

м'язова тканина 2013.ppt
- Количество слайдов: 40
 М’ЯЗОВА ТКАНИНА
М’ЯЗОВА ТКАНИНА

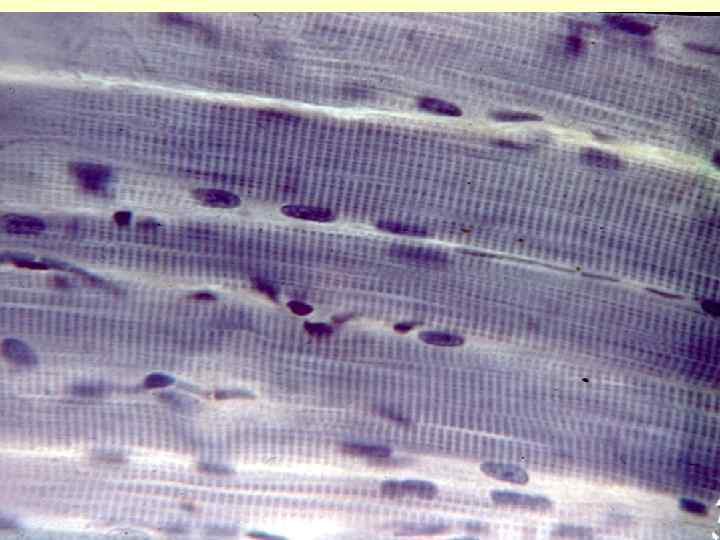

 М’язова тканина Міотом – це джерело клітин, які здатні до міграції в місця закладки конкретних м’язів. Це міогенний тип клітин. В них відбуваються зміни, а саме: клітини міотома – міграція – міобласти мітотичні – міобласти постмітотичні (злиття) – м’язеві трубочки (симпласти) - синтез білків, формування саркомерів - м’язові волокна (функція скорочення). Маркером клітин міогенного типу є десмін.
М’язова тканина Міотом – це джерело клітин, які здатні до міграції в місця закладки конкретних м’язів. Це міогенний тип клітин. В них відбуваються зміни, а саме: клітини міотома – міграція – міобласти мітотичні – міобласти постмітотичні (злиття) – м’язеві трубочки (симпласти) - синтез білків, формування саркомерів - м’язові волокна (функція скорочення). Маркером клітин міогенного типу є десмін.
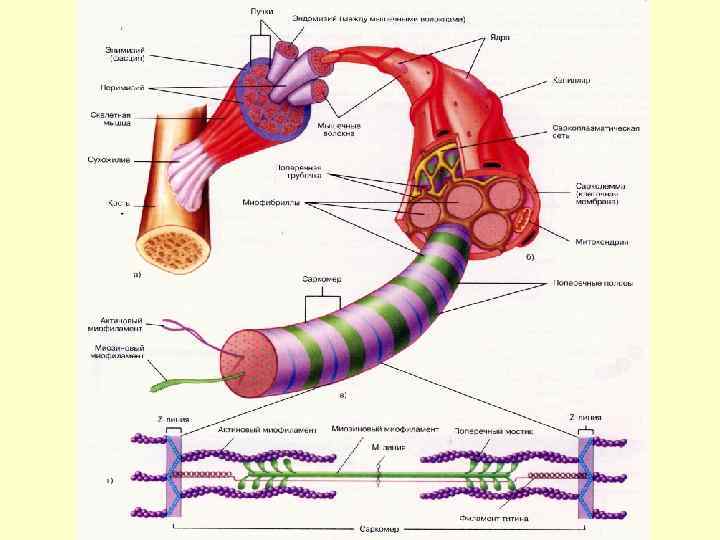
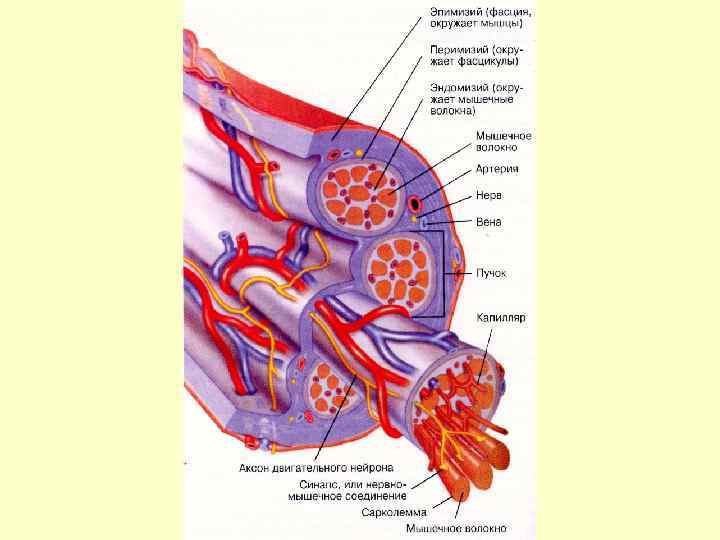
 М’язове волокно це симпласт. Оболонка волокна - плазмалема + базальна мембрана. Між ними ядра міосателітів. В саркоплазмі – міофібрили, саркоплазматична сітка – депо Са++ , гранули глікогена. Характерні структури – Ттрубочки- впячування сарколеми.
М’язове волокно це симпласт. Оболонка волокна - плазмалема + базальна мембрана. Між ними ядра міосателітів. В саркоплазмі – міофібрили, саркоплазматична сітка – депо Са++ , гранули глікогена. Характерні структури – Ттрубочки- впячування сарколеми.
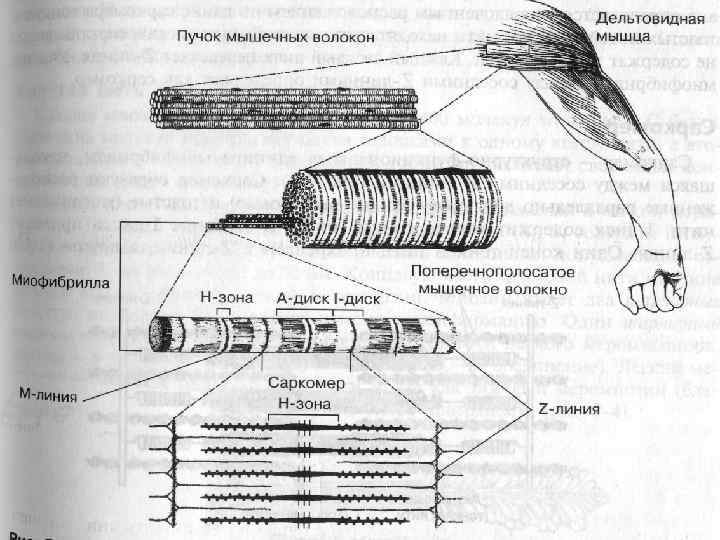
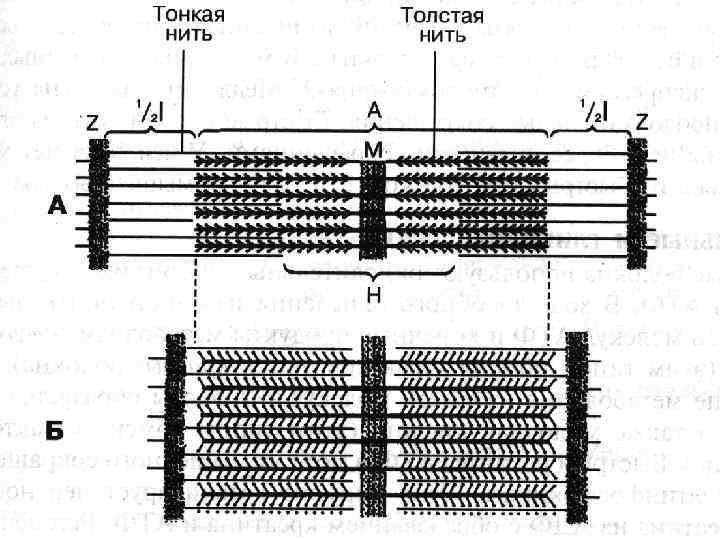
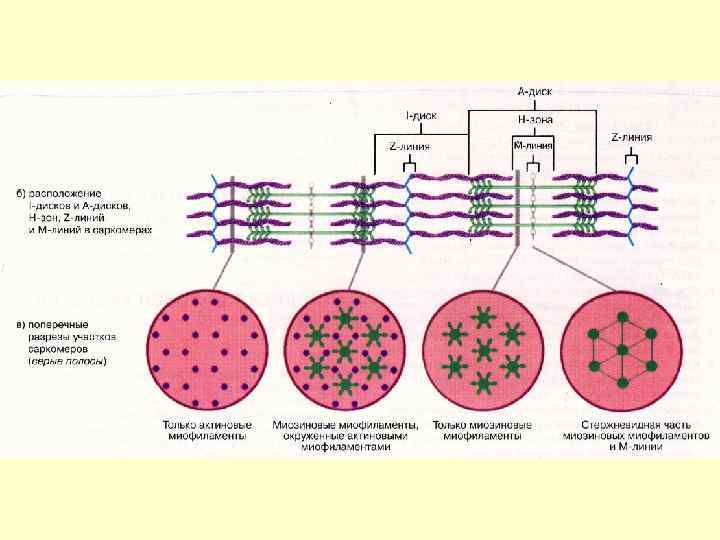
 Міофібрили Мають посмугованість, яка визначається дисками, по різному заломлюючих світло – анізотропні і ізотропні. Різне заломлення дисків визначається впорядкованим розміщенням по довжині саркомерів тонких і товстих міофіламентів – Товсті – в темних А-дисках Тонкі - в світлих І-дисках; Кожний світлий диск пересікається Z-лініями. Між сусідніми Z-лініями – саркомер Саркомер=1/2 І +А+1/2 І Н-зона – містить тільки товсті міофіламенти
Міофібрили Мають посмугованість, яка визначається дисками, по різному заломлюючих світло – анізотропні і ізотропні. Різне заломлення дисків визначається впорядкованим розміщенням по довжині саркомерів тонких і товстих міофіламентів – Товсті – в темних А-дисках Тонкі - в світлих І-дисках; Кожний світлий диск пересікається Z-лініями. Між сусідніми Z-лініями – саркомер Саркомер=1/2 І +А+1/2 І Н-зона – містить тільки товсті міофіламенти

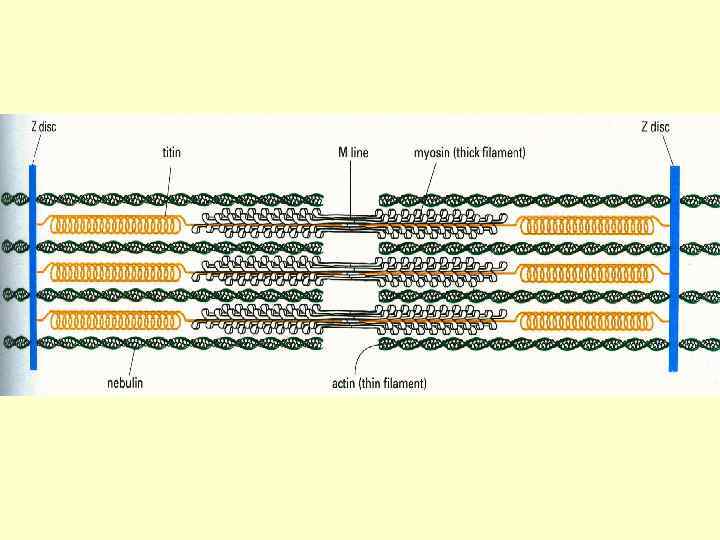
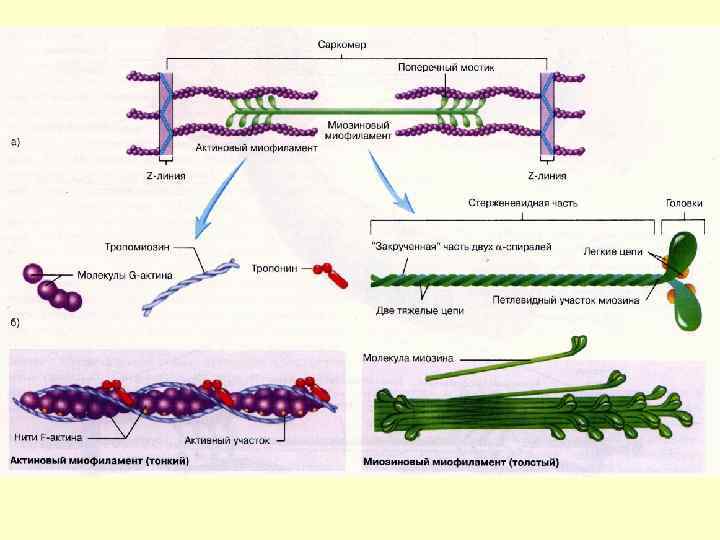
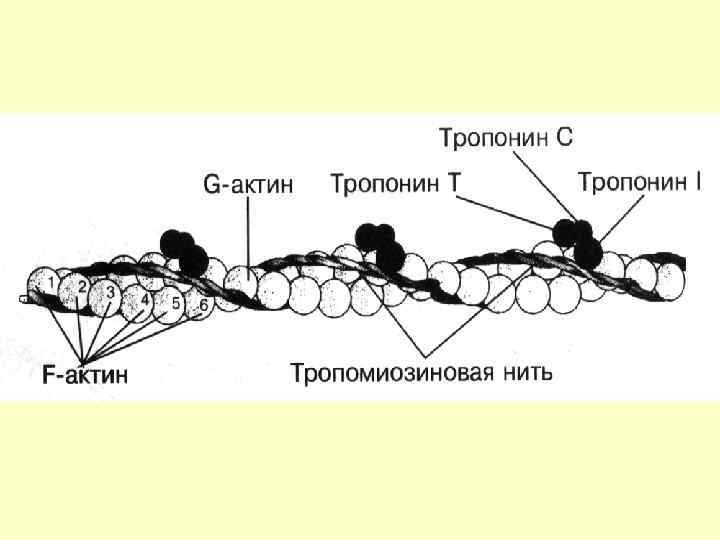
 Тонкі міофіламенти Актин , тропоміозин і тропоніни Тропонін – комплекс, утворений трьома глобулярними субодиницями – Тп. Т, Тп. С, ТпІ. Тп. Т – має ділянки для зв’язку з тропоміозином. Тп. С – Са++зв’язуючий білок ТпІ – перешкоджає взаємодії актину з міозином
Тонкі міофіламенти Актин , тропоміозин і тропоніни Тропонін – комплекс, утворений трьома глобулярними субодиницями – Тп. Т, Тп. С, ТпІ. Тп. Т – має ділянки для зв’язку з тропоміозином. Тп. С – Са++зв’язуючий білок ТпІ – перешкоджає взаємодії актину з міозином
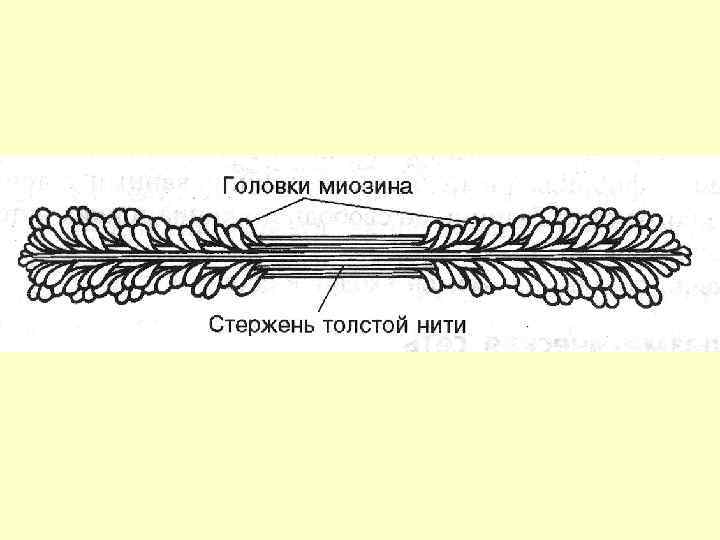
 Товсті міофіламенти Кожна нитка міозина складається із 300400 молекул міозина і С-білка, який стабілізує структуру міозинових ниток – впливає на агрегацію молекул міозина, забезпечує однаковий діаметр і довжину товстих ниток. Білок тітін – зв’язує кінці товстих ниток з Z-лінією, запобігає перерозтягненню. Небулін - зв’язує тонкі і товсті нитки.
Товсті міофіламенти Кожна нитка міозина складається із 300400 молекул міозина і С-білка, який стабілізує структуру міозинових ниток – впливає на агрегацію молекул міозина, забезпечує однаковий діаметр і довжину товстих ниток. Білок тітін – зв’язує кінці товстих ниток з Z-лінією, запобігає перерозтягненню. Небулін - зв’язує тонкі і товсті нитки.
 Z-лінії До складу Z-лінії входять альфа-актинін, десмін і віментин. У Z-лінію вплетений один кінець тонкої нитки. Другий кінець - між товстими нитками і асоційований з ними за допомогою небуліна.
Z-лінії До складу Z-лінії входять альфа-актинін, десмін і віментин. У Z-лінію вплетений один кінець тонкої нитки. Другий кінець - між товстими нитками і асоційований з ними за допомогою небуліна.
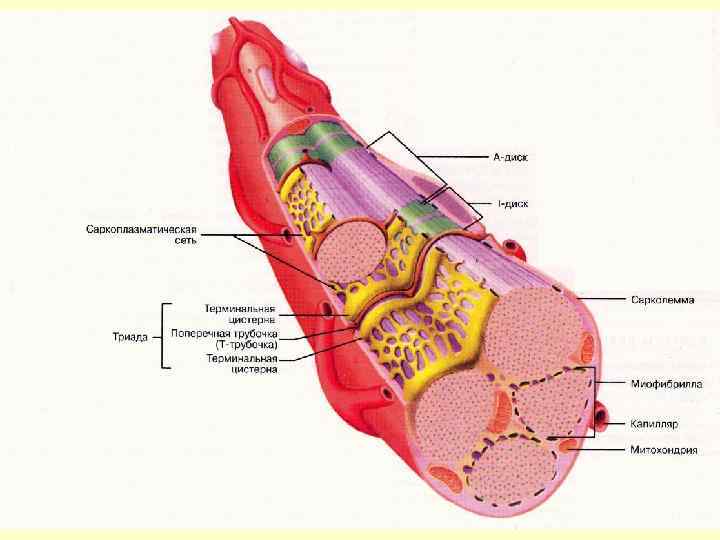
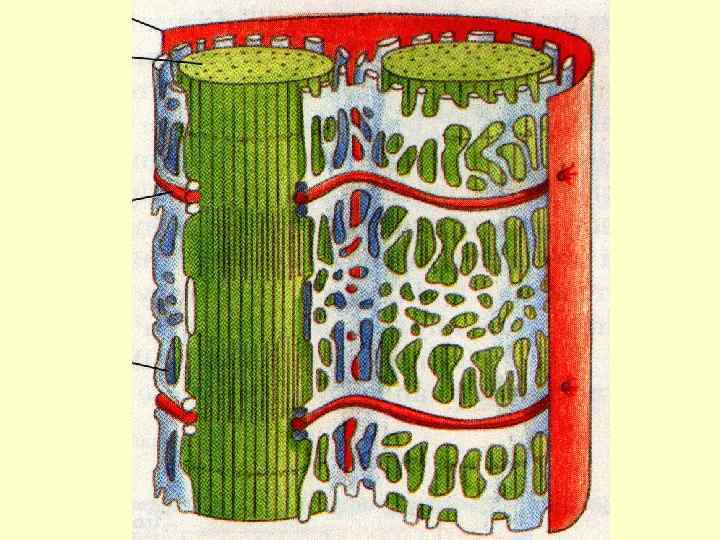
 Т-трубочки - вп’ячування плазмолеми,") Передача збудження Для передачі збудження існують спеціальні мембранні структури: а) Т-трубочки - вп’ячування плазмолеми, які ідуть в поперечному напрямку навколо міофібрили; б) L-канальці – компоненти глад. ЕПС, оточують кожну міофібрилу і орієнтовані вздовж її довгої осі. в) в області Т-трубочки L-канальці розширюються і переходять в кінцеві (термінальні) цистерни.
Передача збудження Для передачі збудження існують спеціальні мембранні структури: а) Т-трубочки - вп’ячування плазмолеми, які ідуть в поперечному напрямку навколо міофібрили; б) L-канальці – компоненти глад. ЕПС, оточують кожну міофібрилу і орієнтовані вздовж її довгої осі. в) в області Т-трубочки L-канальці розширюються і переходять в кінцеві (термінальні) цистерни.
") Транспортні системи в мембрані цистерн В мембрані термінальних цистерн є 2 транспортні системи: 1) Са++ насос, який використовує енергію АТФ, активно закачує всередину цистерн іони Са ++, тому в стані спокою поза цистернами, в саркоплазмі дуже низька концентрація , а всередині цистерн – висока концентрація цих іонів.
Транспортні системи в мембрані цистерн В мембрані термінальних цистерн є 2 транспортні системи: 1) Са++ насос, який використовує енергію АТФ, активно закачує всередину цистерн іони Са ++, тому в стані спокою поза цистернами, в саркоплазмі дуже низька концентрація , а всередині цистерн – висока концентрація цих іонів.
 Са++ канали. В м’язовому волокні в стані спокою канали закриті, а при збудженні") 2) Са++ канали. В м’язовому волокні в стані спокою канали закриті, а при збудженні - відкриваються. Кальсеквестрин – зв’язує Са++ всередині саркоплазматичного ретикулуму.
2) Са++ канали. В м’язовому волокні в стані спокою канали закриті, а при збудженні - відкриваються. Кальсеквестрин – зв’язує Са++ всередині саркоплазматичного ретикулуму.
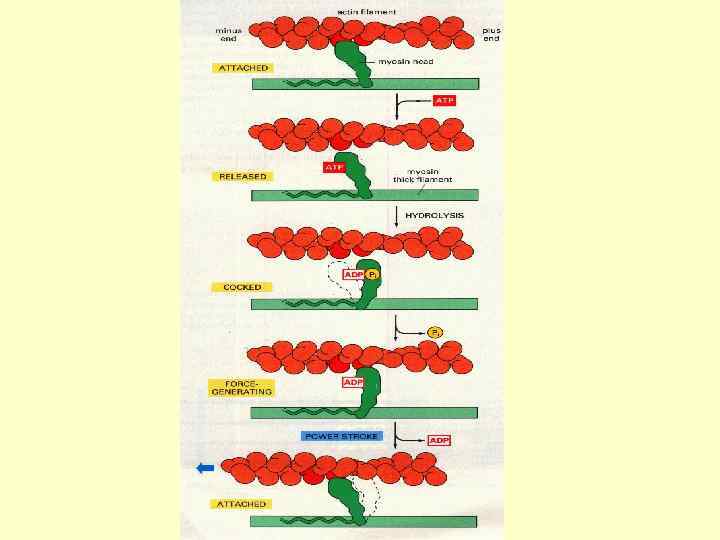
 М’язове скорочення починається зі зв’язування Са з субодиницею Тп. С тропоніну, в результаті чого оголюється ділянка зв’язування міозину на молекулі актину. На другому етапі міозинова головка зв’язується з актином, і АТФ розщеплюється до АДФ, даючи енергію для руху міозинової головки. Внаслідок цієї зміни міозину, зв’язані з ним тонкі філаменти ковзають по товстим філаментам. Цей процес повторюється багато разів протягом одного скорочення, призводить до повного перекривання актину і міозину і обумовлює вкорочення всього м’язевого волокна.
М’язове скорочення починається зі зв’язування Са з субодиницею Тп. С тропоніну, в результаті чого оголюється ділянка зв’язування міозину на молекулі актину. На другому етапі міозинова головка зв’язується з актином, і АТФ розщеплюється до АДФ, даючи енергію для руху міозинової головки. Внаслідок цієї зміни міозину, зв’язані з ним тонкі філаменти ковзають по товстим філаментам. Цей процес повторюється багато разів протягом одного скорочення, призводить до повного перекривання актину і міозину і обумовлює вкорочення всього м’язевого волокна.
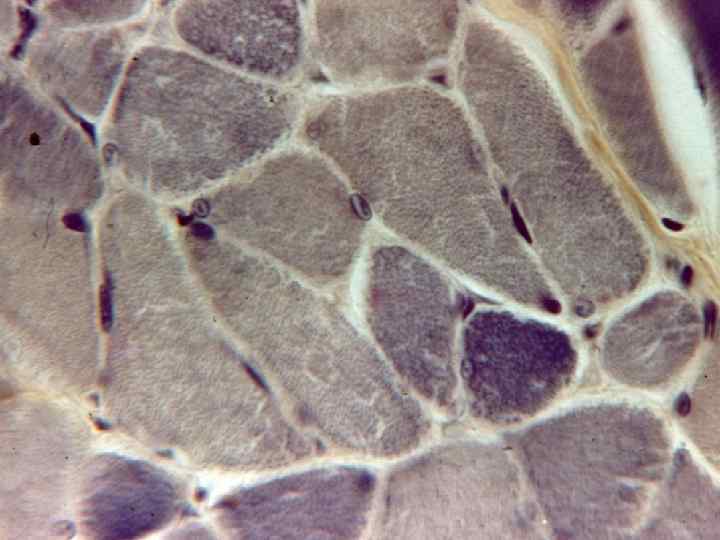
 М’язові волокна за швидкістю скорочення БІЛІ м’язові волокна виконують інтенсивну, але короткочасну роботу. Осн. джерело енергії – анаеробний (не потребує кисню) розпад глікогену або глюкози до молочної кислоти. Якщо кисень не використовується, то не потрібен і міоглобін, тому і колір такий. Вміст глікогену високий. Всі реакції анаеробного розпаду відбуваються в цитозолі, мітохондрії не приймають участь, низька активність мітохондріальних ферментів. Швидкість розпаду АТФ (АТФ-азна активність) значно вища, ніж в червоних.
М’язові волокна за швидкістю скорочення БІЛІ м’язові волокна виконують інтенсивну, але короткочасну роботу. Осн. джерело енергії – анаеробний (не потребує кисню) розпад глікогену або глюкози до молочної кислоти. Якщо кисень не використовується, то не потрібен і міоглобін, тому і колір такий. Вміст глікогену високий. Всі реакції анаеробного розпаду відбуваються в цитозолі, мітохондрії не приймають участь, низька активність мітохондріальних ферментів. Швидкість розпаду АТФ (АТФ-азна активність) значно вища, ніж в червоних.
 М’язеві волокна за швидкістю скорочення ЧЕРВОНІ м’язові волокна – повільного типу, тривала робота. Енергія за рахунок аеробного (окислювального) розпаду енергетичних субстратів – глюкози, жирних кислот. Для створення запасу кисню – міоглобін, тому колір червоний. Оскільки утворюється велика кількість АТФ, немає необхідності створювати запаси глікогену. Численні ліпідні включення і висока активність СДГ (сукцинатдегідрогенази) – в мітохондріях (один із ферментів циклу Кребса).
М’язеві волокна за швидкістю скорочення ЧЕРВОНІ м’язові волокна – повільного типу, тривала робота. Енергія за рахунок аеробного (окислювального) розпаду енергетичних субстратів – глюкози, жирних кислот. Для створення запасу кисню – міоглобін, тому колір червоний. Оскільки утворюється велика кількість АТФ, немає необхідності створювати запаси глікогену. Численні ліпідні включення і висока активність СДГ (сукцинатдегідрогенази) – в мітохондріях (один із ферментів циклу Кребса).
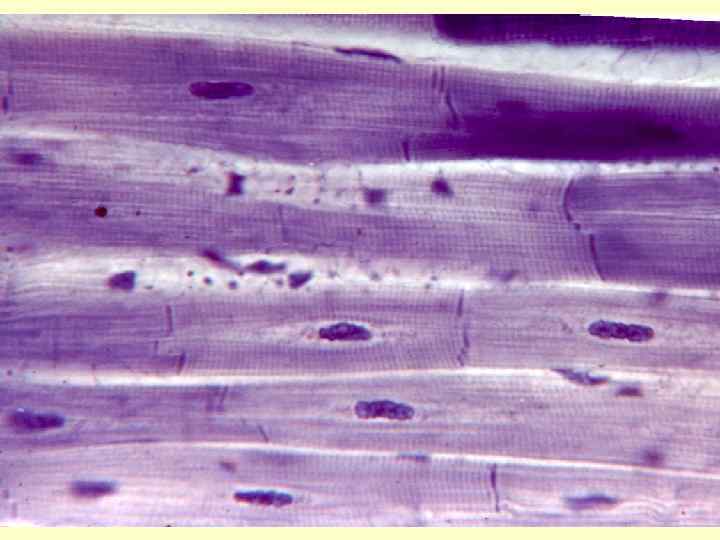
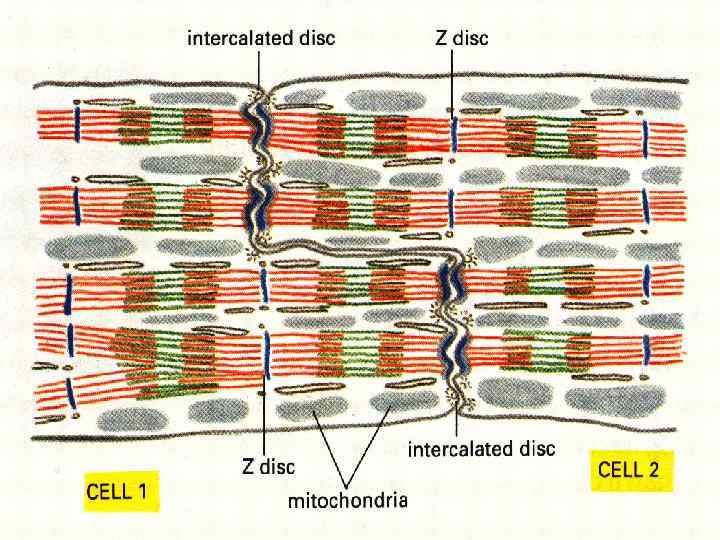
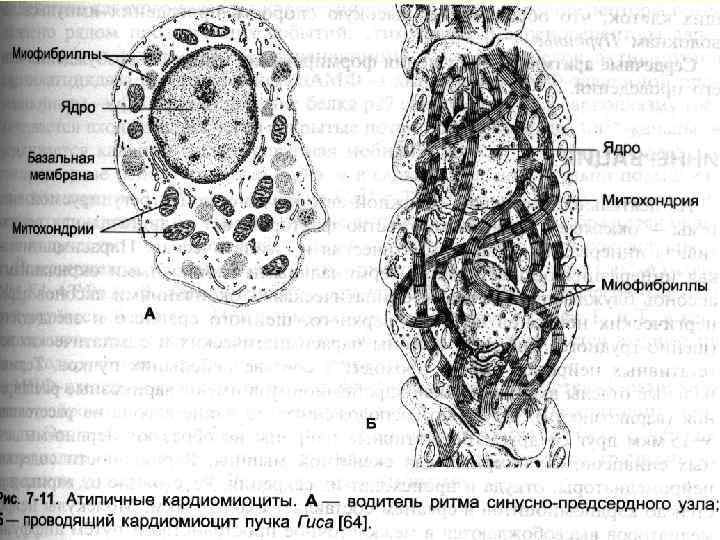
 Кардіоміоцити • Робочі – міофібрили, саркоплазматичний ретикулум, міжклітинні контакти. Мітохондрії. Т -трубочки на рівні Z-ліній. Вставні диски. • Атипові - формують провідну систему серця. Серед них водії ритму і провідні міоцити. Головний водій ритму – клітини синуснопередсердного вузла – генерують ритм 60 -90 імпульсів за хвилину. Провідні – утворюють довгі волокна. Волокна Пуркіньє – найбільші клітини міокарду. • Секреторні – атріопентин – гормон, регулюючий артеріальний тиск.
Кардіоміоцити • Робочі – міофібрили, саркоплазматичний ретикулум, міжклітинні контакти. Мітохондрії. Т -трубочки на рівні Z-ліній. Вставні диски. • Атипові - формують провідну систему серця. Серед них водії ритму і провідні міоцити. Головний водій ритму – клітини синуснопередсердного вузла – генерують ритм 60 -90 імпульсів за хвилину. Провідні – утворюють довгі волокна. Волокна Пуркіньє – найбільші клітини міокарду. • Секреторні – атріопентин – гормон, регулюючий артеріальний тиск.
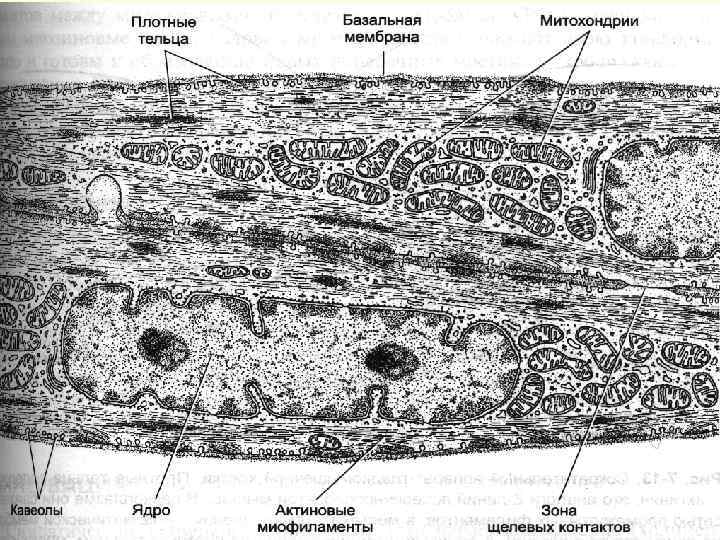
 Гладенька м’язова тканина Молекули актину утв. стабільні актинові нитки, прикріплені до щільних тілець. Міозинові формуються між актиновими міофіламентами при скороченні ГМК. Зборку товстих ниток і взаємодію актинових і міозинових ниток активують іони кальцію. Щільні тільця – аналог Z-ліній - специфічні компоненти цитоскелета, містять альфа-актинін. Вони діляться на 2 види: 1) щільні пластинки плазмолеми – пучки тонких мікрофіламентів, які йдуть під плазмалемою і формують ребристий каркас міоцита і лише між пластинками плазмолема здатна утворювати кавеоли. 2) щільні тільця цитоплазми – мають овальну форму, з’єднані актином в ланцюги (місця прикріплення тонких міофіламентів)
Гладенька м’язова тканина Молекули актину утв. стабільні актинові нитки, прикріплені до щільних тілець. Міозинові формуються між актиновими міофіламентами при скороченні ГМК. Зборку товстих ниток і взаємодію актинових і міозинових ниток активують іони кальцію. Щільні тільця – аналог Z-ліній - специфічні компоненти цитоскелета, містять альфа-актинін. Вони діляться на 2 види: 1) щільні пластинки плазмолеми – пучки тонких мікрофіламентів, які йдуть під плазмалемою і формують ребристий каркас міоцита і лише між пластинками плазмолема здатна утворювати кавеоли. 2) щільні тільця цитоплазми – мають овальну форму, з’єднані актином в ланцюги (місця прикріплення тонких міофіламентів)
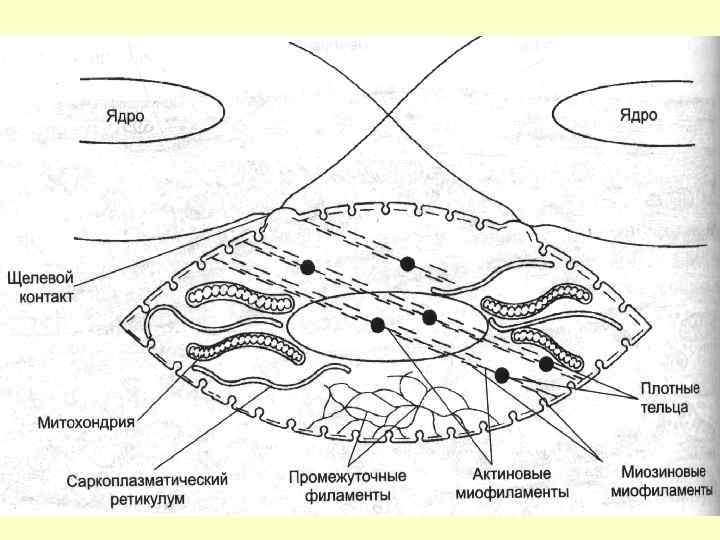
 Депо Са++ - саркоплазматичний ретикулум + кавеоли Са++ АТФаза постійно відкачує Са++ із цитоплазми ГМК у цистерни ЕПС. Активація Са++каналів відбувається при зміні мембранного потенціалу й за допомогою рецепторів ріанодина та інозитолтрифосфата.
Депо Са++ - саркоплазматичний ретикулум + кавеоли Са++ АТФаза постійно відкачує Са++ із цитоплазми ГМК у цистерни ЕПС. Активація Са++каналів відбувається при зміні мембранного потенціалу й за допомогою рецепторів ріанодина та інозитолтрифосфата.

 ДЯКУЮ ЗА УВАГУ
ДЯКУЮ ЗА УВАГУ