лимфоцитам передают антигенпредставляющие клетки (АПК), которые осуществляют процессинг антигена и предоставляют его в иммуногенной форме лимфоцитам в виде комплекса: антигенный пептид+ молекула MHC. • В зародышевых центрах ЛУ роль АПК играют дендритные клетки.
Роль антигенпрезентирующих клеток (АПК) в активации В лимфоцитов при ответе на Т- зависимые антигены (большинство белковых антигенов) • Т- зависимые антигены одновременно распознаются и Т- и В-лимфоцитами, при этом Влимфоциты распознают эпитопы антигенов, а Тлимфоциты –белки-носители антигена. • Информацию об антигене Т- и В-лимфоцитам передают антигенпредставляющие клетки (АПК), которые осуществляют процессинг антигена и предоставляют его в иммуногенной форме лимфоцитам в виде комплекса: антигенный пептид+ молекула MHC. • В зародышевых центрах ЛУ роль АПК играют дендритные клетки.
 Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • В лимфоциты презентируют антиген Т клеткам, и от Т-клеток получают сигналы «помощи» и запуска пролиферации и дифференцировки. • В ходе иммунного ответа помогают В лимфоцитам только те Т-клетки, которым уже был представлен комплекс АГ-пептид +MHC антигенпрезентирующей клеткой.
Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • В лимфоциты презентируют антиген Т клеткам, и от Т-клеток получают сигналы «помощи» и запуска пролиферации и дифференцировки. • В ходе иммунного ответа помогают В лимфоцитам только те Т-клетки, которым уже был представлен комплекс АГ-пептид +MHC антигенпрезентирующей клеткой.
 Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • Мембранносвязанный Ig. R(рецептор) на В лимфоците взаимодействует с АГ, расположенным в иккосомах, доставленных В –лимфоциту дендритными клетками. • Путем микропиноцитоза АГ попадает внутрь В лимфоцита, где он деградируется, его фрагменты упаковываются в молекулы MHC II и выносятся на клеточную поверхность В лимфоцита.
Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • Мембранносвязанный Ig. R(рецептор) на В лимфоците взаимодействует с АГ, расположенным в иккосомах, доставленных В –лимфоциту дендритными клетками. • Путем микропиноцитоза АГ попадает внутрь В лимфоцита, где он деградируется, его фрагменты упаковываются в молекулы MHC II и выносятся на клеточную поверхность В лимфоцита.
 Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • Комплекс MHC II +АГ пептид распознается Тклеточным рецепторм (TCR), а молекула MHC II – корецептором (CD 4), после этого распознавания в Т клетку идет первый сигнал активации. • Молекулы CD 80/CD 86 на поверхности В клетки связываются с молекулой CD 28 на поверхности Т -клетки, в Т-клетку идет второй сигнал активации. • Молекула CD 40 L на поверхности Т-клетки связывается с молекулой CD 40 на поверхности В клетки – в В клетку идет сильнейший сигнал к активации.
Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • Комплекс MHC II +АГ пептид распознается Тклеточным рецепторм (TCR), а молекула MHC II – корецептором (CD 4), после этого распознавания в Т клетку идет первый сигнал активации. • Молекулы CD 80/CD 86 на поверхности В клетки связываются с молекулой CD 28 на поверхности Т -клетки, в Т-клетку идет второй сигнал активации. • Молекула CD 40 L на поверхности Т-клетки связывается с молекулой CD 40 на поверхности В клетки – в В клетку идет сильнейший сигнал к активации.
 Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • Белок CD 40 на поверхности В клетки принадлежит к семейству рецепторов фактора некроза опухолей, взаимодействие молекул CD 40 на В лимфоците с CD 40 L на Тлимфоците запускает вход В лимфоцита в клеточный цикл. • Этот сигнал – один из самых важных для активации В клеток, дальнейшего синтеза и переключения классов иммуноглобулинов. • Дефект в гене молекулы CD 40 L приводит к развитию гипер - Ig. M –синдрома – отсутствию переключения классов иммуноглобулинов.
Взаимодействие Т и В лимфоцитов при ответе на Т- зависимые антигены • Белок CD 40 на поверхности В клетки принадлежит к семейству рецепторов фактора некроза опухолей, взаимодействие молекул CD 40 на В лимфоците с CD 40 L на Тлимфоците запускает вход В лимфоцита в клеточный цикл. • Этот сигнал – один из самых важных для активации В клеток, дальнейшего синтеза и переключения классов иммуноглобулинов. • Дефект в гене молекулы CD 40 L приводит к развитию гипер - Ig. M –синдрома – отсутствию переключения классов иммуноглобулинов.
 Взаимодействия Т и В лимфоцитов, приводящие к активации клеток • В процессе взаимодействия Т- и Влимфоцитов участвуют также следующие группы комплементарных молекул: • CD 2 (Т клетка) – LFA-3(В клетка) • LFA-1 (Т клетка) – ICAM-1 (В клетка)
Взаимодействия Т и В лимфоцитов, приводящие к активации клеток • В процессе взаимодействия Т- и Влимфоцитов участвуют также следующие группы комплементарных молекул: • CD 2 (Т клетка) – LFA-3(В клетка) • LFA-1 (Т клетка) – ICAM-1 (В клетка)
 Цитокиновая активация В клеток • Активированные Т клетки секретируют цитокины, активирующие В клетки: • IL-2 –индуктор активации Т и В клеток. • IL-4 –действует на ранней стадии активации и пролиферации В клеток. • IL-6 –активатор В - клеточной дифференцировки. • TNF и - рост В –клеток.
Цитокиновая активация В клеток • Активированные Т клетки секретируют цитокины, активирующие В клетки: • IL-2 –индуктор активации Т и В клеток. • IL-4 –действует на ранней стадии активации и пролиферации В клеток. • IL-6 –активатор В - клеточной дифференцировки. • TNF и - рост В –клеток.
 Ответ В клеток на Т-независимые антигены • При ответе на Т-независимые антигены В лимфоцитам не нужна помощь Т-лимфоцитов. • Т - независимые антигены –липополисахариды бактерий, бактериальные углеводы –декстран, леван, белки бактерий – флагеллин, эндотоксин и др. – все они представляют собой молекулы с часто повторяющимися структурными участками, в высоких концентрациях они являются митогенами –то есть способны активировать многие клоны В –клеток (поликлональная активация), в низких концентрациях – только специфические клоны В клеток.
Ответ В клеток на Т-независимые антигены • При ответе на Т-независимые антигены В лимфоцитам не нужна помощь Т-лимфоцитов. • Т - независимые антигены –липополисахариды бактерий, бактериальные углеводы –декстран, леван, белки бактерий – флагеллин, эндотоксин и др. – все они представляют собой молекулы с часто повторяющимися структурными участками, в высоких концентрациях они являются митогенами –то есть способны активировать многие клоны В –клеток (поликлональная активация), в низких концентрациях – только специфические клоны В клеток.
 LPS связывается с CD 14, LPSBP и TLR 4 B") T независимые антигены (липополисахариды) LPS связывается с CD 14, LPSBP и TLR 4 B B B Y Y Y 6 различных B клеток, для активации которых требуется 6 различных АГ Высокая доза LPS вызывает поликлональную активацию всех 6 типов Вклеток, независимо от их специфичности. Поэтому Т-независимые антигены называются МИТОГЕНАМИ Y YY YY Y Y Y YY YY YY
T независимые антигены (липополисахариды) LPS связывается с CD 14, LPSBP и TLR 4 B B B Y Y Y 6 различных B клеток, для активации которых требуется 6 различных АГ Высокая доза LPS вызывает поликлональную активацию всех 6 типов Вклеток, независимо от их специфичности. Поэтому Т-независимые антигены называются МИТОГЕНАМИ Y YY YY Y Y Y YY YY YY
 Ответ В клеток на Т-независимые антигены • В ответе В клеток на Т-независимые антигены отсутствует взаимодействие Т- и В-клеток, нет взаимодействия молекул CD 40 -CD 40 L, нет сильного сигнала к активации В клеток. • Первичный ответ – слабый, слабое формирование клеточной памяти. Такой ответ в основном осуществляет В 1 популяция клеток, синтезирующих только Ig. M –антитела (нет переключения изотипов).
Ответ В клеток на Т-независимые антигены • В ответе В клеток на Т-независимые антигены отсутствует взаимодействие Т- и В-клеток, нет взаимодействия молекул CD 40 -CD 40 L, нет сильного сигнала к активации В клеток. • Первичный ответ – слабый, слабое формирование клеточной памяти. Такой ответ в основном осуществляет В 1 популяция клеток, синтезирующих только Ig. M –антитела (нет переключения изотипов).
 B-1 клетки Распознают эпитопы АГ с повторяющимися структурами - фосфотидилхолин, липополисахариды и др. CD 5 Натуральные антитела Y B Y Y Y Y • присутствуют с рождения Y Y Y Не являются частью адаптивного иммунитета: • нет иммунологической памяти • нет более высокой эффективности ответа при повторном введении АГ Ig. M Могут производить Ig без помощи T клеток
B-1 клетки Распознают эпитопы АГ с повторяющимися структурами - фосфотидилхолин, липополисахариды и др. CD 5 Натуральные антитела Y B Y Y Y Y • присутствуют с рождения Y Y Y Не являются частью адаптивного иммунитета: • нет иммунологической памяти • нет более высокой эффективности ответа при повторном введении АГ Ig. M Могут производить Ig без помощи T клеток
 Захват иккосом /антигенов В клеткой В ИККОСОМАХ НАХОДЯТСЯ анти- B Ig Y Y Y РАЗЛИЧНЫЕ АНТИГЕНЫ B Поверхностный Ig захватывает АГ CD 40 Перекрестное связывание рецепторов с антигенами активирует В клетку Активированные В клетки экспрессируют CD 40
Захват иккосом /антигенов В клеткой В ИККОСОМАХ НАХОДЯТСЯ анти- B Ig Y Y Y РАЗЛИЧНЫЕ АНТИГЕНЫ B Поверхностный Ig захватывает АГ CD 40 Перекрестное связывание рецепторов с антигенами активирует В клетку Активированные В клетки экспрессируют CD 40
 Антигены интернализируются В - клеткой B 1. Связывание антигена максимизирует захват антигенов 2. Связывание и интернализация через Ig вызывает экспрессию CD 40 3. Антиген подвергается процессингу 4. Пептидные фрагменты АГ упаковыва ются в молекулы MHC внутри клетки. Комплексы MHC/пептид экспресси руются на поверхность клетки B
Антигены интернализируются В - клеткой B 1. Связывание антигена максимизирует захват антигенов 2. Связывание и интернализация через Ig вызывает экспрессию CD 40 3. Антиген подвергается процессингу 4. Пептидные фрагменты АГ упаковыва ются в молекулы MHC внутри клетки. Комплексы MHC/пептид экспресси руются на поверхность клетки B
 T клетки помогают B клеткам сигнал 2 - T клеточная помощь B YYY сигнал 1 антиген и рецептор к антигену Th Th 1. T клеточный рецептор 2. Ко-рецептор (CD 4) 3. CD 40 лиганд
T клетки помогают B клеткам сигнал 2 - T клеточная помощь B YYY сигнал 1 антиген и рецептор к антигену Th Th 1. T клеточный рецептор 2. Ко-рецептор (CD 4) 3. CD 40 лиганд
 T клеточная помощь - сигнал 2 цитокины Th B IL-4 IL-5 IL-6 IFN- TGF- цитокины Y YY сигнал 1 B клетки погибают путем апоптоза, однако сигналы1 и 2 повышают экспрессию Bcl-XL на В клетках, а Bcl-XL предотвращают апоптоз сигналы 1 и 2 позволяют В – клеткам выживать T клетки регулируют выживаемость B клеток и, т. о. , контролируют их клональную селекцию
T клеточная помощь - сигнал 2 цитокины Th B IL-4 IL-5 IL-6 IFN- TGF- цитокины Y YY сигнал 1 B клетки погибают путем апоптоза, однако сигналы1 и 2 повышают экспрессию Bcl-XL на В клетках, а Bcl-XL предотвращают апоптоз сигналы 1 и 2 позволяют В – клеткам выживать T клетки регулируют выживаемость B клеток и, т. о. , контролируют их клональную селекцию
 Первичный ответ на АГ • При первичном попадании антигена в организм развивается первичный гуморальный ответ, он протекает в 4 фазы: • Лаг – фаза –антитела в сыворотке крови не обнаруживаются. • Лог - фаза – титр антител нарастает логарифмически. • Фаза плато –стабилизация титра антител. • Фаза затухания –катаболизирование или выведение из организма антител.
Первичный ответ на АГ • При первичном попадании антигена в организм развивается первичный гуморальный ответ, он протекает в 4 фазы: • Лаг – фаза –антитела в сыворотке крови не обнаруживаются. • Лог - фаза – титр антител нарастает логарифмически. • Фаза плато –стабилизация титра антител. • Фаза затухания –катаболизирование или выведение из организма антител.
 Вторичный ответ на АГ • При повторном попадании антигена в организм развивается вторичный иммунный ответ, который характеризуется укороченной лаг - фазой и более продолжительными фазами плато и затухания. • Уровень антител во время фазы плато обычно более, чем в 10 раз превышает количество антител при первичном ответе.
Вторичный ответ на АГ • При повторном попадании антигена в организм развивается вторичный иммунный ответ, который характеризуется укороченной лаг - фазой и более продолжительными фазами плато и затухания. • Уровень антител во время фазы плато обычно более, чем в 10 раз превышает количество антител при первичном ответе.
 Сравнительная характеристика первичного и вторичного иммунного ответа
Сравнительная характеристика первичного и вторичного иммунного ответа
 Продукция Ig. M- и Ig. G-антител при первичном и вторичном иммунном ответе Всего Уровень антител, усл. ед/мл Вторичный ответ Первичный ответ Всего Время, сут 1 -я иммунизация 2 -я иммунизация
Продукция Ig. M- и Ig. G-антител при первичном и вторичном иммунном ответе Всего Уровень антител, усл. ед/мл Вторичный ответ Первичный ответ Всего Время, сут 1 -я иммунизация 2 -я иммунизация
 Временные характеристики первичного ответа • При первичном иммунном ответе первыми синтезируются антитела класса М (начало –на 3 -4 сутки, пик –на 7 сутки, затухание – на 14 сутки после введения АГ) • Позже происходит синтез антител класса G (начало – на 6 сутки, пик – на 14 сутки, затухание - на 21 сутки).
Временные характеристики первичного ответа • При первичном иммунном ответе первыми синтезируются антитела класса М (начало –на 3 -4 сутки, пик –на 7 сутки, затухание – на 14 сутки после введения АГ) • Позже происходит синтез антител класса G (начало – на 6 сутки, пик – на 14 сутки, затухание - на 21 сутки).
 Временные характеристики вторичного ответа • При вторичном иммунном ответе синтезируются антитела класса G уже на 23 сутки, пик их уровня достигается на 7 сутки, фаза плато – более длительная. • Антитела класса М при вторичном ответе присутствуют лишь в небольших количествах.
Временные характеристики вторичного ответа • При вторичном иммунном ответе синтезируются антитела класса G уже на 23 сутки, пик их уровня достигается на 7 сутки, фаза плато – более длительная. • Антитела класса М при вторичном ответе присутствуют лишь в небольших количествах.
 Аффинность антител Аффинитет –это степень сродства антител и антигенов, сила связывания антителом антигенов. • Аффинность Ig. M антител на протяжении всего ответа остается постоянной. • Афинность Ig. G антител при вторичных ответах возрастает (феномен созревания аффинности), причем при использовании более низких доз антигена продуцируются более высокоаффинные Ig. G –антитела.
Аффинность антител Аффинитет –это степень сродства антител и антигенов, сила связывания антителом антигенов. • Аффинность Ig. M антител на протяжении всего ответа остается постоянной. • Афинность Ig. G антител при вторичных ответах возрастает (феномен созревания аффинности), причем при использовании более низких доз антигена продуцируются более высокоаффинные Ig. G –антитела.
 Контроль аффинитета связи АГ с АТ Все эти 5 В клеточ ных рецепторов специфичны для , но аффинитет связи различается вследствие сомати ческой гипермута ции генов Ig в заро дышевом центре B B B Только эта клетка, которая обладает наиболее высокой аффинностью к антигену, может экспрессировать CD 40. Только эта клетка может получить сигнал 2. Только эта клетка избежит апоптоза – т. е. подвергнется клональной селекции, остальные – погибнут путем апоптоза.
Контроль аффинитета связи АГ с АТ Все эти 5 В клеточ ных рецепторов специфичны для , но аффинитет связи различается вследствие сомати ческой гипермута ции генов Ig в заро дышевом центре B B B Только эта клетка, которая обладает наиболее высокой аффинностью к антигену, может экспрессировать CD 40. Только эта клетка может получить сигнал 2. Только эта клетка избежит апоптоза – т. е. подвергнется клональной селекции, остальные – погибнут путем апоптоза.
 Роль цитокинов в переключении изотипов Ig • Т- клеточные цитокины, освобождаемые в непосредственной близости к В-клеткам, определяют новый изотип молекулы иммуноглобулина. • Например, IL-4 –активирует процесс переключения изотипа с Ig. G 1 на Ig. E, одновременно ингибируя переключение и синтез Ig. G 2. • Трансформирующий ростовый фактор бета (TGF ) –переключение на Ig. A и Ig. G 2, а интерферон – гамма (IFN- ) – на Ig. G 2.
Роль цитокинов в переключении изотипов Ig • Т- клеточные цитокины, освобождаемые в непосредственной близости к В-клеткам, определяют новый изотип молекулы иммуноглобулина. • Например, IL-4 –активирует процесс переключения изотипа с Ig. G 1 на Ig. E, одновременно ингибируя переключение и синтез Ig. G 2. • Трансформирующий ростовый фактор бета (TGF ) –переключение на Ig. A и Ig. G 2, а интерферон – гамма (IFN- ) – на Ig. G 2.
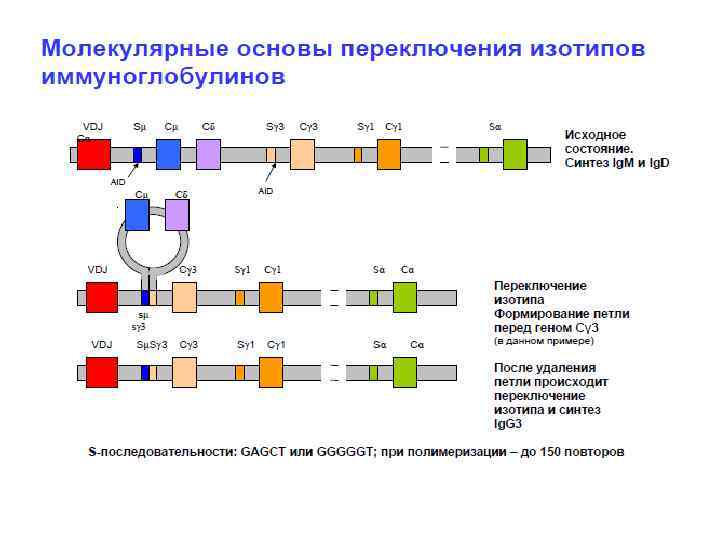
 AID-зависимый механизм переключения изотипов антител VDJ Sμ Cμ Cμ Cδ Sγ 3 Cγ 3 Sγ 1 Cγ 1 Sα Cα Исходное состояние. Синтез Ig. M и Ig. D Cδ activation-induced cytidin deaminase (AID) превращает дезоксицитидин в дезоксиуридин Cγ 3 VDJ Sγ 1 Cγ 1 Sα Cα Переключение изотипа. Формирование петли перед геном Сγ 3 (в sμ sγ 3 VDJ SμSγ 3 Cγ 3 Sγ 1 Cγ 1 Sα Cα данном примере) После удаления петли происходит переключение изотипа и синтез Ig. G 3
AID-зависимый механизм переключения изотипов антител VDJ Sμ Cμ Cμ Cδ Sγ 3 Cγ 3 Sγ 1 Cγ 1 Sα Cα Исходное состояние. Синтез Ig. M и Ig. D Cδ activation-induced cytidin deaminase (AID) превращает дезоксицитидин в дезоксиуридин Cγ 3 VDJ Sγ 1 Cγ 1 Sα Cα Переключение изотипа. Формирование петли перед геном Сγ 3 (в sμ sγ 3 VDJ SμSγ 3 Cγ 3 Sγ 1 Cγ 1 Sα Cα данном примере) После удаления петли происходит переключение изотипа и синтез Ig. G 3
 Роль цитокинов в дифференцировке и переключении изотипов антител у мышей Плазматические клетки В-лимфоциты Антиген TGFβ IFNγ m. Ig. A/G 2 b IL-2, IL-4, IL-5 IL-4 m. Ig. G 2 a/3 m. Ig. M m. Ig. G 1/E IL-21 участвует на всех этапах дифференцировки и переключения! IL-2, IL-4, IL-5 Ig. M Пролиферация Ig. G 1, Ig. E Ig. G 2 a, Ig. G 3 Ig. A, Ig. G 2 b Переключение изотипов. Дифференцировка
Роль цитокинов в дифференцировке и переключении изотипов антител у мышей Плазматические клетки В-лимфоциты Антиген TGFβ IFNγ m. Ig. A/G 2 b IL-2, IL-4, IL-5 IL-4 m. Ig. G 2 a/3 m. Ig. M m. Ig. G 1/E IL-21 участвует на всех этапах дифференцировки и переключения! IL-2, IL-4, IL-5 Ig. M Пролиферация Ig. G 1, Ig. E Ig. G 2 a, Ig. G 3 Ig. A, Ig. G 2 b Переключение изотипов. Дифференцировка
 Последовательность переключения классов иммуноглобулинов Cm Cd C 3 C 1 C 2 C 4 Cd Ce Cd C 2 Sg 3 C 3 Cm Sg 1 Cm C 1 VDJ C 3 VDJ C 1 Ig. G 3 продукция Переключение с Ig. M Ig. A 1 продукция Переключение с Ig. G 3 Ig. A 1 продукция Переключение с Ig. M Переключение классов иммуноглобулинов в одной клетке
Последовательность переключения классов иммуноглобулинов Cm Cd C 3 C 1 C 2 C 4 Cd Ce Cd C 2 Sg 3 C 3 Cm Sg 1 Cm C 1 VDJ C 3 VDJ C 1 Ig. G 3 продукция Переключение с Ig. M Ig. A 1 продукция Переключение с Ig. G 3 Ig. A 1 продукция Переключение с Ig. M Переключение классов иммуноглобулинов в одной клетке
 Роль T клеточных цитокинов в T клеточной помощи сигнал 2 цитокины IL-4 IL-5 IL-6 IFN- TGF- Th B Y YY сигнал 1 B B B B ПК B B B Пролиферация и дифференцировка Ig. M IL 4 IL-5 IFN- TGF- B B B Ig. G 3 Ig. G 1 Ig. G 2 b Ig. G 2 a Ig. E + Ig. A + + +
Роль T клеточных цитокинов в T клеточной помощи сигнал 2 цитокины IL-4 IL-5 IL-6 IFN- TGF- Th B Y YY сигнал 1 B B B B ПК B B B Пролиферация и дифференцировка Ig. M IL 4 IL-5 IFN- TGF- B B B Ig. G 3 Ig. G 1 Ig. G 2 b Ig. G 2 a Ig. E + Ig. A + + +
 Регуляция специфичности иммунного ответа 1. T клетки могут помогать только тем В-клетками, которые презентируют им антиген. 2. B клетки выполняют функцию презентации антигена, то есть являются профессиональными антигенпредставляющими клетками. 3. B клетки наиболее эффективно поглощают те антигены, которые обладают наибольшим сродством к их В – клеточным рецепторам. 4. Т и В клетки помогают другу, что усиливает иммунный ответ на один и тот же антиген. После окончания ответа остаются Т и В – клетки памяти
Регуляция специфичности иммунного ответа 1. T клетки могут помогать только тем В-клетками, которые презентируют им антиген. 2. B клетки выполняют функцию презентации антигена, то есть являются профессиональными антигенпредставляющими клетками. 3. B клетки наиболее эффективно поглощают те антигены, которые обладают наибольшим сродством к их В – клеточным рецепторам. 4. Т и В клетки помогают другу, что усиливает иммунный ответ на один и тот же антиген. После окончания ответа остаются Т и В – клетки памяти
 И Т клеткой (зеленый цвет)") Связь между В клеткой (красный цвет) И Т клеткой (зеленый цвет)
Связь между В клеткой (красный цвет) И Т клеткой (зеленый цвет)
 , окруженная В – клетками, цитоскелет окрашен зеленым цветом") T клетка (в центре) , окруженная В – клетками, цитоскелет окрашен зеленым цветом
T клетка (в центре) , окруженная В – клетками, цитоскелет окрашен зеленым цветом
 Активация В клеток • Спустя несколько суток после проникновения АГ в организм, Влимфоциты, активированные Т-клетками посредством связывания CD 40 c CD 40 L, мигрируют в первичные фолликулы, где имеется густая сеть фолликулярных ДК. • В этих фолликулах клетки быстро делятся, в них происходит соматическая гипермутация генов Ig и созревание Врецепторов.
Активация В клеток • Спустя несколько суток после проникновения АГ в организм, Влимфоциты, активированные Т-клетками посредством связывания CD 40 c CD 40 L, мигрируют в первичные фолликулы, где имеется густая сеть фолликулярных ДК. • В этих фолликулах клетки быстро делятся, в них происходит соматическая гипермутация генов Ig и созревание Врецепторов.
 в ЛУ и селезенке • B клетки экспрессируют поверхностные") Формирование зародышевых центров (центров размножения) в ЛУ и селезенке • B клетки экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК • Происходит апоптоз «несовершенных» В клеток : выживают только те В клетки, у которых рецепторы к данного антигену являются высокоаффинными, остальные –погибают путем апоптоза. • Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток
Формирование зародышевых центров (центров размножения) в ЛУ и селезенке • B клетки экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК • Происходит апоптоз «несовершенных» В клеток : выживают только те В клетки, у которых рецепторы к данного антигену являются высокоаффинными, остальные –погибают путем апоптоза. • Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток
 экспрессируют поверхностные Ig, прекращают делиться и получают") Микроанатомия зародышевого центра 2. B клетки (центроциты) экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК 4. Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток Primary Follicles become secondary follicles B Темная зона Светлая зона when germinal centres develop T ДК «отбирают» В лимфоциты 3. Апоптоз аутореактивных В клеток 1. B клетки (центробласты) пролиферируют, в них происходит соматическая гипермутация генов Ig созревание высокоаффинных Врецепторов
Микроанатомия зародышевого центра 2. B клетки (центроциты) экспрессируют поверхностные Ig, прекращают делиться и получают костимулирующие сигналы от T клеток и ДК 4. Отобранные клетки покидают ЛУ в виде клеток памяти или плазматических клеток Primary Follicles become secondary follicles B Темная зона Светлая зона when germinal centres develop T ДК «отбирают» В лимфоциты 3. Апоптоз аутореактивных В клеток 1. B клетки (центробласты) пролиферируют, в них происходит соматическая гипермутация генов Ig созревание высокоаффинных Врецепторов
 Свойства антител • Нейтрализация • Опсонизация • Активация системы комплемента
Свойства антител • Нейтрализация • Опсонизация • Активация системы комплемента
 Эффекторные механизмы защиты против внеклеточных патогенов НЕЙТРАЛИЗАЦИЯ ТОКСИН ` ` ` Y БАКТЕРИЯ ` ` Блокировка адгезии к клеткам хозяина Предотвращение токсичности ` Y Блокировка освобождения токсина инвазии Нейтрализующие антитела ` Y
Эффекторные механизмы защиты против внеклеточных патогенов НЕЙТРАЛИЗАЦИЯ ТОКСИН ` ` ` Y БАКТЕРИЯ ` ` Блокировка адгезии к клеткам хозяина Предотвращение токсичности ` Y Блокировка освобождения токсина инвазии Нейтрализующие антитела ` Y
 Эффекторные механизмы защиты против внеклеточных патогенов ОПСОНИЗАЦИЯ Внеклеточная бактерия + АТ ОПСОНИЗАЦИЯ Сязывание с Fc рецептором фагоцитоз
Эффекторные механизмы защиты против внеклеточных патогенов ОПСОНИЗАЦИЯ Внеклеточная бактерия + АТ ОПСОНИЗАЦИЯ Сязывание с Fc рецептором фагоцитоз
 Эффекторные механизмы защиты против внеклеточных патогенов Активация комплемента Бактерия в плазме лизис + АТ+ комплемент опсонизация Комплемент и Fc рецепторное связывание фагоцитоз
Эффекторные механизмы защиты против внеклеточных патогенов Активация комплемента Бактерия в плазме лизис + АТ+ комплемент опсонизация Комплемент и Fc рецепторное связывание фагоцитоз