
Молекулярные основы биорегуляции Сигнальные системы клетки. Первичные посредники





 - это процесс,")

.")
 1971 г. – Нобелевская премия по")





 1. Высокое сродство рецептора к сигнальным молекулам.")



")
")
")
 1. «Узкое» место СТ - область")














")

 – это химические соединения или физические факторы (кванты света), которые")

 Щитовидная железа Производное")
 Преимущественно в печени Белок (8, 4 к. Да,")

 1. Биосинтез ПМ в секреторных клетках 2.")

 — биологически")
 Эффекты гормонов проявляются в крайне малых концентрациях — в диапазоне от")



: ФСГ, лютеонизирующий")






 и тироксин (Т 4) – это гормоны на")




 - это биологически активные")



,")
")

 Медиатор по ЭПС переносится в")

, который")
 содержит токсин, представляющий собой")


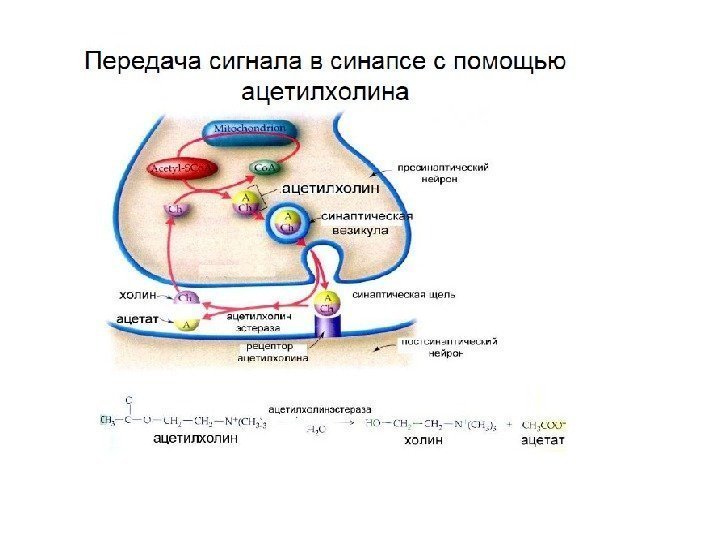

 Синтез из холина и ацетил-Ко.")

 запускает выброс Ац. Х в синапти-ческую щель, после чего")


 – антагонист")
 (1 -2 нейронов, мотивационно-эмоциональная сфера) Синтез из тирозина в ЦНС (черная")
, фермент – тирозин-гидроксилаза 2. L-DOPA путем")








, фермент – тирозин-гидроксилаза 2. L-DOPA путем")



 – 1 -2 нейронов, мотивационно-эмоциональная сфера Синтез из 5 - окситриптофана")
;")

 Одержимые пляской Святой Антоний")
 в коре, гиппокампе, стриатуме, гипоталамусе,")

 мосте, продолговатом мозге, гиппокампе")
 Белковый синтез (11 а. к. ) в спинномозговых ганглиях, стенках полых")
 Действуют в концентрациях 10 -9 (в 1000")

















 (гиперкортицизм ) • ИЗБЫТОК СИГНАЛА •")
 и мимикрия сигнала (ТГИХ, β-карболины)")
 – алкоголизм, наркомания - связаны с глубокими нарушениями медиаторного обмена")





 и мезокортикальный (")
 — ключевой")
, центральный медиатор -")



 - хроническое прогрессирующее заболевание головного мозга с дегенерацией нейронов черной субстанции")


: — нарушение")
. Белок альфа-синуклеин играет роль в синаптическом везикулярном транспорте и")
 – это заболевания, связанные с нарушнием механизмов нативной укладки клеточных")
, бокового амиотрофического")





 Семейство АТФаз Р-типа Семейство АТФаз F- и V-типа Суперсемейство АТФаз")



 имеют ряд")


 „ Диабет не болезнь, а образ")


 — системное аутоиммунное заболевание,")







")

4.prezent.integralynye_reakcii_kletki_pri_povreghdenii.pptx
- Размер: 41.3 Мб
- Автор:
- Количество слайдов: 208
Описание презентации Молекулярные основы биорегуляции Сигнальные системы клетки. Первичные посредники по слайдам
Молекулярные основы биорегуляции Сигнальные системы клетки. Первичные посредники
Фундаментальные свойства живых систем — Открытые — Сложные — Иерархические — Неравновесные — Автономные — Саморегулируемые — Самовоспроизводящиеся
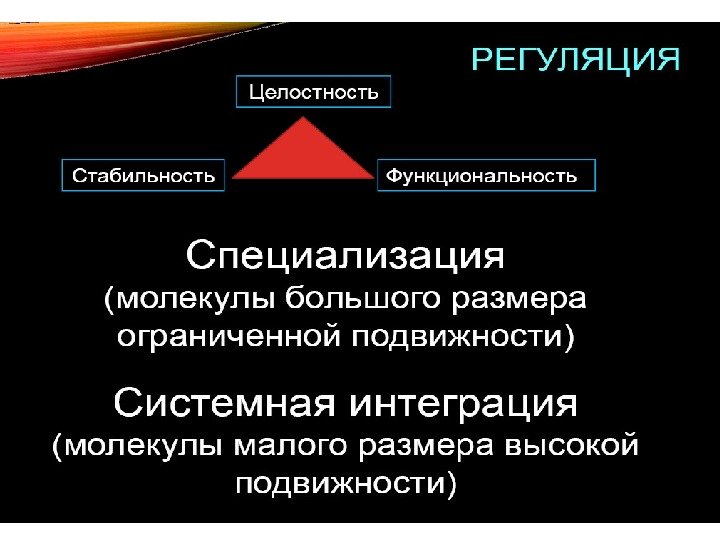
Для чего нужна регуляция в биосистемах? Целостность Стабильность Функциональность
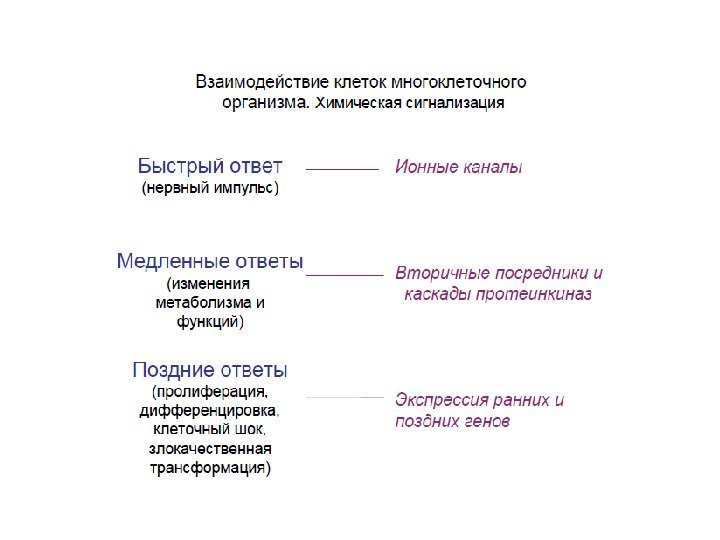
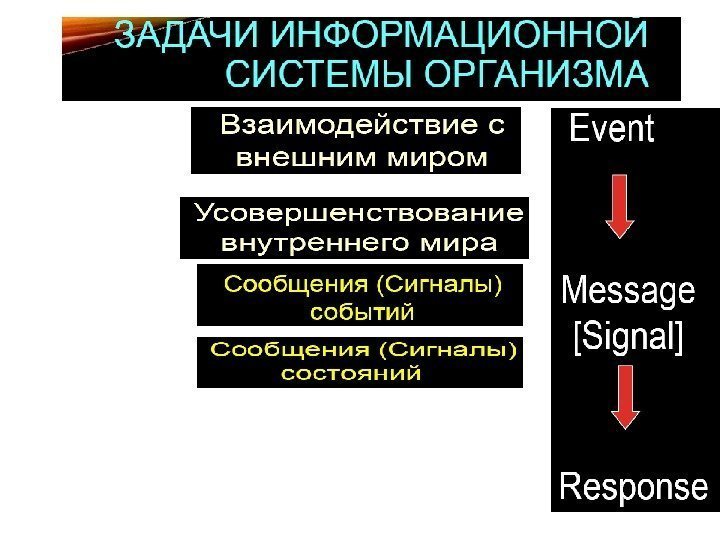
1. Фундаментальное свойство живых систем – получать сигналы извне и отвечать на них. 2. Сигнал — это информация , которая распознается и принимается специфическими рецепторами , затем преобразуется в клеточный ответ, всегда включающий химический процесс. 3. Превращение сигнала в химические реакции называется передачей (трансдукцией) сигнала
1. Передача сигнала (сигнальная трансдукция, signal transduction ) — это процесс, при помощи которого клетка превращает один тип сигнала в другой, что приводит к включению адаптивных программ. Сигнальная трансдукция — процесс преобразования сигнала на пути от клеточной мембраны к ядру. 2. К компонентам сигнальной трансдукции относятся: — мембранные рецепторы (молекулы, воспринимающие сигнал на поверхности клетки) — молекулы-эффекторы (адапторы, исполнители), которые с помощью белок-белковых взаимодействий передают сигнал к ядру — факторы транскрипции — белковые молекулы, влияющие на экспрессию определенных генов, что приводит к возникновению клеточного ответа. 3. Рецептор и связанные с ним эффекторы формируют сигнальный путь (signalling pathway). Сигнальные пути могут интегрировать сигналы, образуя сигнальные сети (signalling network ). 4. Сигнальные пути образуют сигнальные каскады, в которых происходит усиление (амплификация) первичного сигнала. Основные принципы клеточной сигнализации
Уровни передачи информации в клетке
Функционирование системы внутриклеточной сигнальной трансдукции (формирование ответа клетки на внешний сигнал).
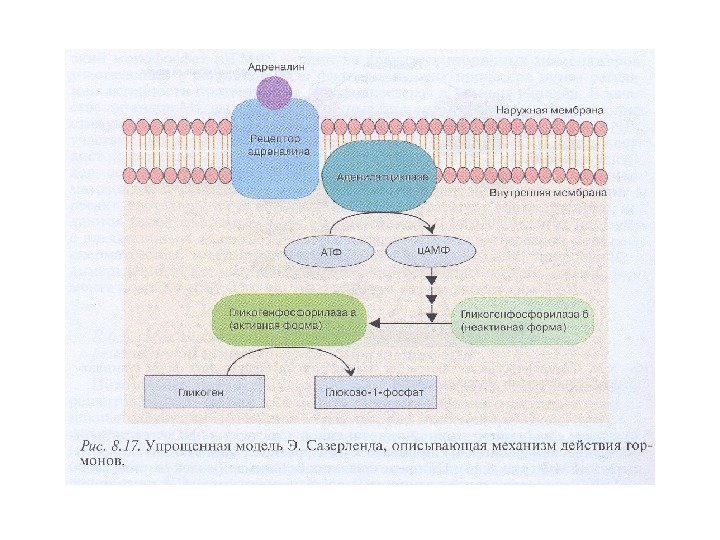
Эрл Уилбор Сазерленд (1915 -1974 ) 1971 г. – Нобелевская премия по физиологии и медицине «за открытие, связанное с выяснением механизма действия гормонов» Он выделил циклический АМФ (ц. АМФ) и показал его связь с функциями многих гормонов, «открыл один из фундаментальных принципов процессов жизнедеятельности» . «ц. АМФ влияет на все – от памяти до кончиков пальцев»
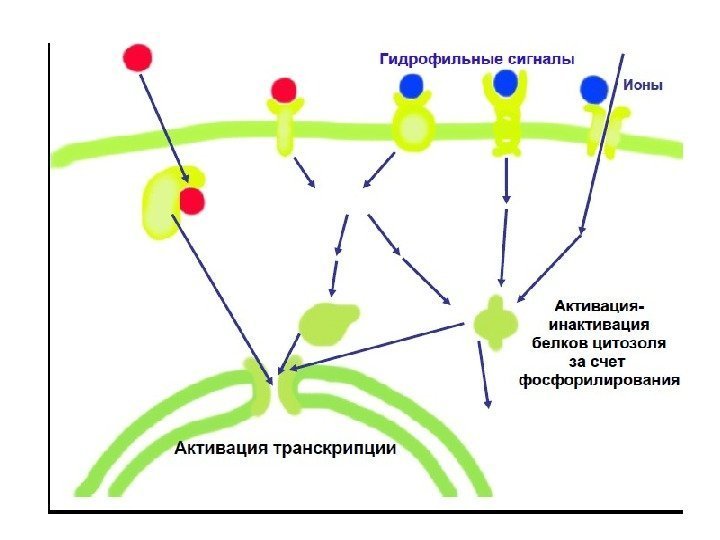
Регулирующие сигналы вызывают различные ответы у прокариот и эукариот В ответ на регулирующие сигналы: Клетка прокариот отвечает включением или выключением транскрипции Эукариотическая клетка многоклеточного организма отвечает: — включением/выключением или усилением/ослаблением транскрипции; — активацией/инактивацией белков ранее синтезированных клеткой
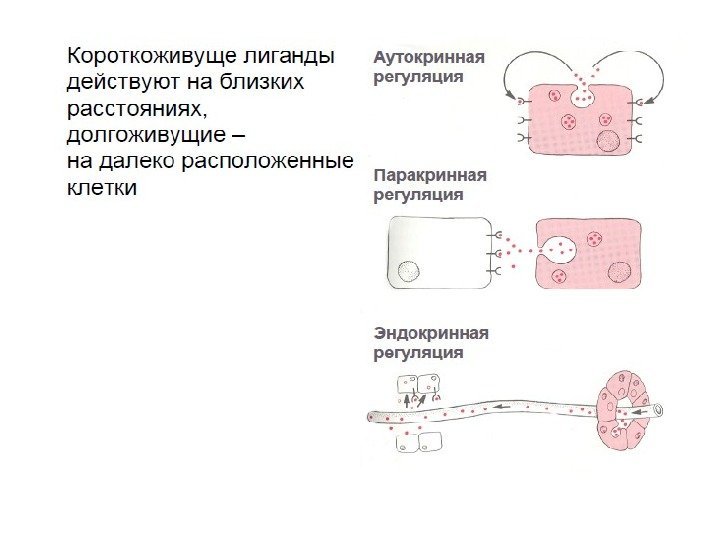
Типы межклеточной сигнализации
Общие свойства передачи сигнала 1. Все клетки имеют специфические и высоко чувствительные сигнал-передающие механизмы , которые сохраняются в процессе эволюции. 2. Множество различных сигналов действуют через специфические белковые рецепторы в клеточной мембране. 3. Рецепторы связывают сигнальную молекулу, усиливают и интегрируют сигнал, передают его в клетку. Десенсибилизация рецептора уменьшает или выключает сигнал. 4. Многоклеточные организмы имеют 6 основных механизмов передачи сигнала.
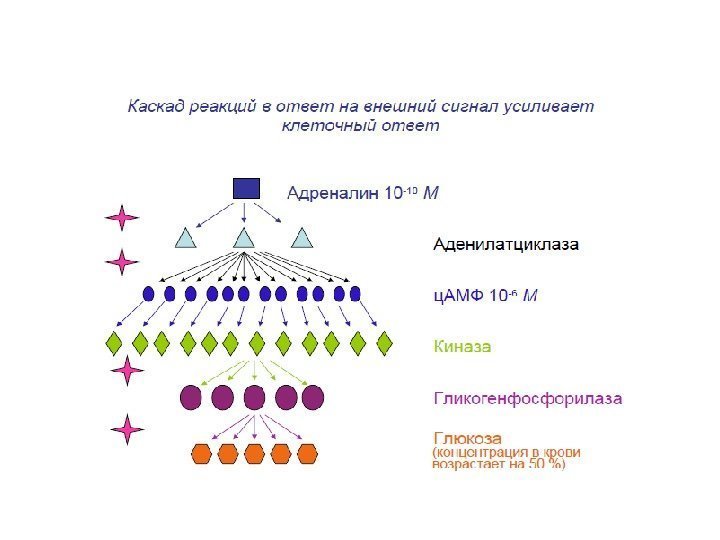
Три фактора обеспечивают высокую чувствительность передатчика (трансдуктора) 1. Высокое сродство рецептора к сигнальным молекулам. Рецептор обнаруживает пикомолярные концентрации вещества (Кd = 10 -8 M). 2. Кооперативность лиганд-рецепторного взаимодействия. Приводит к сильной активации рецептора при малых изменениях концентрации лиганда. 3. Усиление сигнала ферментативными каскадами (амплификация). Такие сигналы могут в течение мсек. увеличивать усиление на несколько порядков.
Характерные особенности клеточной сигнализации 1. Специфичность – определяется комплементарностью между сигнальной молекулой и рецептором. 2. Способность к усилению сигнала — определяется: а)высоким сродством сигнальной молекулы к рецептору; б)усилением за счет включения каскада ферментов. 3. Десенситизация – потеря чувствительности при постоянном воздействии сигнала до тех пор, пока сигнал не снизится ниже порогового уровня. 4. Интеграция – способность клетки получать и объединять множество сигналов от различных первичных посредников (гормонов, медиаторов и др. ).
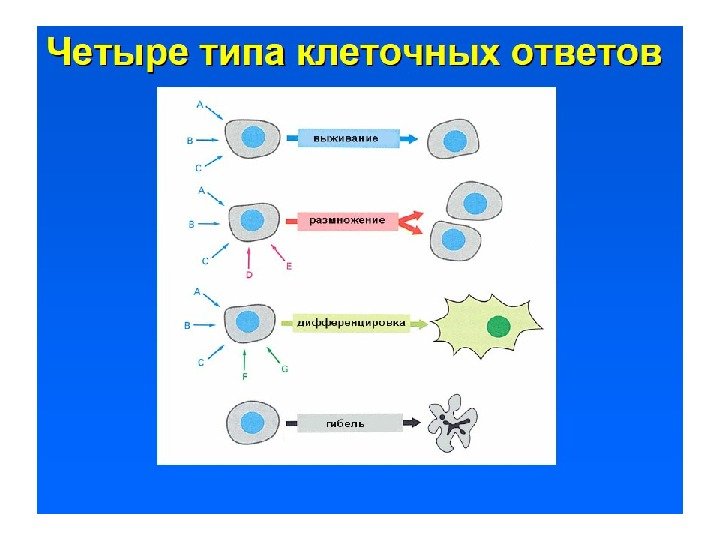
Четыре отличительных свойства систем передачи сигнала
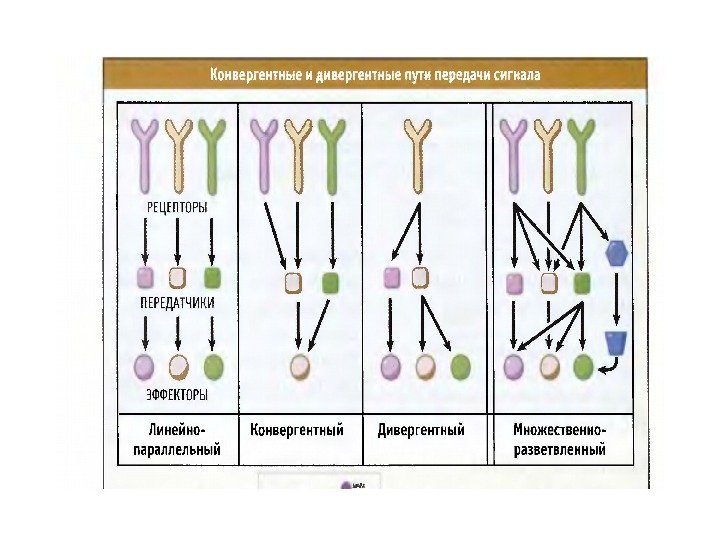
В сигнальных сетях сигналы сортируются и интегрируются • Процессы передачи сигнала являются многоступенчатыми и могут дивергировать и конвергировать • Дивергенция обеспечивает множественность ответа на один сигнал • Конвергенция позволяет сигналам интегрироваться и выполняет координирующую функцию
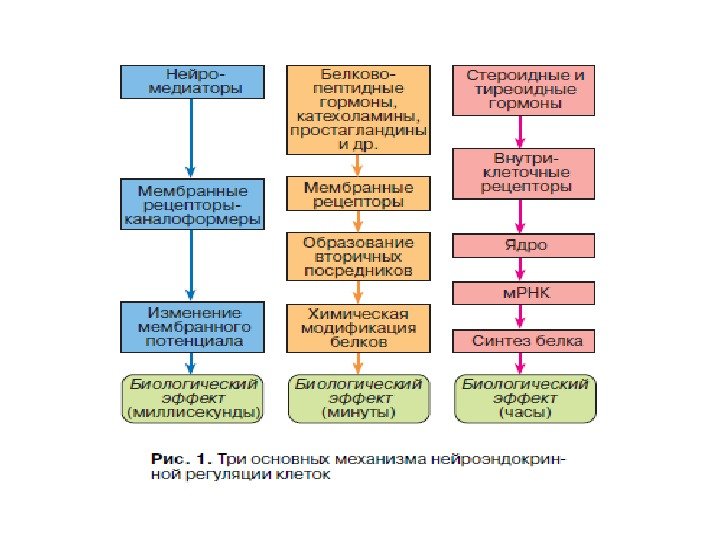
Структура холинергического рецептора никотинового типа, формирующего ионный канал (быстрый ответ)
Система проведения гормонального сигнала путем образования вторичных посредников и фосфорилирования белков (медленный ответ)
Механизм действия гормонов на процессы транскрипции и синтеза белка (поздний ответ)
«Песочные часы» путей сигнальной трансдукции (СТ) 1. «Узкое» место СТ — область передачи сигнала от рецептора к факторам транскрипции (ФТ) ( критический узел ), т. к. количество эффекторов здесь ограничено. 2. В критических узлах идет формирование различных клеточных ответов на сигналы. 3. Ограниченное число эффекторов компенсируется большим количест-вом их комбинаций на адапторных белках , которые определяют специфичность СТ. 4. Средняя часть сети СТ ( «горлышко» песочных часов) отвечает за мо-дулирование и разнообразие клеточных ответов и используется многими типами рецепторов. 5. В сети СТ имеются позитивные и негативные петли обратной связи.
Система сигнальной трансдукции
Шесть основных сигнальных механизмов 1. Рецепторы, сопряженные с G-белками , которые активируют ферменты, образующие вторичные посредники. 2. Рецепторные тирозинпротеинкиназы (РТП) являются ферментами и при активации запускают киназные каскады. 3. Рецепторные гуанилатциклазы с цитоплазматическим ферментативным доменом образуют вторичный посредник – ц. ГМФ , который активирует протеинкиназу, фосфорилирующую клеточные белки. 4. Управляемые (регулируемые) ионные каналы плазматической мембраны, состояние которых регулируется внешними сигналами или мембранным потенциалом. 5. Адгезионные рецепторы , которые взаимодействуют с внеклеточным матриксом и передают цитоскелету сигналы к миграции клетки или сцеплению с матриксом. 6. Ядерные рецепторы , которые при связывании с лигандом изменяют экспрессию генов.
Шесть основных механизмов передачи сигнала
Каркасные структуры увеличивают эффективность систем передачи сигналов и улучшает их пространственную организацию • Каркасные структуры организуют группы сигнальных белков и могут обеспечивать специфичность процесса передачи сигналов, запрещая функционирование компонентов, имеющих несколько партнеров. • Каркасные структуры селективно связывают определенные сигнальные белки и увеличивают локальную концентрацию сигнальных белков. • Каркасные структуры размещают пути передачи сигнала на местах их функционирования.
Каркасный белок INAD организует белки, передающие зрительные сигналы на фоторецепторные кдетки дрозофилы
Важное условие передачи сигнала – белок-белковые взаимодействия за счет независимых модульных доменов адапторных белков 1. Адапторные белки – не обладают ферментативной активностью, связывают сигнальные молекулы и располагают их в пространстве для передачи сигнала. 2. Характерные особенности адапторных белков: — состоят исключительно из модульных доменов или представляют собой связывающие последовательности; — характеризуются многофункциональностью, т. к. их домены и последовательности не обладают высокой специфичностью; — группируют сигнальные белки и способствуют образованию эффективной организации сигнального пути.
Типы адапторных белков 1. Адапторы – небольшие белки, имеющие два участка связывания для двух белков-эффекторов. 2. Платформы или скэффолд-белки – крупные белки, имеющие несколько доменов для взаимодействия, способные связывать и регулировать несколько белков сигнального пути. 3. Стыковочные или докинг-белки – имеют аналогичные структурные и функциональные особенности, что и у скэффолд-белков, но их отличительная черта – способность локализоваться на мембране рядом с активированным рецептором.
Типы адапторных белков
Домены адапторных белков, распознающих и связывающихся с фосфорилированными остатками тирозина или серина
Связывающие модули сигнальных белков
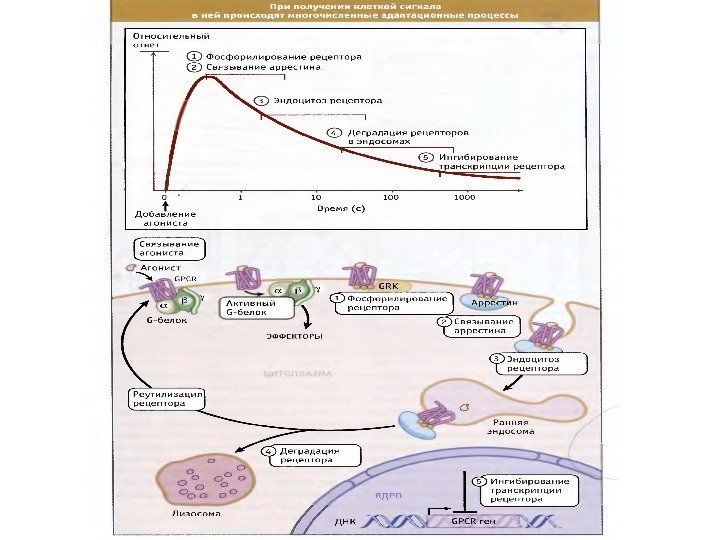
Система передачи сигнала характеризуется высокой способностью к адаптации 1. Универсальное свойство систем СТ – адаптируемость к входному сигналу. 2. Чувствительность путей СТ варьирует в широких пределах (в 100 -кратном диапазоне). 3. Системы СТ регулируются с помощью механизмов обратной связи. 4. Адаптация, селективная в отношении определенного рецептора, называется гомологичной адаптацией. 5. Адаптация, при которой регулируется активность как инициирующего, так и других рецепторов, называется гетерологичной адаптацией.
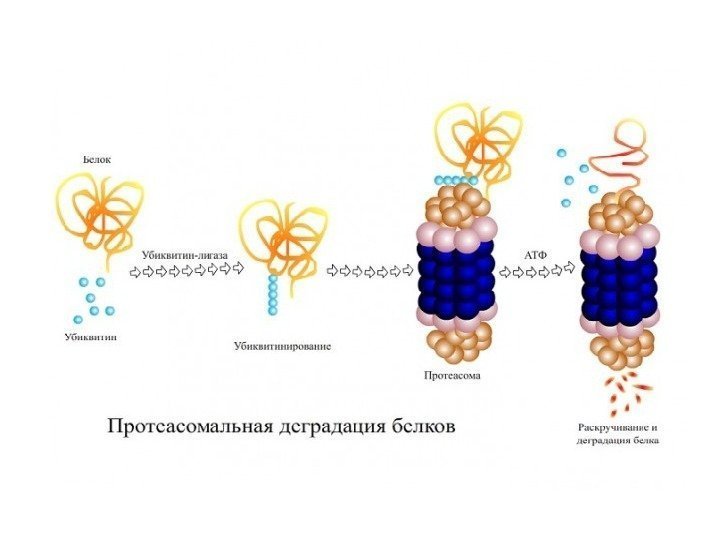
Способы регуляции сигнальных систем 1. Сигнальные белки экспрессируются в виде множественных форм, изоформы расширяет возможности регуляции и адаптации сигнальных систем. 2. Активация и инактивация – различные реакции и регулируются независимо. Разделение этих реакций обеспечивает их тонкую регулировку по амплитуде и по времени. 3. Активность белка регулируется за счет аллостерической и ковалентной модификации его структуры: — аллостерическая регуляция – способность молекулы связываться с белком-мишенью нековалентной связью и изменять его конформацию; — ковалентная модификация осуществляется путем: фосфорилирования по Тир, Сер, Тре с участием АТФ и протеинкиназ ; убиквитинилирования с помощью убиквитина и родственных белков. 4. Фосфопротеинфосфатазы аннулируют действие киназ (путем дефосфорилирования ) и регулируются независимым образом.
Множественность сигнальных G-белков и их мишени
Протеинкиназы переносят γ-фосфатную группу от АТФ на остатки Тир, Сер, Тре в белковых субстратах
Контроль функций белков с помощью убиквитина 1. Убиквитин – малый белок (76 АК) – относится к сем. убиквитин-подобных белков ( Ubl ). 2. Ubl белки образуют конъюгаты с субстратным белком за счет образования изопептидной связи между аминогруп-пой субстрата и С-концевым Гли белка Ubl с участием ферментов Е 1, Е 2, Е 3. 3. Моноубиквитинилирование – сигнальная модификация. Полиубиквитинилирование по остаткам Лиз 48 в самом убиквитине направляет белок в протеасомы для деградации. Полиубиквитинилирование по Лиз 63 способствует передаче сигнала и не приводит к деградации.
Активация и деградация фактора транскрипции NF-k. B включает взаимодействие с убиквитином
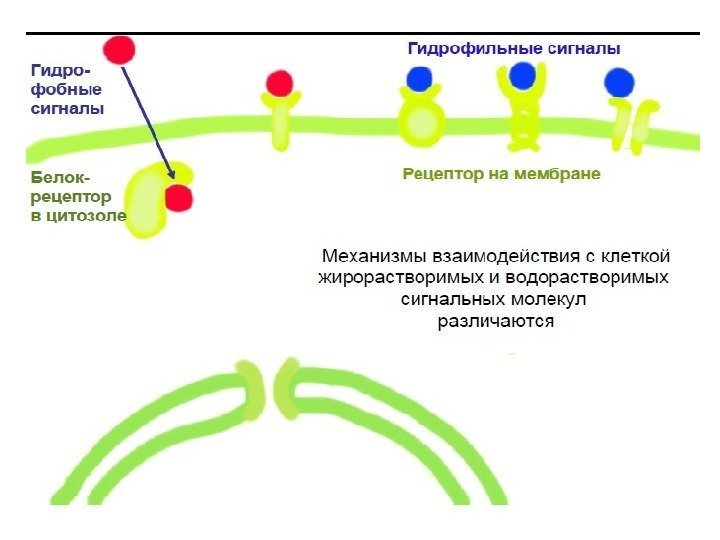
Первичные мессенджеры (ПМ) – это химические соединения или физические факторы (кванты света), которые несут информацию в клетку и активируют запуск сигнальной трансдукции. ПМ – экстраклеточные сигналы К первичным мессенджерам относятся: • Гормоны • Медиаторы (нейротрансмиттеры) • Цитокины • Эйкозаноиды • Нутриенты (АТФ, аминокислоты)
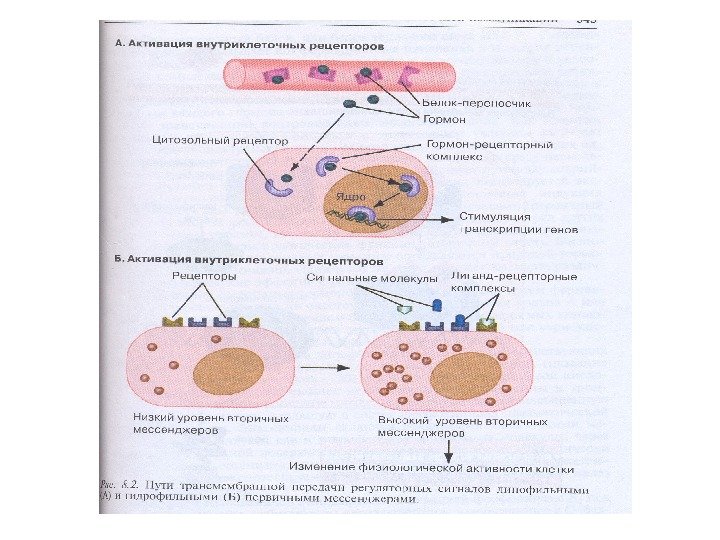
Классификация первичных мессенджеров 1. Небольшие липофильные молекулы, взаимодействующие с внутриклеточными рецепторами (стероидные, тиреоидные гормоны) 2. Липофильные молекулы, взаимодействующие с рецепторами плазматической мембраны (эйкозаноиды) 3. Гидрофильные молекулы, взаимодействующие с рецепторами плазматической мембраны (пептидные гормоны, адреналин, гистамин, цитокины, нейротрансмиттеры)
Сигнальная молекула Локализация Природа соединения Физиологические эффекты Липофильные Тиреоидный гормон (тироксин) Щитовидная железа Производное тирозина Повышение метаболизма большинства клеток Эстрадиол Яичники, плацента Производное холестерола Влияние на развитие и поддержание женских вторичных половых признаков Тестостерон Семенники Производное холестерола Влияние на развитие и поддержание мужских вторичных половых признаков Прогестерон Яичники(желтое тело), плацента Производное холестерола Подготовка и сохранение беременности, развитие системы молочных желез Простагландин Е 2 Клетки многих типов Производное арахидоновой кислоты Болевые реакции, сокращение гладкой мускулатуры Гидрофильные Гомоны (эндокринная регуляция) Инсулин β-клетки островков Лангерганса поджелуд. железы Белок (6 к. Да): α-цепь – 21 АК, β-цепь – 30 АК Стимуляция утилизации глюкозы, синтеза липидов и белков Соматостатин Гипоталамус Пептид (14 АК) Подавление секреции соматотропина передней долей гипофиза Гормон роста (соматотропин) Передняя доля гипофиза Белок (22, 9 к. Да, 191 АК) Стимуляция синтеза в печени соматомедина-1 Сигнальные молекулы, или первичные мессенджеры
Продолжение Соматомедин-1 (инсулиноподобный фактор роста 1) Преимущественно в печени Белок (8, 4 к. Да, 70 АК) Стимуляция дифференцировки мышеч-ной, жировой, хрящевой ткани, участвует в регуляции метаболизма Са 2+ и фосфатов Адренокортикотропный гормон (АКТГ) Передняя доля гипофиза Белок (4, 7 к. Да, 39 АК) Стимулирует синтез кортизола корой надпочечников и мобилизацию жирных кислот из адипоцитов Паратгормон Паращитовидные железы Белок (10 к. Да, 84 АК) Усиливает резорбцию кости, ↑ уровень Са 2+ и фосфата в крови; усиливает реабсорбцию почками Са 2+ и Mg 2+ и ↓ реабсорбцию фосфата Эпидермальный фактор роста (ЭФР) Клетки эпителиаль-ного происхождения Белок (6 к. Да) Стимулирует пролиферацию эпидерми-са и др. клеток Тиреолиберин Гипоталамус Пептид (3 АК) Стимуляция синтеза ТТГ передней долей гипофиза Тиреотропный гормон (ТТГ) Передняя доля гипофиза Белок (24, 5 к. Да, α-цепь – 92 АК, β-цепь – 112 АК) Стимулирует синтез тироксина в щитовидной железе и мобилизацию жирных кислот из адипоцитов Адреналин Мозговое вещество надпочечников Катехоламин Стимулирует распад гликогена в пече-ни и мышцах. Мобилизация жиров. Локальные химические медиаторы (паракринная регуляция) Гистамин Тучные клетки Производное гистидина Участие в воспалении. Вызывает рас-ширение и увеличение проницаемости кровеносных сосудов
Продолжение Фактор роста нервов Кожа, ткани, иннервируемые симпатическими нервами Белок (14 к. Да, 2 цепи по 118 АК) Регуляция роста и поддержание жизнеспособности нейронов Нейротрансмиттеры (синаптическая передача) Глицин Нервные окончания Аминокислота Тормозной медиатор в ЦНС γ-аминомасляная кислота (ГАМК) Нервные окончания Аминокислота Тормозной медиатор в ЦНС Энкефалин Нервные окончания Пептид (5 АК) Торможение передачи болевых сигналов в ЦНС Норадреналин Нервные окончания Производное фенилаланина Возбуждающий медиатор ЦНС и периферической нервной системы Ацетилхолин Нервные окончания Эфир уксусной кислоты и холина Возбуждающий и тормозной медиатор в ЦНС и периферической нервной системе; возбуждающий медиатор в нервно-мышечном соединении
Этапы клеточной сигнализации посредством первичных мессенджеров (ПМ) 1. Биосинтез ПМ в секреторных клетках 2. Секреция ПМ 3. Транспорт ПМ к клеткам-мишеням 4. Восприятие сигнала специфическими рецепторами 5. Изменение клеточного метаболизма или экспрессии генов 6. Устранение действия сигнального механизма и прекращение клеточного ответа на сигнал
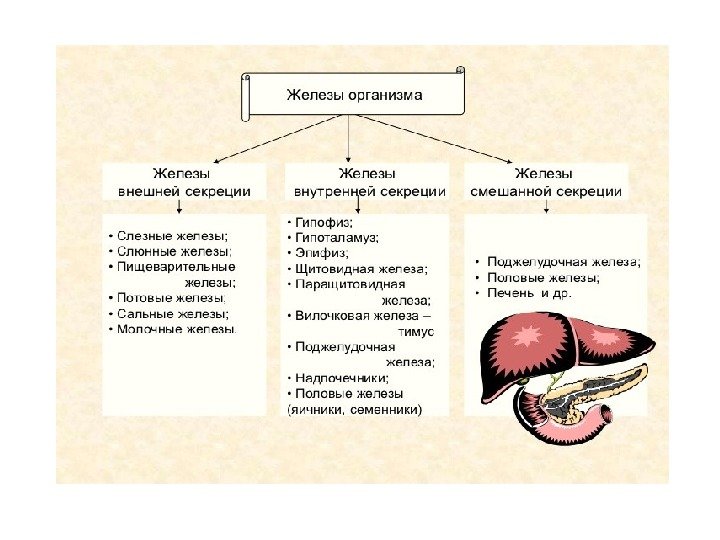
Гормоны Эндокринная система — система регуляции деятельности внутренних органов посредством гормонов, выделяемых эндокринными клетками непосредственно в кровь либо диффундирующих через межклеточное пространство в соседние клетки.
Гормоны: определение и функции Гормоны (греч. ρμάω — возбуждаю, ὁ побуждаю) — биологически активные вещества органической природы, вырабатывающиеся в специализированных клетках желёз внутренней секреции , поступающие в кровь , связывающиеся с рецепторами клеток-мишеней и оказывающие регулирующее влияние на обмен веществ и физиологические функции. Гормоны служат гуморальными (переносимыми с кровью) регуляторами определённых процессов в различных органах и системах. Физиологическое действие: — гуморальная регуляция биологических процессов; — поддержание целостности и постоянства внутренней среды организма; — регуляция процессов роста, созревания и репродукции; — регуляция активности всех клеток организма; — гормоны влияют на остроту мышления и физическую подвижность, телосложение и рост, определяют рост волос, тональность голоса, половые признаки и поведение.
1 ) Эффекты гормонов проявляются в крайне малых концентрациях — в диапазоне от 10 − 6 до 10 − 12 М. 2) Гормоны поступают в межклеточную жидкость и кровь из секретирующих клеток; эндокринные железы не имеют выводных протоков , как железы внешней секреции. 3) Гормон действует только на определенные органы-мишени , имеющие специальные рецепторы для взаимодействия с ним. 4 ) Гормон не является источником энергии и строительным материалом для нужд организма. 5) Центральная нервная система контролирует действие гормонов и оказывает определяющее влияние на их взаимодействие с организмом. 6) Между гормонами и железами внутренней секреции , их вырабатывающими, существует как прямая , так и обратная связь , объединяющая их в общую систему. Специфические особенности гормонов:
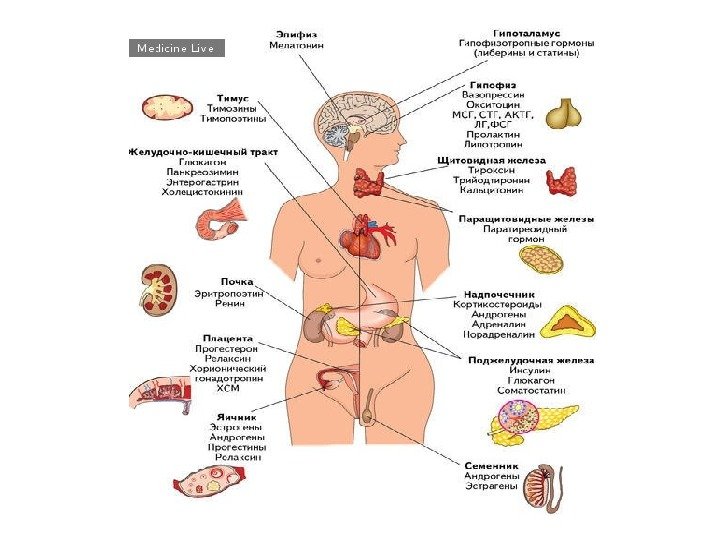
Эндокринная система и ткани мишени
Классификация гормонов 1. По месту синтеза 2. По химической природе 3. По биологическим функциям 4. По механизму действия
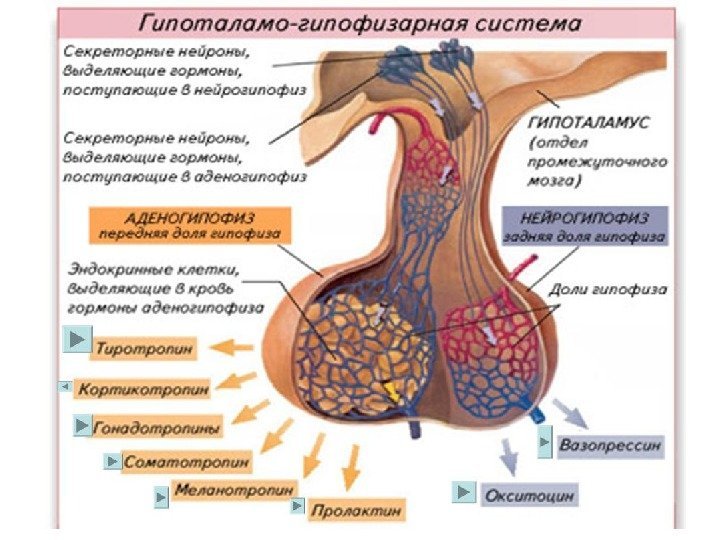
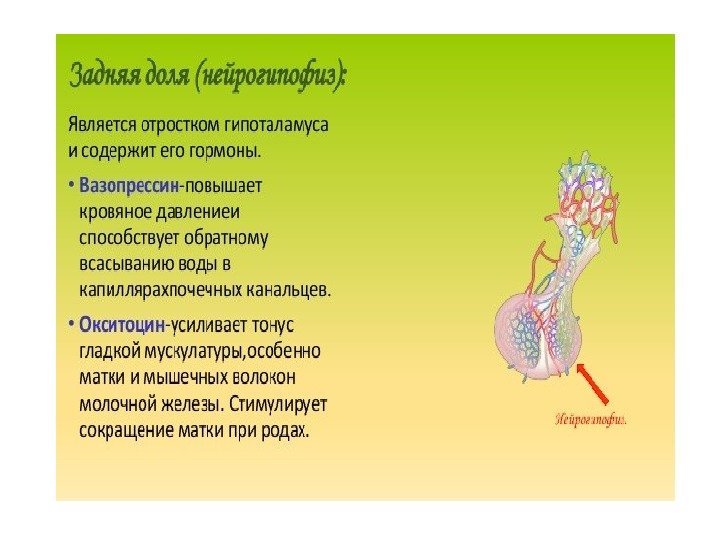
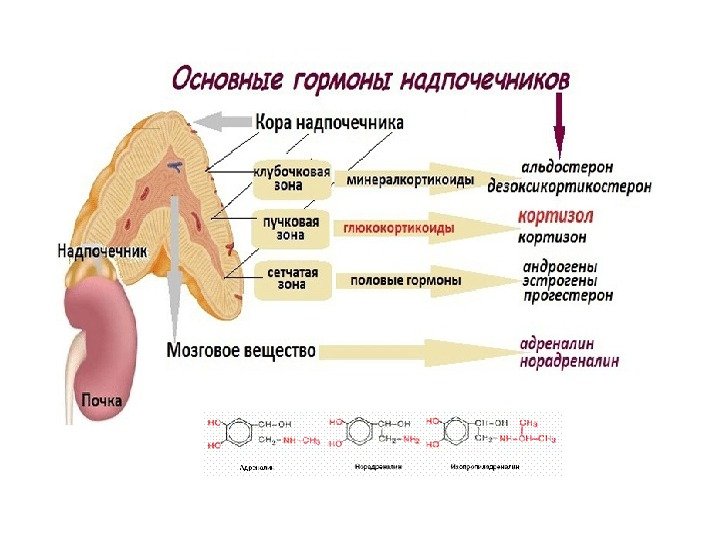
Классификация гормонов по месту синтеза Гормоны центральных эндокринных желез. Гормоны гипоталамуса: 1. Нейропептиды (либерины и статины). 2. Вазопрессин и окситоцин. Гормоны гипофиза: передняя доля (аденогипофиз): – СТГ (соматотропный гормон); – АКТГ (адренокортикотропный гормон); – липотропный гормон (альфа – и бета-липотропины); – ТТГ (тиреотропный гормон). Гонадотропные гормоны: – ФСГ (фолликулин стимулирующий гормон); – лютеонизирующий гормон (ЛГ); – пролактин (лютеотропный гормон); средняя доля – МСГ (меланоцитстимулирующий гормон) задняя доля (нейрогипофиз): – вазопрессин гипоталамуса; – окситоцин гипоталамуса. Гормоны эпифиза: 1. Мелатонин. 2. Адреногломерулотропин. Гормоны щитовидной железы: 1. Иодтиронины – тироксин, трииодтиронин. 2. Тиреокальцитонин. Гормон паращитовидной железы: – паратгормон. Гормон тимуса – тимозин. Гормоны поджелудочной железы: 1. Инсулин. 2. Глюкагон. Гормоны надпочечников: Гормоны коркового вещества: 1. Минералокортикоиды – альдостерон, 11 -дезоксикортикостерон. 2. Глюкокортикоиды – кортизон, кортикостерон, гидрокортизон. 3. Адрогены и эстрогены. Гормоны мозгового вещества: адреналин, норадреналин. Гормоны половых желез: Андрогены: тестостерон, андростерон, метитестостерон; Гормоны фолликул яичников: эстрогены- эстрон, эстрадиол, эстриол. Гормоны желтого тела: 1. Гестагены: прогестерон (лютеостерон). 2. Реклаксин.
Классификация гормонов по химической природе 1. Гормоны сложные белки (глипопротеины): ФСГ, лютеонизирующий гормон (ЛГ), ТТГ. 2. Гормоны простые белки и пептиды : СТГ, пролактин, АКТГ, вазопрессин, окситоцин, МСГ, паратгормон, тиреокальцитонин, инсулин, глюкагон. 3. Гормоны производные аминокислот: адреналин, тироксин, трийодтиронин, мелатонин. 4. Стероидные гормоны: андрогены, эстрогены, гестагены, минералокортикоиды и глюкокортикоиды.
Классификация гормонов по биологическим функциям 1. Гормоны, регулирующие обмен белков, жиров и углеводов : инсулин, глюкагон, адреналин, глюкокортикоиды. 2. Гормоны, регулирующие водно-солевой, минеральный обмен: минералокортикоиды, вазопрессин, паратгормон, тиреокальцитонин. 3. Гормоны, регулирующие энергетический обмен: тироксин, трийодтиронин, адреналин. 4. Гормоны – регуляторы репродуктивных функций организма : половые гормоны (андрогены, эстрогены), окситоцин, пролактин, гестагены. 5. Гормоны – регуляторы функций эндокринных желез: тропные гормоны гипофиза (СТГ, ТТГ, ФСГ, АКТГ, лютеинизирующий гормон). 6. Гормоны – регуляторы тропных гормонов гипофиза (статины, либерины).
Классификация гормонов по механизму действия 1. Гормоны мембранно-цитозольного механизма, реализующие эффекты на ферменты через циклазы, ц. АМФ или ц. ГМФ и протеинкиназы или Са 2+. 2. Гормоны цитозольного механизма, усиливающие синтез ферментов. 3. Гормоны мембранного механизма действия, изменяющие проницаемость мембран для субстратов.
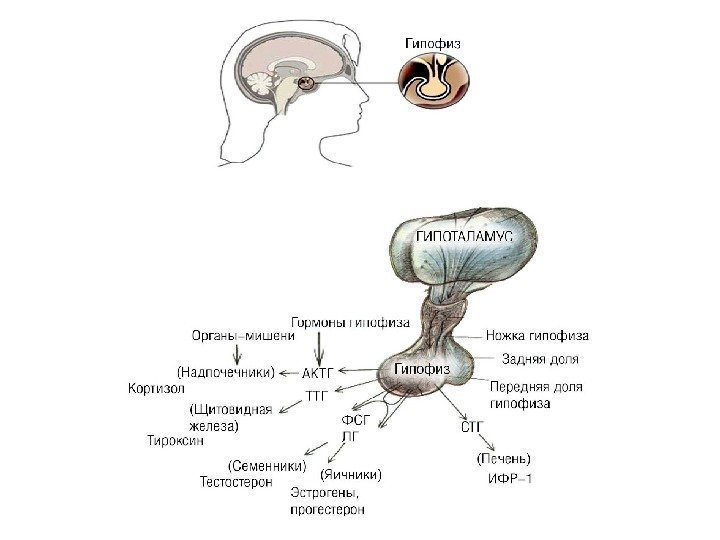
По механизму регуляторного действия гормоны делятся на 3 группы 1. Гормоны 1 -го порядка – истинные, действуют на орган-мишень (инсулин, глюкагон, тиреоидные гормоны и др. ). 2. Гормоны 2 -го порядка – тропные гормоны , влияют на эндокринные железы (гормоны передней доли гипофиза). 3. Гормоны 3 -го порядка – гормоны гипоталамуса (статины и либерины) управляют тропными гормонами.
Гормоны передней доли гипофиза управляют эндокринными железами
Кора надпочечников вырабатывает кортикостероиды. Часть из них регулирует работу почек, а часть (глюкокортикоиды, главный из них – кортизол) управляет обменом глюкозы в организме, в частности, усиливает ее образование из других веществ (белков, жиров); особенно ярок этот эффект при стрессе Выделение глюкокортикоидов усиливает кортикотропин = адренокортикотропный гормон (АКТГ). Выделение АКТГ активирует кортиколиберин (CRH). Кортизол через гипоталамус тормозит выброс АКТГ и CRH.
Щитовидная железа — небольшой эндокринный орган, лежащий на передней поверхности шеи чуть выше грудины. Он состоит из правой и левой долей, которые связаны между собой небольшим перешейком Щитовидная железа вырабатывает три гормона-тироксин (Т 4), трийодтиронин (Т 3) и кальцитонин. Два первых регулируют процессы обмена жиров, белков и углеводов, функцию сердечно-сосудистой системы, желудочно-кишечного тракта, психическую и половую деятельность, кальцитонин — один из факторов управления обменом кальция в клетках, участник процессов роста и развития костного аппарата. Как избыточная (гипертиреоз, тиреотоксикоз), так и недостаточная (гипотиреоз) функциональная активность щитовидной железы является причиной разнообразных заболеваний. На деятельность щитовидки влияют гормоны гипофиза — гормон ТТГ и гипоталамуса — гормон ТРГ (тиреотропин-релизнг-фактор).
Гормоны щитовидной железы, трийодтиронин (Т 3) и тироксин (Т 4) – это гормоны на основе тирозина, вырабатываемые щитовидной железой, отвечающие за регулирование обмена веществ. Для производства Т 3 и Т 4 необходим йод.
Нейроэндокринная регуляция половых гормоно в
Половые гормоны
Связь между центральной нервной системой и эндокринной системой
Различия эндокринной и синаптической сигнализации Эндокринная сигнализация Синаптическая сигнализация Низкая скорость передачи сигнала Высокая скорость передачи сигнала Низкие действующие концентрации Высокая специфичность и точность сигнала Высокое сродство к рецептору Возможность импульсной регуляции активности клетки-мишени Высокие действующие концентрации и низкое сродство нейромедиатора к рецептору
Медиаторы Нейромедиаторы (от лат. mediator посредник: синоним — нейротрансмиттеры ) — это биологически активные вещества, секретируемые нервными окончаниями и обеспечивающие передачу нервного импульса в синапсах нейронов, а также от нейронов к мышечной ткани.
Критерии нейромедиаторов 1. Вещество выделяется из нейрона при его активации. 2. В клетке присутствуют ферменты для синтеза данного вещества. 3. В соседних клетках (клетках-мишенях) выявляются белки-рецепторы, активируемые данным медиатором. 4. Фармакологический (экзогенный) аналог имитирует действие медиатора. Иногда медиаторы объединяют с модуляторами, то есть веществами, которые прямо не участвуют в процессе передачи сигнала (возбуждения или торможения) от нейрона к нейрону, но могут, однако, этот процесс существенно усиливать или ослаблять.
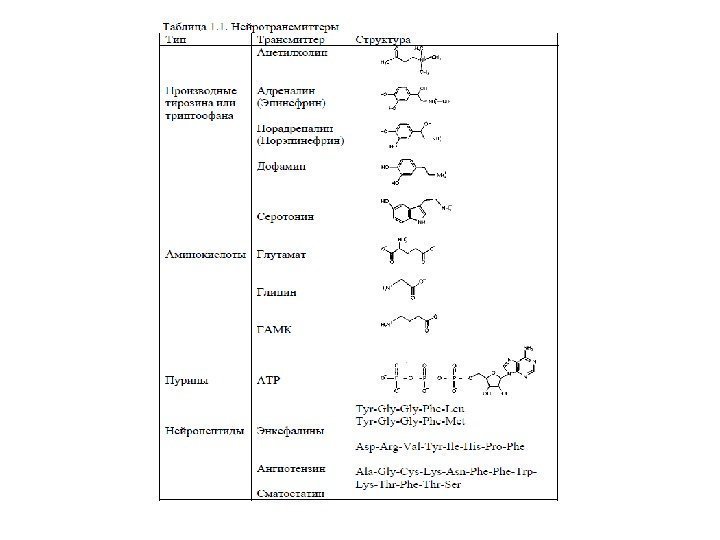
Основные медиаторы: 1. Ацетилхолин 2. Дофамин 3. Норадреналин 4. Серотонин 5. Глутамат 6. ГАМК 7. Глицин 8. SP 9. Опиоиды 10. Нитроксид (NO) 11. Пурины (АТФ, Аденозин ) и др. катехоламины биогенные амины Принцип Дейла : нейрон представляет собой единую метаболическую систему и выделяет один и тот же медиатор во всех терминалях. аминокислоты Нейропептиды Обычно выделяться несколько медиаторов совместно, либо медиатор и несколько модуляторов. Роль модуляторов: Изменяют выброс медиатора Изменяют чувствительность рецепторов
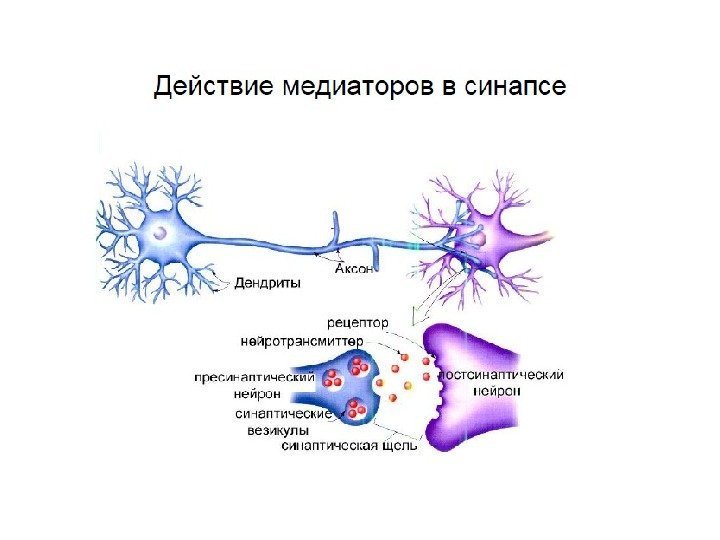
Синапсы — специализированные функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. Термин «синапс» ввел Ч. Шеррингтон (С. Sherrington, 1897).
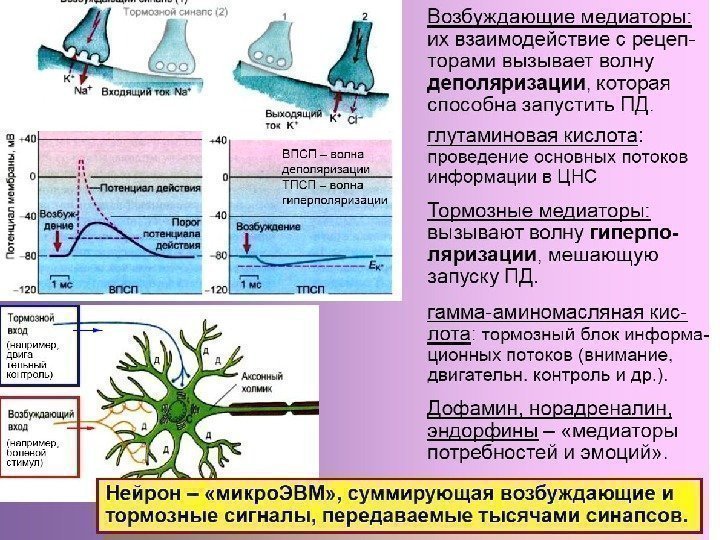
Основные стадии передачи сигнала в синапсе 1. Потенциал действия ( ПД) запускает движение молекул и выброс медиатора в синаптиче-скую щель 2. Медиатор взаимодействует с рецепторами постсинаптической мембраны. 3. Рецепторы вызывают возбуждение или торможение следующей клетки. Возбуждение вызывает генерацию ПД , торможение препятствует возникновению ПД , затрудняет или блокирует проведение сигнала.
Ведущая роль в синаптической передаче принадлежит медиатору. 4 этапа превращений медиатора в синапсе 1. Синтез и накопление в пресинаптическом окончании. 2. Выброс медиатора в синаптическую щель при появлении ПД. 3. Действие на рецепторы постсинаптической мембраны (запуск возбуждения или торможения постсинаптической клетки). 4. Инактивация (прекращение действия медиатора на рецептор).
Если синтез идет в теле нейрона , то: 1) Медиатор по ЭПС переносится в комплекс Гольджи 2) Комплекс Гольджи образует везикулы с медиатором 3) Везикулы по аксону переносятся в пресинаптическое окончание, где и накапливаются
Если синтез идет в пресинаптическом окончании , то далее медиатор поступает а пустые везикулы с помощью белков-транспортеров. 1) Комплекс Гольджи поставляет пустые везикулы 2) Наполнение везикул медиатором 3) Отделение пустых везикул от пресинаптической мембраны А (пустая везикула) медиатор Б (загрузка медиатора) В (везикула заполнена (5 -10 тыс. молекул) «Круговорот» везикул в пре- синаптическом окончании
1. Экзоцитоз медиатора в синаптическую щель происходит после появления потенциала действия (ПД), который вызывает открывание φ-чувствительных Са 2+ -каналов, что приводит к входу в пресинаптическое окончание ионов Са 2+ , которые активируют белки, запускающие экзоцитоз. Для экзоцитоза одной везикулы требуется не менее 5 ионов Са 2+. 2. Специфические белки-насосы удаляют Са 2+ из пресинаптического окончания, иначе выброс медиатора не прекратится. 3. Приход одного ПД вызывает выброс содержимого 50 везикул.
Яд каракурта ( «черная вдова» ) содержит токсин, представляющий собой белок, сходный с постоянно открытым Са 2+ -каналом. После укуса паука токсин встраивается в мембрану пресинаптического окончания, вызывая мощный вход Са 2+ , выброс медиатора и судороги. Затем запас медиатора истощается, наступает паралич и остановка дыхания. Токсин почвенной анаэробной бактерии ботулизма блокирует белки, отвечающие за экзоцитоз. При отравлении (если бактерия находится в консервах) развивается слепота, параличи, смерть. Clostridium botulinum. Каракурт
Инактивация – это удаления медиатора с рецептора для предотвращения слишком длительной передачи сигнала В каждом синапсе используется один из 3 путей инактивации: 1) разрушение медиатора с помощью фермента 2) перенос медиатора в пресинаптическое окончание ( обратный захват ) 3) перенос медиатора в глиальные клетки
Расщепление, удаление и обратный захват трансмиттера на примере ацетилхолина
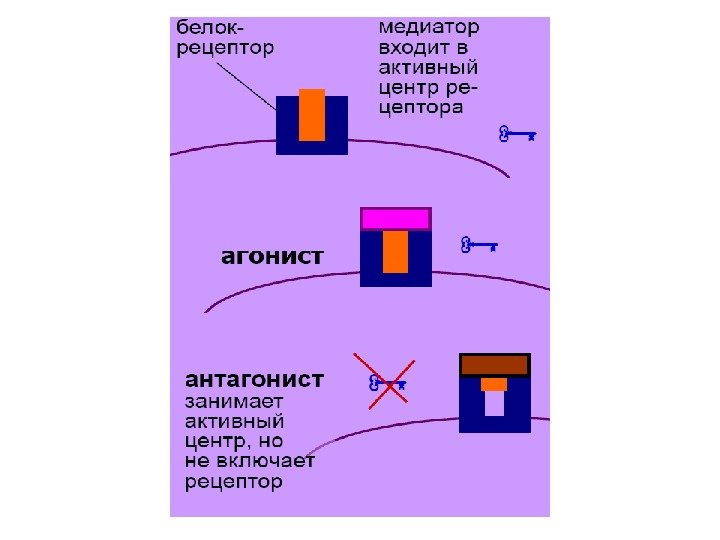
АГОНИСТЫ И АНТАГОНИСТЫ РЕЦЕПТОРОВ МЕДИАТОРОВ • Агонист: вещество, действующее как медиатор; обычно – сильнее и длительнее. Молекула состоит из ключевой и защитной частей. Ключевая часть сходна с медиатором и включает рецептор; защитная часть мешает работать системам инактивации. • Антагонист: вещество, противодействующее эффектам медиатора. Молекула состоит из защитной части и неполной ключевой части. Последняя из них занимает активный центр рецептора, но не включает его, работая как «сломанный» ключ и мешая медиатору. • Антагонисты и агонисты рецепторов – вещества, поступающие в организм извне. Многие из них являются токсинами, которые возникли в ходе эволюции растений для защиты от животных. При разбавлении их человек получает лекарства; не разбавив – яды и наркотики.
Ацетилхолин (5% нейронов, главный медиатор периферической НС) Синтез из холина и ацетил-Ко. А в ЦНС ( кора, таламус, хвостатое ядро, ретикулярная формация), вегетативных ганглиях, мотонейронах. Рецепторы: 2 типа — Н (никотиновые): Н 1 — мышечные, Н 2 — нейронные. Действие на катионный канал, Блокируются курарином. М (мускариновые): М 1, М 2, М 5 – действие на ИФ 3 путь М 2 – действие на Gi (открытие К+ канала) Блокатор – атропин (беладонна) Роль: движение, мнестические процессы (старческое слабоумие)
АЦЕТИЛХОЛИН: Первый описанный нейромедиатор. Обеспечивает сокращения скелетных мышц, замедляет сердечный ритм и понижает артериальное давление, отвечает за внимание и обучение. Сначала была доказана его роль в работе вегетативной нервной системы (ВНС), затем – нервно-мышечных синапсов, позже – ЦНС (центральной нервной системы). В 1936 г. Отто Лёви и Генри Дейл (H. Dale) получили Нобелевскую премию за «открытие химического механизма синаптической передачи» .
Появление потенциала действия (ПД) запускает выброс Ац. Х в синапти-ческую щель, после чего медиатор действует на рецепторы. Известны 2 типа рецепторов к Ац. Х: 1. Никотиновый рецептор реагирует на Ац. Х и никотин (токсин табака) — агонист. Антагонист – курарин. 2. Мускариновый рецептор реагирует на Ац. Х и мускарин (токсин мухомора) — агонист. Антагонист – атропин. 3. Курарин и атропин – ацетилхолин-подобные растительные токсины. Белена
• Никотин как агонист рецепторов Ац. Х защищает растения от поедания насекомыми; для человека – слабый разрешенный наркотик. • Никотин при курении не влияет на нервно-мышечные синапсы. • При первых попытках курения никотин стимулирует парасимпатические нейроны (парасимпатические эффекты: тошнота, скачки давления). • Позднее эти эффекты исчезают и сменяются стимуляцией симпатических нейронов (активация ССС, ослабление сигналов от ЖКТ). • Никотин также способен оказывать нормализующее действие
• Возможно проникновение никотина через ГЭБ и формирование привыкания и зависимости. • Формирование привыкания и зависимости – типичное следствие приема препаратов, влияющих на мозг. • Для восстановления деятельности Ац. Х-синапсов требуются недели и месяцы. • Явный признак наличия зависимости – с утра хочется курить и первая сигарета доставляет наибольшее удовольствие.
• Курарин (основное действующее вещество яда кураре; кора растений рода Strychnos) – антагонист рецепторов Ац. Х. • Препятствует присоединению Ац. Х к рецепторам. • Основное действие оказывает на нервно-мышечные синапсы (паралич, остановка дыхания). • Используется аборигенами для отравления стрел на охоте. • В клинике – для выключения нервно-мышечных синапсов и сокращения мышц во время длительных хирургических операций (при этом пациент подключается к аппарату искусственного дыхания).
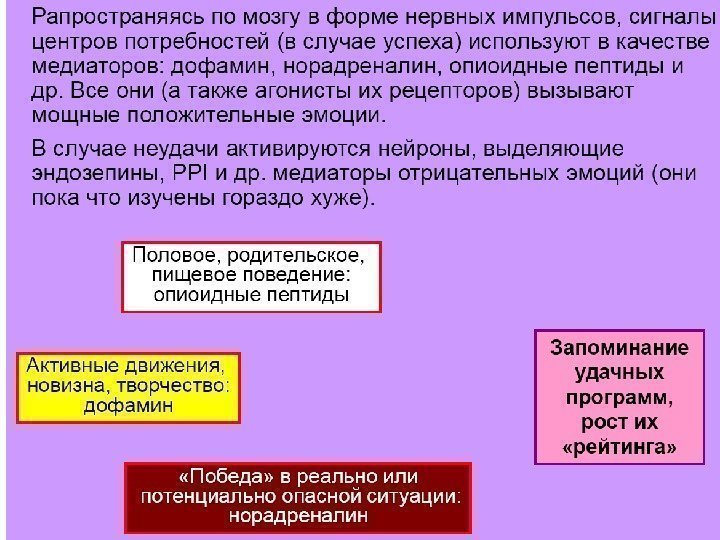
Дофамин (ДА) (1 -2% нейронов, мотивационно-эмоциональная сфера) Синтез из тирозина в ЦНС (черная субстанция, вентральная покрышка, гипоталамус) Рецепторы: 5 типов (метаботропные) Д 1: активируют аденилатциклазу Д 2: подавляют аденилатциклазу Агонист: бромкриптин ( алкалоид спорыньи: тормозит синтез пролактина, лечение бесплодия ) Антагонисты – аминазин, галоперидол (нейролептики ) Роль: контроль произвольных движений ( паркинсонизм) , пищевое поведение, положительные эмоции ( шизофрения )
Синтез дофамина 1. Тирозин превращается в L-DOPA (диоксифенилаланин), фермент – тирозин-гидроксилаза 2. L-DOPA путем декарбоксилирования превращает-ся в дофамин Дофамин образуется в высоких концентрациях в нейронах: А) черной субстанции среднего мозга (аксоны идут в базальные ганглии) Б) покрышки среднего мозга (аксоны идут в кору больших полушарий) В) гипоталамуса (короткие аксоны, локальные влияния, эндокринная функция)
Цикл дофамина в синапсе 1. Синтез в пресинаптическом окончании и экзоцитоз при появлении ПД. 2. Действие на постсинаптичес-кие рецепторы , сопряженные с G-белками. 3. Действие на пресинаптичес-кие рецепторы – аутоторможение экзоцитоза. 4. Инактивация : обратный захват и последующее повторное использование или разрушение с помощью моноаминоксидазы (МАО). МАО – фермент, расщепляющий моноамины
Эффекты дофамина, вырабатываемого нейронами разных зон ЦНС (гипоталамусом, черной субстанцией, ядрами вентральной покрышки среднего мозга) Гипоталамус – главный центр эндокринной и вегетативной регуляции, а также биологических потребностей и связанных с ними эмоций (голод и жажда, страх, агрессия, половая и родительская мотивация). 1. ДА гипоталамуса оказывает тормозящее действие на секрецию гипофизом пролактина , который активирует лактацию, родительское поведение, а также тормозит половую мотивация и овуляцию. ДА(гипоталамус) →пролактин↓(гипофиз) 2. ДА оказывает тормозное влияние на центры одних биологических потребностей (голод, страх, тревожность) и активирующее – на центры других (половое поведение, агрессия). 3. При периферическом введении ДА не проходит ГЭБ и превращается в адреналин и норадреналин, является мягким кардиостимулятором.
Эффекты дофамина черной субстанции среднего мозга Черная субстанция среднего мозга : ДА аксоны идут в базальные ганглии, определяя общий уровень двигательной активности , положительные эмоции , связанные с движениями (танцы, физические упражнения)
Ядра вентральной покрышки среднего мозга : аксоны идут в кору больших полушарий, регулирует скорость обработки сенсорной информации , скорость мышления, положительные эмоции, связанные с получением новых знаний, юмором (мезокортикальная система)Дофамин в ядрах вентральной покрышки среднего мозга
Гомеостаз – поддержание постоянства внутренней среды организма (состава крови, температуры и т. п. ) Потребность — реальное или прогнозируемое нарушение гомеостаза, которое может быть предотвращено с помощью целенаправленного поведения. Пример: потребность в пище. Мотивация — эмоционально окрашенное состояние, направляющее поведение на удовлетворение потребности. Пример: ощущение голода. Эмоция — субъективная реакция, проявляющаяся в виде удовольствия или неудовольствия, страха, гнева, тоски, радости, надежды, грусти и т. п. Пример: неудовольствие, раздражение при голоде.
Эмоции являются важнейшим подкрепляющим фактором и закрепляют обучение : и человек, и животное стремятся повторять те действия которые ведут к положительным эмоциям, и избегать тех, которые ведут к отрицательным эмоциям.
Синтез из дофамина, фермент — дофамин -β-гидроксилаза в ЦНС: (голубое пятно — ок. 1000 нейонов), симпатические нервы, но аксоны расходятся по всему головному мозгу Рецепторы: α 1: , агонист: клонидин. β 1, β 2: Gs белок, агонист: изопротеренол, Норадреналин (НА) (<1% нейронов, главный медиатор симпатической НС, медиатор стресса) Роль: обучение, эмоции, настроение, подавление боли, бодрствование Антагонисты — нейролептики с седативным эффектом (резерпин, аминазин), Агонисты — психостимуляторы (амфетамин, кокаин).
Синтез адреналина 1. Тирозин превращается в L-DOPA (диоксифенилаланин), фермент – тирозин-гидроксилаза 2. L-DOPA путем декарбоксилирования превращается в дофамин 3. ДА превращается в норадреналин (NE) 4. Из норадреналина в надпочечниках образуется адреналин.
2. Появление ПД запускает вход Са 2+ и выброс НА в синаптическую щель, после чего он действует на рецепторы как постсинаптической, так и пресинаптической мембраны. 2. Известны 2 типа рецепторов к НА: альфа- и бета-адренорецепторы (α- и β-). Они, в свою очередь, подразделяются на α 1 -, α 2 -, β 1 — и β 2 -подтипы. 1. Синтез НА – в пресинаптическом окончании, после чего НА переносится внутрь везикул и готов к экзоцитозу.
НА влияет на нейроны ЦНС через α- и β-рецепторы Основные эффекты НА — «психическое сопровождение стресса» : — общая активация деятельности мозга (торможение центров сна, бессонница) — увеличение двигательной активности («не сидится на месте») — снижение болевой чувствительности (стресс-индуцированная анальгезия) — улучшение обучения, запоминания (на фоне умеренного стресса, «учимся избегать опасности») — положительные эмоции при стрессе (азарт, «чувство победы», «экстрим»)
Положительные эмоции, связанные с выделением НА и адреналина : спорт, экстремальный спорт, игромания (компьютерные игры, казино). Зависимость от НА — реальная проблема; игроманию лечат в тех же клиниках и теми же методами, что и наркоманию. С системой НА и адреналина связаны реакции страха, агрессии, особенности темперамента человека. Стресс (от «stress» : напряжение) — неспецифическая, общая реакция организма на сильное и очень сильное воздействие (физическое или психологическое), а также соответствующее состояние нервной системы и организма в целом.
Серотонин (5 -НТ) – 1 -2% нейронов, мотивационно-эмоциональная сфера Синтез из 5 — окситриптофана (источник- триптофан) в ЦНС (ядра шва), мозговом слое надпочечников, ЖКТ Рецепторы: 7 типов 5 НТ 1: Gi белок (тормозной) 5 НТ 2: ИФ 3 -путь (возбуждающий) 5 НТ 3 катионный канал 5 НТ 4 -7 Gs белок Блокаторы: ципрогептадин Роль: терморегуляция, сон, циркадные ритмы, тревожность, депрессия и агрессия, сенсорное восприятие (агонист — LSD — диэтиламид лизергиновой к-ты из спорыньи; синестезия )
Серотонин – медиатор ЦНС; его вырабатывают нейроны ядер шва ( «ствол» мозга); их отростки-аксоны расходятся по всей ЦНС, образуя контакты (синапсы) Серотонин (5 -НТ) в ЦНС вызывает чаще тормозные эффекты; нередко конкурирует с влияниями НА. Центральное серое вещество среднего мозга и ядра шва: — главные центры сна — снижают фоновый уровень болевой чувствительности — блокируют слабые сигналы в коре б. п/ш. (снижают «шум» ) — снижают активность центров отрицательных эмоций (антидепрессантное действие). При депрессиях – нарушение равновесия между центрами положительных и отрицатательных эмоций (гипоталамус, базальные ганглии), прежде всего, из-за ослабления положит. эмоций ( «жизнь не радует» ). Антидепрессанты – препараты, активирующие 5 -НТ, NE и DA синапсы. В больших дозах – психостимулирующее действие, привыкание, зависимость. 5 -НТ сдерживает отрицательные эмоции (прозак), а НА и DA поднимают уровень положительных эмоций. Препараты, нарушающие работу серотонина в мозге, способны вызвать галлюцинации (ЛСД и другие наркотики-психоделики)
Периферические эффекты серотонина 1. Регулирует тонус сосудов – вазоконстрикция. 2. Выделяется тромбоцитами, участвует в тромбообразовании. (образование белого и красного тромба). 3. Разрушение тромбоцитов →повышение тонуса сосудов→ падение уровня 5 -НТ →избыточное расслабление сосудов мозга→ мигрень (сильная пульсирующая боль). 4. Эрготамин и др. токсины спорыньи – агонисты многих рецепторов 5 -НТ. Основа для синтеза ЛСД-25 – диэтиламида лизергиновой ктслоты (психоделика-галлюциногена). 5. В Средние века часты отравления спорыньей и развитие эрготизма (огонь святого Антония, «огненная чума» ) – спазм периферических сосудов→гангрена→отмирание органов. 6. Психотическая форма отравления спорыньей – танцевальная мания (пляска Святого Вита), массовая истерия в некоторых районах выращивания ржи.
Больной эрготизмом П. Брейгель мл. (1554 -1636) «Одержимые пляской» Святой Антоний
Глутамат Синтез из глюкозы (цикл Кребса) в коре, гиппокампе, стриатуме, гипоталамусе, таламусе и др. Инактивация: захват глутамата глией в синаптической щели Рецепторы: NMDA, AMPA: Са канал m. Glu. R 1 -5: ИФ 3 путь Антагонист: кетамин (наркоз) Агонист: каинат Роль: основной возбуждающий медиатор ЦНС, память (эпилепсия ), 40% нейронов Распад глутамата до ГАМК
ГАМК – 40% нейронов, главный тормозный медиатор Синтез из глутатмата В коpе, черной субстанции, гиппокампе, стриатуме, мозжечке, спинном мозге, и обонятельной луковице. Рецепторы: ГАМКа, ГАМКс: Cl- канал ГАМКб: Gi-белок Инактивация : обратный захват Роль: главный тормозный медиатор ЦНС, 40% нейронов, моторная и эмоциональная активность (анксиолитическая — модулятор дофаминеpгических нейpонов ), сон, память (эпилепсия)Агонисты: бензодиазепины, барбитураты (б). Антагонисты: бикукулин (а), пикротоксин (а и с), пенициллин.
Глицин Синтез из ацетил-КОА в спинном мозге, (клетки Реншоу) мосте, продолговатом мозге, гиппокампе Рецепторы: Gly. R: анионный канал агонист: таурин, антагонист: стрихнин Роль: регуляция рефлекторной деятельности
Субстсанция П (SP) Белковый синтез (11 а. к. ) в спинномозговых ганглиях, стенках полых органов (болевой вход в спинной мозг). Часто бывает модулятором (при Д, НА) (5 НТ+SP – угнетающее действие на выброс 5 НТ, 5 НТ стимулирует секрецию SP) Рецептор: Gs- белок Роль: восприятие болевого сигнала, сокращение гладких мышц Пептидные медиаторы (признано около 10 видов)
Опиоидные пептиды (группа – 13 пр. ) Действуют в концентрациях 10 -9 (в 1000 раз ниже, чем низкомолекулярные медиаторы) Синтез : В соме на рибосомах в ЭПР предшественники, (далее распад, гликозилирование, S-S мостики), в АГ (фосфорилирование, сульфатирование), далее медленный аксонный транспорт. В ЦНС (ствол, гипоталамус, кора) Роль: система положительного подкрепления, сон, обезболивание Рецепторы: , d: Gi белок : торможение Са тока Агонист: морфин (m), Антагонист: налоксон b-эндорфин H -Y-G-G-F-L-M-T-S-E-K-S-Q-T-P-L-V-T- L-F-K-N-A-I-V-K-N-A-H-K-K-G-Q- OH
Гистамин Синтез из гистидина в заднем гипоталамусе, волокна – по всему мозгу, характерен несинаптический выброс медиатора Роль: регуляция общего метаболизма — повышает уровень бодрствования, мышечная активность, пищевое и половое поведение (антигистаминовые препараты) Рецепторы: Н 1: ИФ 3 путь (+) Н 2: Gs-белок Н 3: Gi-белок (пресинаптический) Кроме медиаторной роли выполняет функцию расширения сосудов, фактора воспаления, секреции желудочного сока
Цитокины — регуляторные пептиды, продуцируемые клетками организма. К системе цитокинов в настоящее время относят около 200 индивидуальных полипептидных веществ. • Характерные свойства – плейотропность, взаимозаменяемость биологического действия, отсутствие антигенной специфичности, проведение сигнала путем взаимодействия со специфическими клеточными рецепторами, формирование цитокиновой сети. Важнейшие функции: • Регуляция эмбриогенеза, закладки и развития ряда органов, в том числе органов иммунной системы. • Регуляция отдельных нормальных физиологических функций, например, иммунитета, нормального кроветворения. • Регуляция защитных реакций организма на местном и системном уровне. • Регуляция процессов регенерации для восстановления поврежденных тканей. • Цитокины могут быть выделены в новую самостоятельную систему регуляции функций организма, существующую наряду с нервной и гормональной регуляцией.
Классификация цитокинов
Классификация рецепторов цитокинов
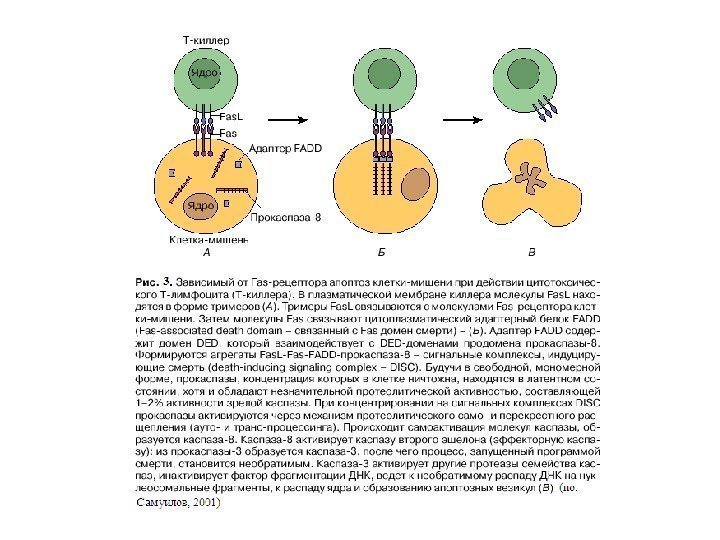
Индукция ФНО рецепторного пути апоптоза
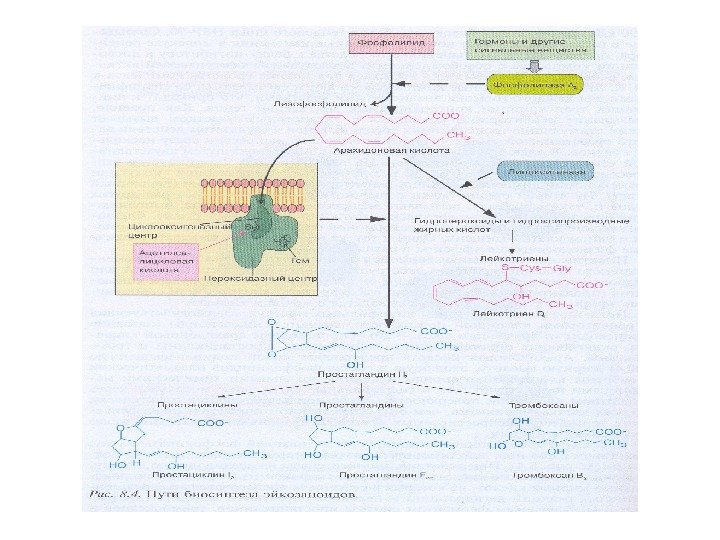
Эйкозаноиды – производные ПНЖК семейства ω-3 и ω-6, обладающие биологической активностью: простагландины, простациклины, тромбоксаны (ЦОГ) и лейкотриены (ЛОГ) • Контролируют сокращение гладкой мускулатуры (сосуды, бронхи, матка, ЖКТ) • Контролируют секрецию продуктов внутриклеточного синтеза (соляная кислота, гормоны) • Являются посредниками секреции гормонов • Регулируют болевые реакции • Регулируют свертываемость и вязкость крови • Оказывают влияние на иммунную систему • Регулируют метаболизм костной ткани
n — 7 n — 9 пальмитоеновая 16: 1 7 олеиновая 18: 1 9 n — 3 n — 6 COOH докозагексаеновая 22: 6 3 рыба арахидоновая 20: 4 6 мясо, яйца Сердечно-сосудистые заболевания, воспаление, травмы мозга психозы, шизофрения, нейродегенеративные заболевания 2. Арахидоновая кислота и другие полиеновые жирные кислоты как сигнальные молекулы
Арахидоновая кислота ПРОСТАГЛАНДИНЫ: Широкий спектр воздействий, так как регулируют синтез с. AMP в разных тканях, например: — Стимулируют сокращение гладкой мускулатуры матки; — Влияют на суточные ритмы; — Изменяют чувствительность тканей к гормонам (адреналину и глюкагону) — Вызывают повышение температуры, участвуют в развити реакции воспаления и боли. Prostaglandin E 1 (PGE 1 )Простагландин Е 1 Простагландин F 1α
Арахидоновая кислота ЛЕЙКОТРИЕНЫ — Индуцируют сокращение мускулатуры дыхательных путей; — Гиперпродукция лейкотриенов приводит к астматическому приступу Лейкотриен А
Патология сигнализации, сопряженная с нарушениями первичных мессенджеров
Функционирование и адаптация клеток зависит от 2 факторов: 1. Программного управления 2. Исполнительного аппарата Поэтому природа болезней может рассматриваться с двух позиций – 1) информационных (нарушение сигнализа-ции, рецепции); 2) материально-энергетических (нарушение ДНК, РНК, белков и др. ) Адаптация клетки к повреждению сводится к своевремен-ному включению адаптивных программ , которые опти-мально соответствуют конкретной ситуации.
I. Нарушение программного управления 1. Патология сигнализации 2. Нарушение рецепции сигналов 3. Нарушение пострецепторных механизмов передачи сигналов 4. Дефекты клеточных программ Клетки в организме управляются химическими регуляторными сигналами различных типов: — первичными посредниками ( гормоны, медиаторы, цитокины, эйкозаноиды, нутриенты); — вторичными посредниками (ц. АМФ, ц. ГМФ, IP 3, Са 2+, NO • , H 2 O 2 и др. )
Патология сигнализации реализуется по 3 направлениям: 1. Недостаток или отсутствие сигнала может препятство-вать включению адаптивных программ 2. Избыток сигнала заставляет адаптивные программы функционировать слишком интенсивно или ненормально длительно 3. Мимикрия сигналов – рецептор, контролирующий включение определенных программ стимулируется или блокируется аномальным сигналом, ошибочно принятый клеткой за гормональный или медиаторный стимул ( напри-мер, выработка аутоантител – феномен «создание иммуно-логического имиджа» того или иного сигнала)
Типы первичных сигналов Избыток сигнала Дефицит сигнала Мимикрия сигнала Гормон Синдром Иценко-Кушинга (гиперкортицизм – избыток кортизола) Инсулинзависимый сахарный диабет (недостаток инсулина) Базедова болезнь (антитела к рецептору ТТГ – иммуноло-гичес кий имидж сигнала) Нейромедиатор Алкоголизм (избыток дофамина) Ожирение (избыток субстратов) Шизофрения ( избыток дофамина — позитивные эпизоды) Болезнь Паркинсона (дефицит дофамина) Шизофрения (недостаток дофамина – негативные эпизоды) Алкоголизм ( «ложные медиаторы» — ТГИХ. β-карболи-ны)Патология сигнализации
Гипоталамо-гипофизарно-надпочечникова я ось. Болезнь Иценко-Кушинга (БИК) (гиперкортицизм ) • ИЗБЫТОК СИГНАЛА • БИК – тяжелое нейроэндокринное заболевание, связанное с нарушением работы гипоталамо-гипофизарно-надпочечниковой системы и гиперпродукцией кортикостероидов (гиперкортицизм) • Нарушение в гипофизе → гиперпродукция АКТГ → гиперплазия надпочечников → гиперпродукция кортикостероидов (кортизола) → нарушение метаболизма • Избыток кортикостероидов: • повышение массы тела, ожирение • повышение катаболизма белков → истончение кожи, гематомы • поражение сердечно-сосудистой системы • нарушение электролитного обмена • гиперинсулинемия, инсулинорезистентность (СД II) • вторичный иммунодефицит • системный остеопороз
Алкогольная болезнь – избыток (дофамин) и мимикрия сигнала (ТГИХ, β-карболины)
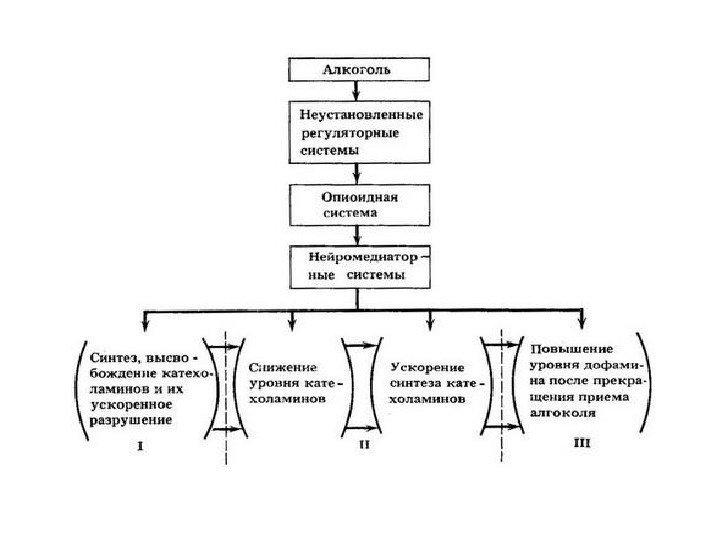
Болезни аддикции (пристрастия) – алкоголизм, наркомания — связаны с глубокими нарушениями медиаторного обмена (катехоламины, опиоиды) в ЦНС
ИЗБЫТОК СИГНАЛА
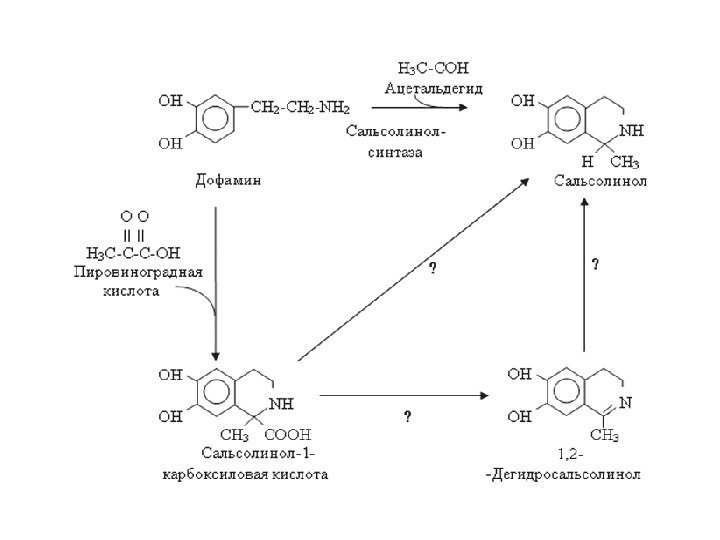
МИМИКРИЯ СИГНАЛА Схема образования эндогенных опиатоподобных веществ в результате употребления алкоголя.
Схема оптимальной работы синапса Медиаторный дефицит
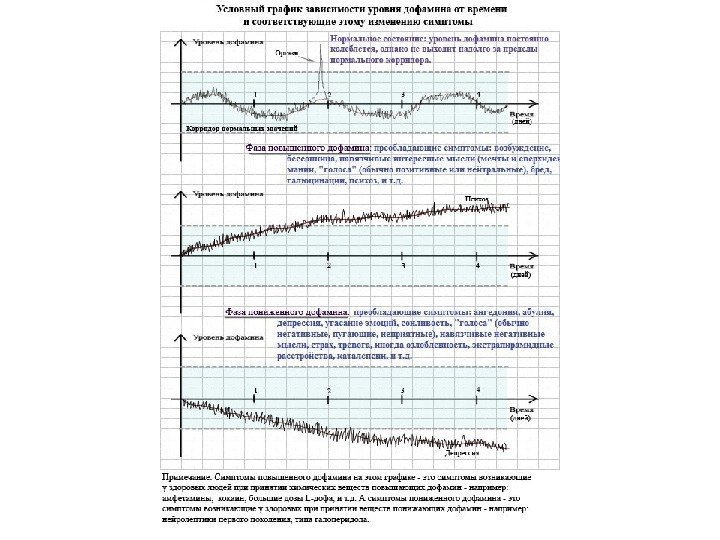
В 1954 г. Дж. Олдс и П. Милнер обнаружили центры « системы награды или подкрепления » в головном мозге. Дофамин является нейромедиатором, с помощью которого реализуется эмоционально-положительное состояние, его называют « молекулой удовольствия » или « антистрессовой молекулой » .
Изменение потока дофамина в мозгу мыши, пристрастившейся к кокаину
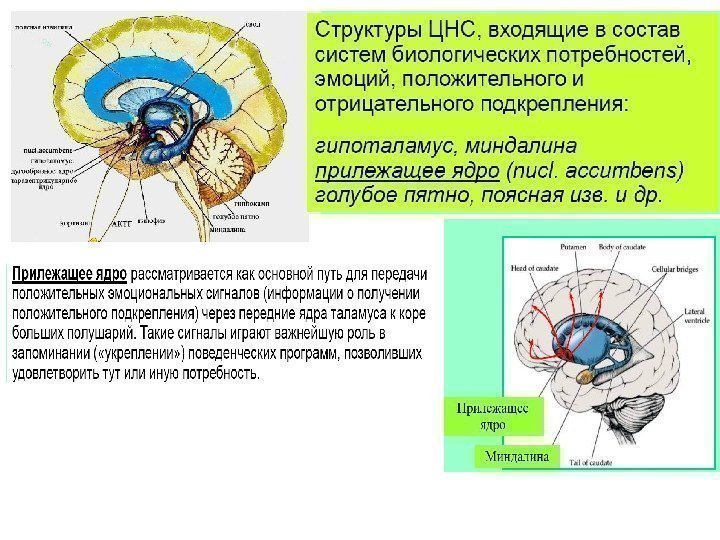
Важнейшие нейронные пути «системы вознаграждения» — мезолимбический ( голубые стрелки ) и мезокортикальный ( синие стрелки ) — образованы отростками дофаминовых нейронов вентральной области покрышки ( VTA ). По первому из них дофаминовые сигналы поступают из VTA в прилежащее ядро, миндалину, гиппокамп и префронтальную кору, по второму — только в префронтальную кору, включая ее орбитофронтальную область. Светло-оранжевым цветом показан средний мозг.
Вентральная область покрышки ( ventral tegmental area , VTA) — ключевой компонент «системы вознаграждения» (также известной как «система внутреннего подкрепления» ) в мозге млекопитающих. Примерно 55– 65% нейронов VTA — это дофаминовые (дофаминэргические) нейроны, передающие сигналы другим нейронам при помощи нейромедиатора дофамина. Дофамин играет в мозге роль «вещества удовольствия» . Дофаминовые сигналы из VTA поступают в префронтальную кору, где происходят сознательные психические процессы, в гиппокамп, управляющий запоминанием (это может быть связано с обучением на положительном опыте), в прилежащее ядро ( nucleus accumbens ), которое можно назвать «главным центром удовольствия» и которое отвечает за мотивацию, привязанности и зависимости, а также в другие «эмоциональные» отделы мозга, такие как миндалина ( amygdala )
Система награды мозга (вентральная область покрышки, прилежащее ядро, голубое пятно), центральный медиатор — дофамин
Образование дополнительных дендритных шипиков повышает чувствительность к наркотикам
Избыток и недостаток сигнала Шизофрения — группа тяжелых, обычно хронических нарушений психики, проявляющихся в расстройствах восприятия, мышления, эмоций и поведения.
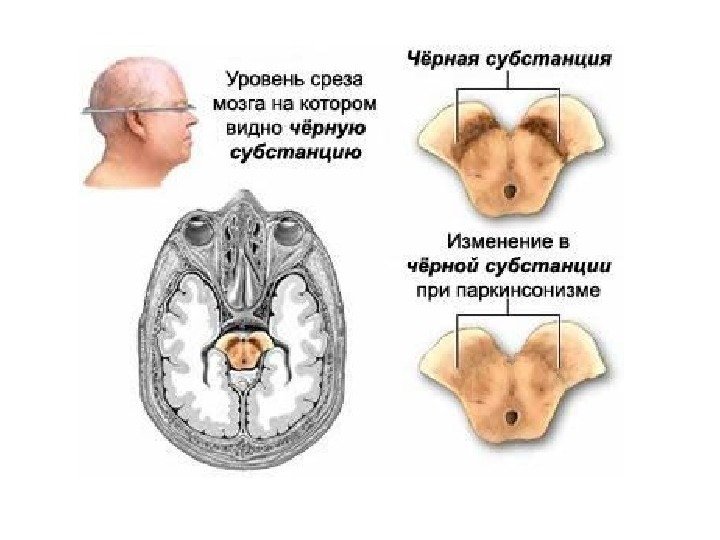
НЕДОСТАТОК СИГНАЛА Болезнь Паркинсона – впервые описана Джеймсом Паркинсоном в 1817 году в «Эссе о дрожжательном параличе» . Основу клинической картины БП составляет классическая триада: акинезия (гипокинезия), мышечная ригидность и тремор покоя.
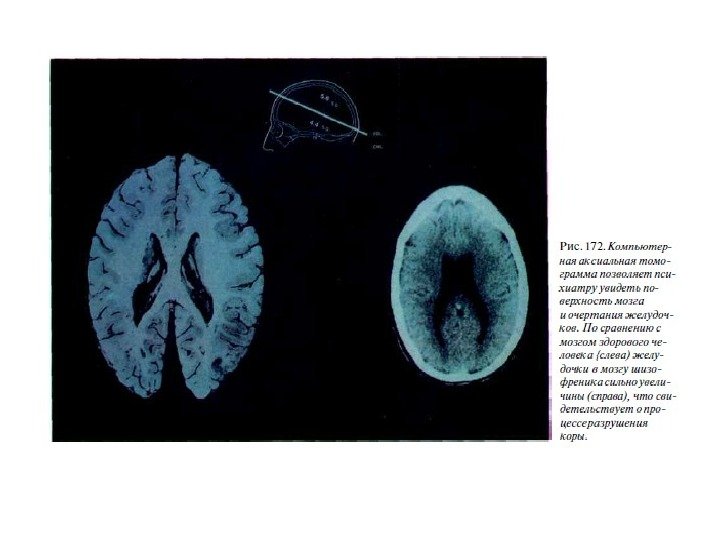
Болезнь Паркинсона (БП) — хроническое прогрессирующее заболевание головного мозга с дегенерацией нейронов черной субстанции и нарушением функции базальных ганглиев. БП — одна из самых частых форм первичных хронических нейродегенеративных заболеваний. Распространенность БП достаточно высока и колеблется от 67 до 350 случаев на 100 тыс. населения. Самая высокая распространенность зарегистрирована в США, самая низкая из европейских стран — в Швеции. Заболевание может быть спорадическим и наследственным. В настоящее время идентифицировано более 15 генов наследственных форм первичного паркинсонизма.
Продукция АКМ в нейронах s. nigra при болезни Паркинсона
Причины гибели нейронов при болезни Паркинсона
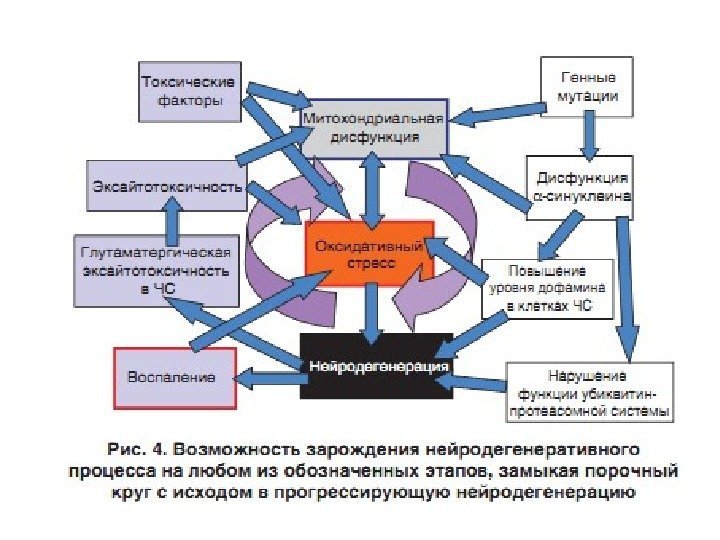
Расстройства внутриклеточного метаболизма, способствующие возникновению нейродегенерации (Крыжановский и др. , 2000): — нарушение ЭТЦ (I ФК ЭТЦ) и повреждение митохондрий ( МРТР — N-метил-4 -фенил-1, 2, 3, 6 -тетрагидро-пиридин) ; — энергетический дефицит нейрона; — усиление свободнорадикального окисления с образованием токсичных продуктов СРО; — перегрузка Fe 2+ нейронов черной субстанции ( ферроптоз ); — Са-перегрузка нейронов; — нарушение метаболизма ксенобиотиков.
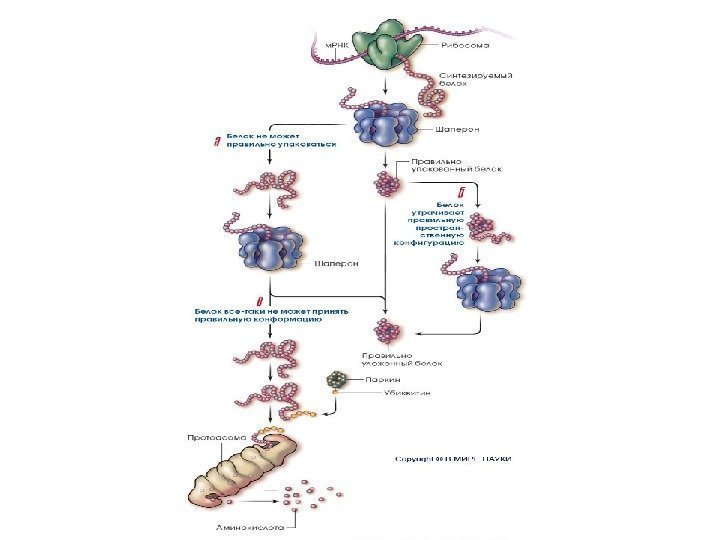
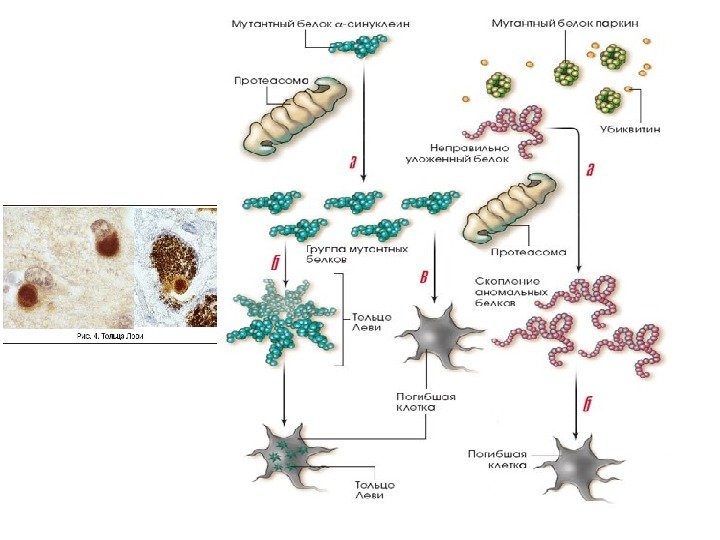
1. PARK 1 (α-синуклеин). Белок альфа-синуклеин играет роль в синаптическом везикулярном транспорте и хранении нейротрансмиттеров. Мутации в гене α-синуклеина → изменение структуры белка → его накопление в нейроне → агрегация белка с образованием телец Леви. В настоящее время α-синуклеин — ключевой молекулярный маркер БП. 2. PARK 2 (паркин). Мутации в гене паркина являются частой причиной раннего паркинсонизма. (до 50 % семейных форм и около 15 % спорадических случаев) Паркин представляет собой убиквитин-протеинлигазу типа Е 3 , функция которой заключается в доставке аномальных белков в протеасомный комплекс для последующего расщепления.
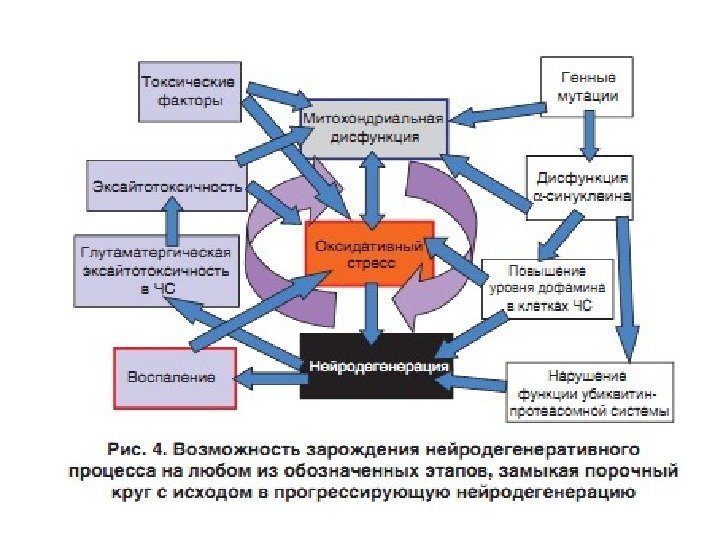
Конформационные болезни (КБ) – это заболевания, связанные с нарушнием механизмов нативной укладки клеточных белков (фолдинг) в процессе их созревания и выполнения ими физиологических функций. При всех этих заболеваниях наблюдается митохондриальная дисфункция , приводящая к повышенной продукции АФК, которые способствуют нарушению нормального фолдинга белков и накопления молекул с аномальной конформацией.
Роль ЭТЦ митохондрий в развитии нейродегенеративных заболеваний – болезни Паркинсона (PD), бокового амиотрофического склероза (ALS), болезни Гентингтона (HD), болезни Альцгеймера (AD)
Митохондриально-направленная терапия нейродегенеративных заболеваний
Митохондриально-направленные антиоксиданты – новое поколение препаратов для лечения патологий, связанных с митохондриальной дисфункцией
ИЗБЫТОК СИГНАЛА « Глутаматная эксайтотоксичность » и механизм нейромедиаторного действия оксида азота
« Глутаматная эксайтотоксичность » . Основные механизмы глутамат-кальциевого каскада
Активный транспорт. Виды активного транспорта. ФГОУ ВПО ЮФУ каф. биохимии и микробиологии. П р и в т о р и ч н о м а к т и в н о м т р а н с п о р т е п е р е н о с в е щ е с т в а о с у щ е с т в л я е т с я з а с ч ё т г р а д и е н т а к о н ц е н т р а ц и и д р у г о в е щ е с т в а , г р а д и е н т к о т о р о г о с о з д а ё т с я з а с ч ё т р а б о т ы п е р е н о с ч и к а , и с п о л ь з у ю щ е г о э н е р г и ю м а к р о э р г и ч е с к и х с в я з е й. Первичный активный транспорт Вторичный активный транспорт. А к т и в н ы й т р а н с п о р т – т р а н с м е м б р а н н ы й п е р е н о с в е щ е с т в п р о т и в г р а д и е н т а к о н ц е н т р а ц и и с з а т р а т о й э н е р г и и м а к р о э р г и ч е с к и х с о е д и н е н и й.
Типы насосов (транспортных АТФаз) Семейство АТФаз Р-типа Семейство АТФаз F- и V-типа Суперсемейство АТФаз АВС
Строение Na + /K + -ATФазы ФГОУ ВПО ЮФУ каф. биохимии и микробиологии. П р о т о м е р ф е р м е н т а с о с т о и т и з о д н о й а л ь ф а — и о д н о й б е т а — с у б ъ е д и н и ц ы. М е м б р а н н а я ч а с т ь п р е д с т а в л е н а 1 1 а л ь ф а — с п и р а л я м и , п е р е с е к а ю щ и м е м б р а н у. П е т л я. п о м е ч е н н а я к p a c н ы м , п р и н и м а е т у ч а с т и е в ф о р м и р о в а н и и ц е н т р а с в я з ы в а н и я и о н о в N а. Н а п е т л е м е ж д у 4 — й и 5 — й к о л о н н а м и л о к а л и з о в а н ы ц е н т р с в я з ы в а н и я АТФ. З е л е н ы м ц в е т о м о к р а ш е н ы а л ь ф а — с п и р а л ь н ы е у ч а с т к и б е т а — с у б ъ е д и н и ц ы , к С — к о н ц у к о т о р о й п р и с о е д и н е н ы г л и к о з и л ь н ы е р а д и к а л ы. З в е з д о ч к а м и о т м е ч е н ы в е р о я т н ы е у ч а с т к и с в я з ы в а н и я с п е ц и ф и ч е с к о г о и н г и б и т о р а N a / K — АТФ а з ы — у а б а и н а С х е м а р а с п о л о ж е н и я N a / K — АТ Ф а э ы в к л е т о ч н о й м е м б р а н е и с т р у к т у р а е е с п е ц и ф и ч е с к о г о и н г и б и т о р а у а б а и н а
Реакционный цикл Na + /K + -ATФазы ФГОУ ВПО ЮФУ каф. биохимии и микробиологии 184 Шесть основных последовательных реакций включают: 1. ) Связынание ионов натрия Е 1 — конформером, его взаимодействие с АТР и образование фосфорилированного интермедиата. 2. ) Окклюзия ионов натрия конформацией Е 1 Р. 3. ) Активируемый ионом и магния переход Е 1 Р – Е 2 Р, приводящий к высвобождению ионов Na во внешнюю среду и связывание с ионным центром K. 4. ) Окклюдирование ионов K. 5. ) Дефосфорилирование фермента, приводящее к высвобождению ионов калия во внутриклеточное пространство. 6. ) Переход конформации E 2 в
Связывание ионов Na + и K в ионных центрах Na + /K + -ATФазы ФГОУ ВПО ЮФУ каф. биохимии и микробиологииа — к р и с т а л л и ч е с к а я р е ш е т к а с о з д а в а е м а я 1 2 к и с л о р о д н ы м и а т о м а м и д и к а р б о н о в ы к а м и н о к и с л о т в к о н ф о р м а ц и и с о о т в е т с т в у ю щ е й с в я з ы в а н и ю т р е х и о н о в н а т р и я ( Е 1 и л и д в у х и о н о в к а л и я Е 2 ) b — п е т л я м е ж д у 2 — й и 3 — й п е п т и д н о й ц е п и а л ь ф а — с у б ъ е д и н и ц ы , у ч а с т в у ю щ а я в ф о р м и р о в а н и и и о н н о г о ц е н т р а ( к р а с н ы м и т о ч к а м и у к а з а н а л о к а л и з а ц и я д и к а р б о н о в ы к а м и н о к и с л о т ). В д в и г а н и е п е т л и м е ж д у к о л о н н а м и 2 и 3 п р и к о н ф о р м а ц и о н н о м п е р е х о д е ф е р м е н т а о б е с п е ч и в а е т и з м е н и е д о с т у п н о с т и и о н н о г о ц е н т р а с н а р у ж н о й и л и в н у т р е н н е й с т о р о н ы м е м б р а н ы.
ФГОУ ВПО ЮФУ каф. биохимии и микробиологии. Все морские птицы (например, альбатрос) имеют ряд специальных приспособлений к круглогодичной жизни на морской воде — мощные солевые железы, через ноздри выводящие излишки соли из организма. При активном функционировании желез с клюва птиц регулярно стекают капли солевого секрета.
НЕДОСТАТОК СИГНАЛА Снижение выработки гонадолиберина гипоталамусом
Дефицит гонадолиберина в гипоталамусе приводит к нарушению функции семенников и дефициту тестостерона Митч Хеллер до лечения после травмы головы
Сахарный диабет 1 типа- недостаток сигнала (инсулин ) „ Диабет не болезнь, а образ жизни. Болеть диабетом — все равно, что вести машину по оживленной трассе — надо знать правила движения».
Биосинтез инсулина в β-клетках поджелудочной железы
Фредерик Г. Бантинг Джон Д. Р. Маклеод Ф. Бантинг и Ч. Бест с одной из первых подопытных собак.
МИМИКРМЯ СИГНАЛА Болезнь Грейвса (Базедова болезнь, диффузный токсический зоб) — системное аутоиммунное заболевание, развивающееся вследствие выработки антител к рецептору тиреоторопного гормона (ТТГ) , клинически проявляющееся поражением ЩЖ с развитием синдрома тиреотоксикоза. Впервые заболевание было описано в 1835 г. Робертом Грейвсом, а в 1840 г. – Карлом фон Базедовым.
Джин Рэт- лиловая леди , создавшая свой стиль, — считает себя избранницей ≪ ≫ лилового цвета. Она различает 130 его оттенков, в которые окрашено почти все, что ее окружает.
1. Существование сложных многоклеточных организмов возможно благодаря координации биохимических процессов , протекающих в их клетках. Основой такой координации служат межклеточная коммуникация и передача сигнала внутри отдельных клеток. Это даёт возможность одной клетке контролировать поведение остальных. 2. Передача сигнала (сигнальная трансдукция, signal transduction ) — это процесс, при помощи которого клетка превращает один тип сигнала или стимула в другой, что приводит к включению адаптивных программ. 3. Передача сигнала внутри клетки – это цепь последовательных биохимических реакций, осуществляемых ферментами , часть из которых активируется вторичными посредниками. Такие процессы обычно являются быстрыми : их продолжительность — порядка миллисекунд в случае ионных каналов, минут — в случае активации протеинкиназ и липид-опосредованных киназ, часы и сутки в случае экспрессии генов. 4. Сигнальные пути организованы как сигнальные каскады : количество молекул белков и других веществ, принимающих участие в передаче сигнала, возрастает на каждом последующем этапе по мере удаления от первоначального стимула. Поэтому даже относительно слабый стимул может вызывать значительный ответ. Это явление называется амплификацией сигнала.
Передача сигнала от мембраны внутрь клетки
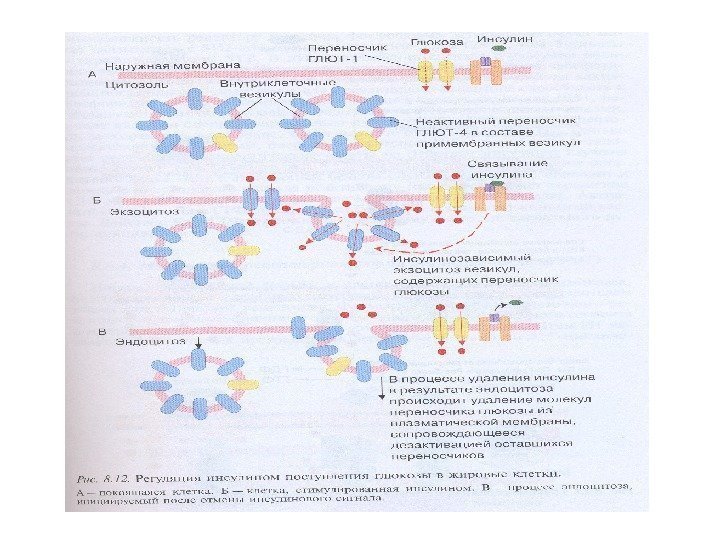
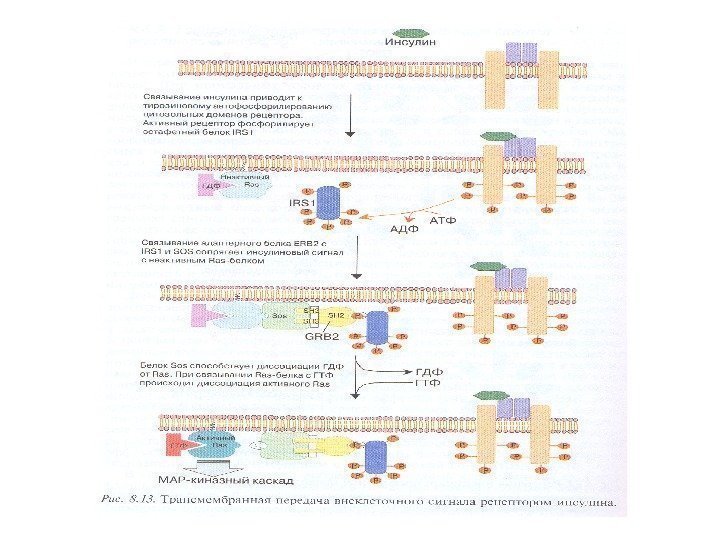
Сигнализация инсулин-подобного фактора роста (IGF 1)
проф. Дубынин В. А. , МГУ